
Abstract
Obesity-induced adipose inflammation plays a crucial role in the development of obesity-induced metabolic disorders such as insulin resistance and type 2 diabetes. In the presence of obesity, hypertrophic adipocytes release inflammatory mediators, including tumor necrosis factor-alpha (TNFα) and monocyte chemoattractant protein-1 (MCP-1), which enhance the recruitment and activation of macrophages, and in turn augment adipose inflammation. We demonstrate that the soy peptide Phe–Leu–Val (FLV) reduces inflammatory responses and insulin resistance in mature adipocytes. Specifically, the soy peptide FLV inhibits the release of inflammatory cytokines (TNFα, MCP-1, and IL-6) from both TNFα-stimulated adipocytes and cocultured adipocytes/macrophages. This inhibition is mediated by the inactivation of the inflammatory signaling molecules c-Jun N-terminal kinase (JNK) and IκB kinase (IKK), and the downregulation of IκBα in the adipocytes. In addition, soy peptide FLV enhances insulin responsiveness and increases glucose uptake in adipocytes. More importantly, we, for the first time, found that adipocytes express peptide transporter 2 (PepT2) protein, and the beneficial action of the soy peptide FLV was disrupted by the peptide transporter inhibitor GlySar. These findings suggest that soy peptide FLV is transported into adipocytes by PepT2 and then downregulates TNFα-induced inflammatory signaling, thereby increasing insulin responsiveness in the cells. The soy peptide FLV, therefore, has the potential to prevent obesity-induced adipose inflammation and insulin resistance.
Introduction
O
Food-derived bioactive peptides have received increasing attention due to their potential efficacies for disease prevention and treatment. Soybeans contain numerous bioactive components, including isoflavones, saponins, and phytic acids, and their action against various diseases has been widely recognized. 14 In addition to the bioactive components of soybeans, soy peptides derived from the soy proteins glycinin and β-conglycinin, which account for 85% of total soy protein in soy hydrolysates, 14,15 have antihypertensive, 16 immunomodulatory, 17 and antioxidative 15 effects. Of note, recent reports show that soy peptides are beneficial for protecting against inflammatory diseases. For example, soy peptide Val–Pro–Tyr (VPY), which corresponds to amino acid residues 152–154 of the soy 11S globulin (glycinin), inhibits the production of inflammatory mediators (such as IL-8 and TNFα) in intestinal epithelial and immune cells, and also reduces dextran sodium sulfate-induced colitis in mice by downregulating the expression of inflammatory cytokines (TNFα, IL-6, IL-1β, and IFN-γ) in the colon. 18 Furthermore, the milk-derived peptide Val–Pro–Pro inhibits the expression of inflammatory cytokines, such as TNFα and IL-1β, in cocultured adipocytes/macrophages, and in the adipose tissue of diet-induced obese mice, 19 indicating that the milk peptide has the potential to protect against obesity-induced adipose inflammation.
In this study, we demonstrate that the soy peptide Phe–Leu–Val (FLV), a β-conglycinin-derived peptide comprising amino acid residues 169–171 of soy 7S globulin, 20 is transported into adipocytes by peptide transporter 2 (PepT2), suppresses TNFα-mediated inflammatory responses in both adipocytes and cocultured adipocytes/macrophages, and improves insulin responsiveness in the adipocytes. The soy peptide FLV might, therefore, be useful for protecting against obesity-induced adipose inflammation and metabolic dysregulation.
Materials and Methods
Soy peptide
Synthetic soy tripeptide FLV was synthesized by GL Biochem (Shanghai, China) and the purity of the peptide was >96% (on the basis of HPLC analysis).
Peptide transporters
Total mRNA and protein for peptide transporter-1 (PepT1; SLC15A1) and PepT2 (SLC15A2) were isolated from various tissues, such as liver, lung, and jejunum, and epididymal adipose tissue from C57BL/6 mice (male, 8 weeks old) (Orient, Ltd., Busan, Korea).
Cell culture
Murine macrophage Raw 264.7 cells were obtained from the Korean Cell Line Bank (KCLB40071, Seoul, Korea) and cultured in RPMI-1640 medium (Gibco BRL, Grand Island, NY, USA) supplemented with 10% (vol/vol) fetal bovine serum (Gibco BRL), 10 mL/L penicillin–streptomycin (Invitrogen, Carlsbad, CA, USA) and 2 mL/L gentamicin (Gibco BRL) at 37°C and 5% CO2. The 3T3-L1 preadipocytes (ATCC, Manassas, VA, USA) were cultured in a basal medium of DMEM supplemented with 200 μM ascorbic acid, 10% newborn calf serum, 10 mL/L penicillin–streptomycin, and 2 mL/L gentamicin at 37°C in a 5% CO2 atmosphere. Differentiation of the 3T3-L1 preadipocytes was induced after the cells had reached confluence by incubation with adipogenic agents (0.5 mM 3-isobutyl-1-methyl-xanthine, 0.25 μM dexamethasone, and 10 μg/mL insulin [Sigma-Aldrich, St. Louis, MO, USA]) in DMEM containing 10% fetal bovine serum, as described previously. 6,21 After 40–42 h, the cell culture medium was changed to a maturation medium (the basal medium plus 5 μg/mL insulin), and the maturation medium was changed every 2 days for 5 days.
Adipocytes and macrophages were cocultured in a contact system as previously described. 6,7 Briefly, RAW 264.7 cells (3 × 105 cells/mL) were plated in 24-well plates containing 3T3-L1 adipocytes (3 × 105 cells/mL). The cells were cultured for 16 h in contact with each other and harvested. As a control, adipocytes and macrophages were also cultured separately, with equal cell numbers per well as those in the contact system; the cells were mixed after harvest.
Polymerase chain reaction analysis
Total RNA extracted from tissues or cultured cells were reverse transcribed to generate cDNA using M-MLV reverse transcriptase (Promega, Madison, WI, USA). Real-time polymerase chain reaction (PCR) of the cDNA was performed in duplicate with the SYBR Premix Ex Taq Kit (TaKaRa Bio, Inc., Kyoto, Japan). All reactions were performed by the same procedure: initial denaturation at 95°C for 5 sec and 60°C for 30 sec. All values for genes of interest were normalized to values for β-actin. The mouse primer sequences used are shown in Table 1. PepT1 and PepT2 data were analyzed using Thermal Cycler Dice Real Time System Software (TaKaRa Bio, Inc.).
PCR, polymerase chain reaction; PepT2, peptide transporter 2.
Measurement of cytokine levels by enzyme-linked immunosorbent assays
Cytokine levels in culture supernatants were measured using enzyme-linked immunosorbent assays (ELISAs). The assays were conducted using an OptEIA™ sets for mouse TNFα and mouse MCP-1 (BD Bioscience Pharmingen, CA, USA), and sets for mouse IL-6 and mouse adiponectin (R&D Systems, Minneapolis, MN, USA). Cytokine levels were derived from standard curves using the curve-fitting program SoftMax (Molecular Devices, Sunnyvale, CA, USA).
Western blotting analysis
Tissue samples were prepared and their protein content determined. Equal quantities of protein (80 μg) were loaded and electrophoresed on an 11% SDS-polyacrylamide gel. After transferring the separated proteins to a nitrocellulose membrane, the membrane was incubated overnight in a blocking buffer [5% non-fat dry milk, 10 mM Tris (pH 7.4) 100 mM NaCl, and 0.1% Tween 20] and treated with rabbit polyclonal PepT1 and PepT2 antibodies (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) for 2 h. After washing, the membrane was incubated with a horseradish peroxidase-conjugated anti-rabbit IgG antibody. The immunoreactive proteins were detected with a chemiluminescent system (the ECL Kit, Amersham, United Kingdom). After exposure to X-ray film, band densities were calculated from optical densities with an image analyzer.
The 3T3-L1 adipocytes were plated at 3 × 105 cells/well in six-well plates and incubated with TNFα in the presence or absence of FLV (0.01–0.1 μM). Total cellular protein was harvested in phosphate-buffered saline lysis buffer (10 mM Tris–HCl, 10 mM NaCl, 0.1 mM EDTA, 50 mM NaF, 10 mM Na4P2O7, 1 mM MgCl2, 0.5% deoxycholate, 1% IGEPAL, and protease inhibitors cocktail). The cell lysates (protein quantities 20–50 μg) were electrophoresed on an 11% SDS-PAGE gel and transferred to a nitrocellulose membrane, and the procedure completed as described above. The following antibodies were used: phosphorylated c-Jun N-terminal kinase (p-JNK), JNK, phosphorylated IκB kinase (p-IKK) and IKKβ (Cell Signaling, Neverly, MA, USA), IκBα (Santa Cruz Biotechnology, Inc.), p-Akt (Akt-pSer473 [also known as protein kinase B]) and AKT (Cell Signaling), phosphorylated insulin receptor substrate-1 (p-IRS-1; Tyr 608; Millipore, Temecula, CA, USA), and β-actin (Sigma-Aldrich), which was used as a loading control.
Measurement of peptide uptake
Mature 3T3-L1 adipocytes in 24-well plates (3 × 105 cells/well) were washed twice with 37°C sodium medium (137 mM NaCl, 5.4 mM KCl, 2.8 mM CaCl2, 1.2 mM MgSO4, 10 mM HEPES, pH 7.4) and then placed in sodium medium that contained Ala–Lys–amino-4-methylcoumarin-3-acetic acid (AMCA) and either glycylsarcosine (GlySar) or soy peptide FLV for 120 min. Cells were then immediately washed three times with ice-cold sodium medium and solubilized in 500 μL 0.5% Triton X-100. The concentration of Ala–Lys–AMCA in cell lysates was determined with a SpectraMax M2 spectrofluorometer (Molecular Devices) with excitation at 350 nM and emission at 455 nM.
Statistical analyses
Results are expressed as mean ± standard error of the mean. Statistical comparisons were performed using Student's t-test or one-way analysis of variance. Differences were considered to be significant at P < .05.
Results
Soy peptide FLV inhibits inflammatory responses in adipocytes and cocultured adipocytes/macrophages
To examine the effect of FLV on obesity-induced adipose inflammation in vitro, we first examined its effect on the production of inflammatory cytokines by mature adipocytes. FLV inhibited the production of MCP-1 and IL-6 in a dose-dependent manner (Fig. 1A). Subsequently, we cocultured adipocytes/macrophages, mimicking inflamed adipose tissue in obesity, and measured the levels of MCP-1, IL-6, and TNFα, and found that FLV reduced the release of MCP-1, IL-6, and TNFα from the cocultured adipocytes/macrophages (Fig. 1B).
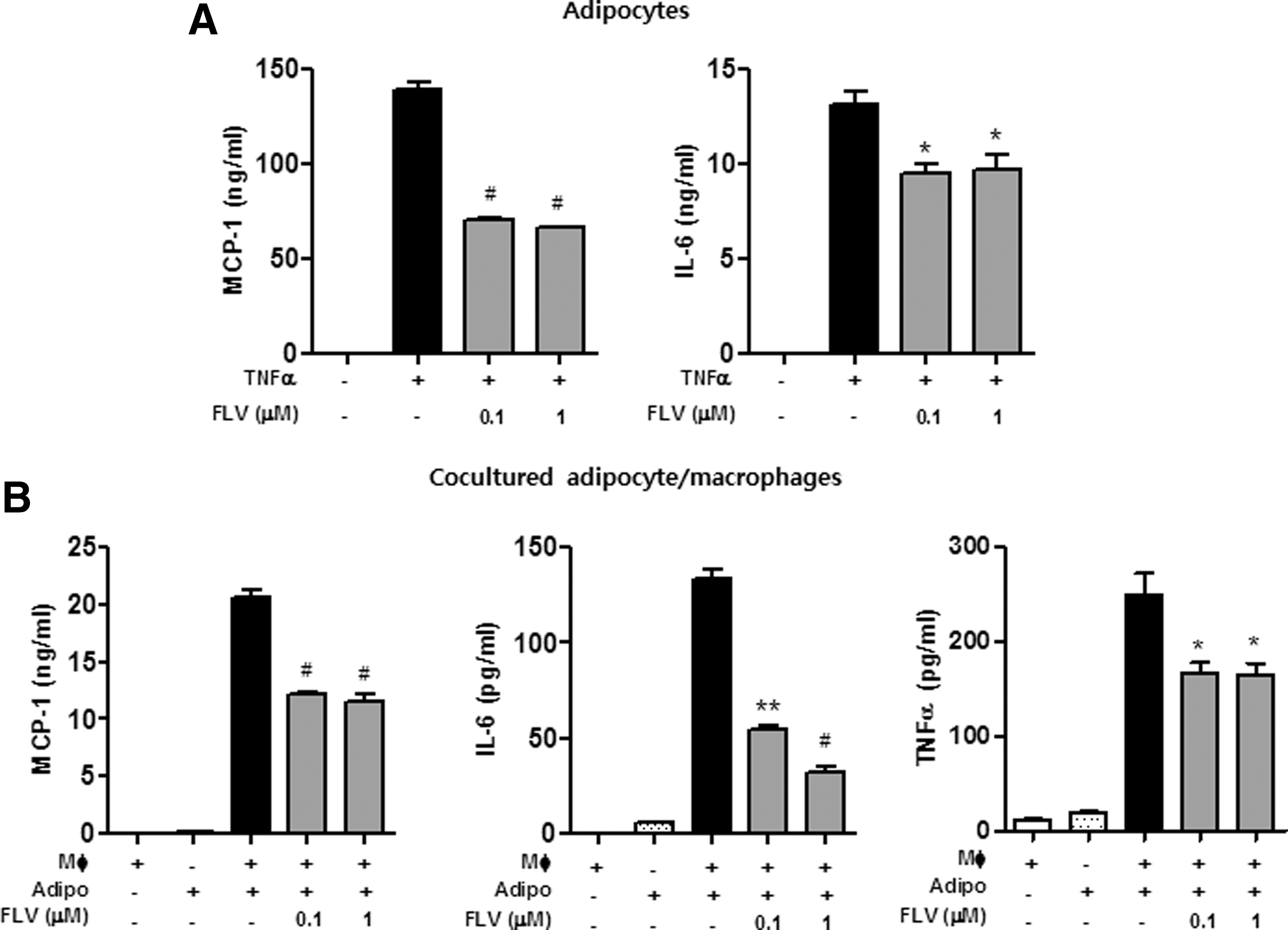
Soy peptide FLV inhibits cytokine release from adipocytes and cocultured adipocytes/macrophages.
Soy peptide FLV inhibits TNFα-induced activation of JNK, IKK, and degradation of IκBα in adipocytes
TNFα plays an important role in the activation of inflammatory signaling molecules such as JNK, IKK, and NF-κB, which are crucial for the production of inflammatory cytokines and induction of insulin resistance. 12,13,22 As shown in Figure 2, TNFα induced both phosphorylation of JNK and IKK and degradation of IκBα, and FLV inhibited the TNFα-induced inflammatory signaling pathways in adipocytes.
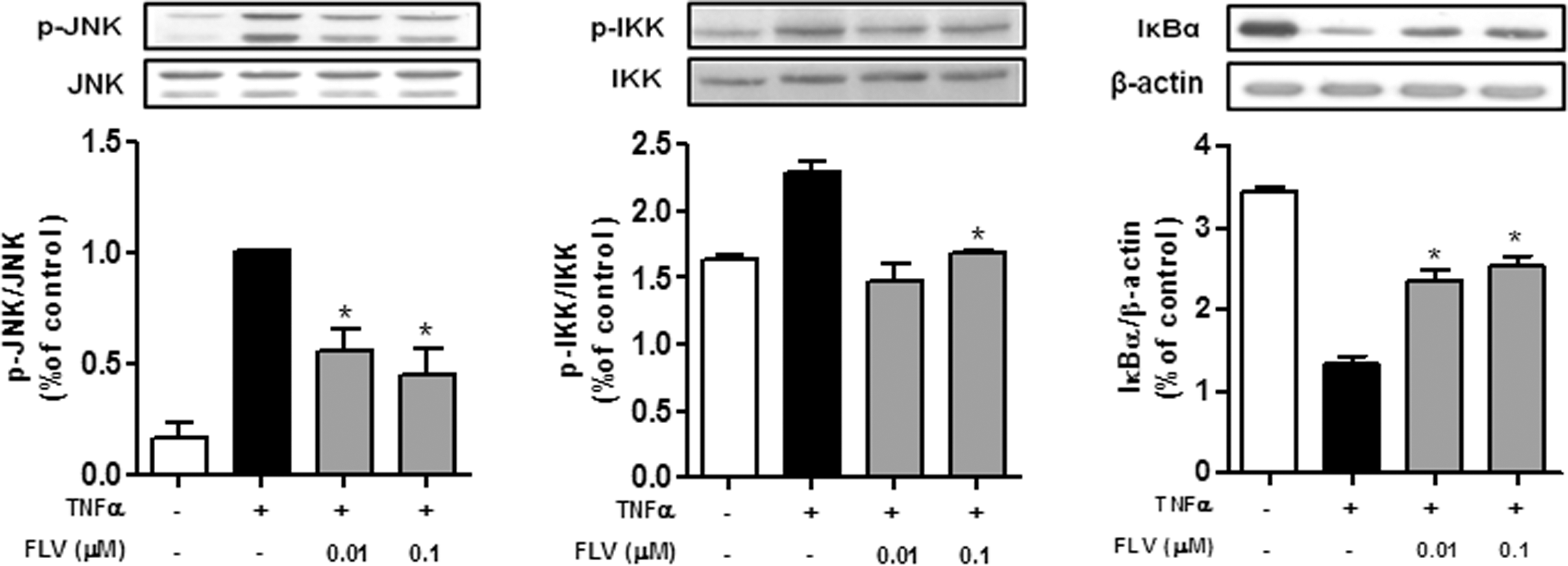
Soy peptide FLV inhibits TNFα-induced activation of JNK and IKK, and TNFα-induced degradation of IκBα in 3T3-L1 adipocytes. 3T3-L1 adipocytes (3 × 105 cells/mL) were treated with 20 ng/mL TNFα in the presence or absence of FLV (0.01–0.1 μM) for 15 min at 37°C. Cell lysates were electrophoresed on 11% SDS-PAGE gels and transferred to nitrocellulose membranes. Protein extracts were analyzed by western blotting. Values are mean ± SEM. *P < .05. IKK, IκB kinase; JNK, Jun N-terminal kinase.
Soy peptide FLV improves insulin sensitivity in adipocytes
To examine whether FLV inhibits TNFα-induced insulin resistance in 3T3-L1 adipocytes, we treated 3T3-L1 adipocytes with 10 ng/mL TNFα and 0.01–0.1 μM FLV for 24 h, and then stimulated the cells with insulin. FLV reversed the TNFα-induced decrease in activation of both IRS-1 and Akt in 3T3-L1 adipocytes (Fig. 3A) and it also increased the insulin-stimulated phosphorylation of Akt (Fig. 3B).
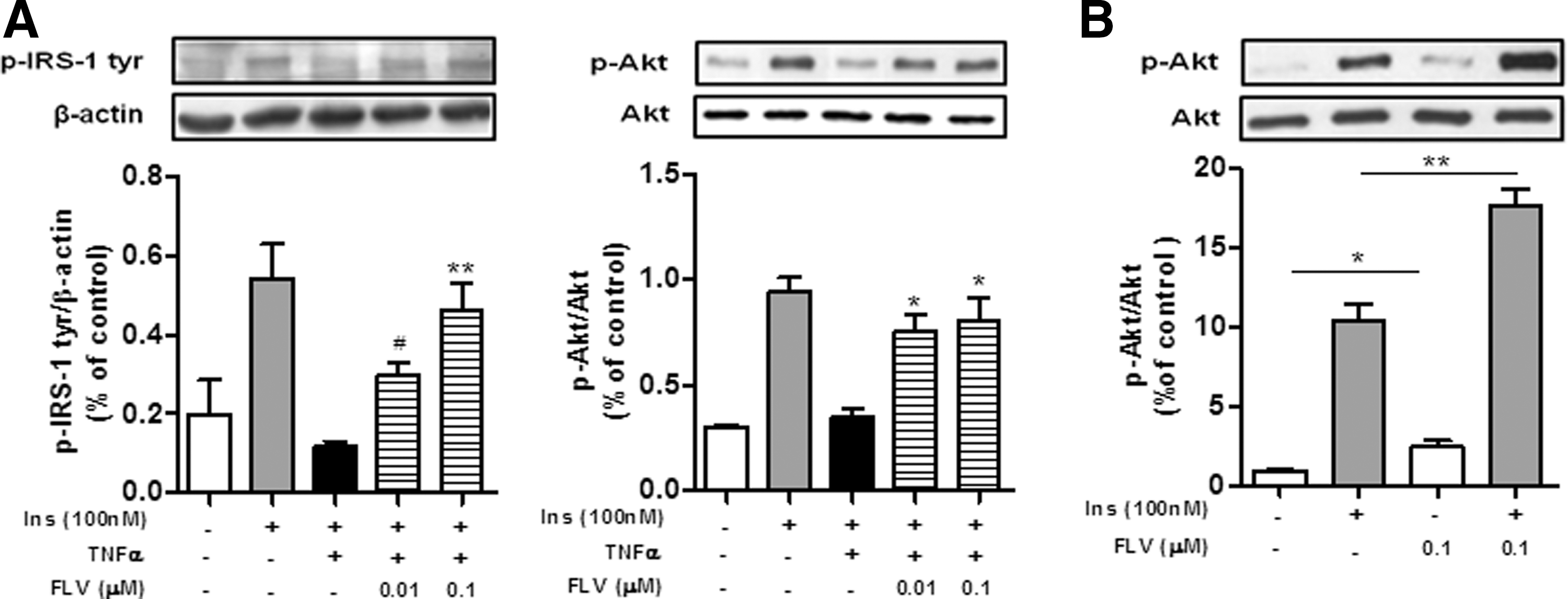
Soy peptide FLV improves insulin sensitivity in 3T3-L1 adipocytes.
Expression of peptide transporters in adipose tissue and adipocytes
We examined whether peptide transporters were expressed in adipose tissue or adipocytes. The mRNA and protein of two representative peptide transporters, PepT1 and PepT2, were measured by real-time PCR and western blotting. Jejunum and lung tissues were used as positive controls for PepT123 and PepT2, 24 respectively. We found that both PepT1 and PepT2 were expressed in epididymal adipose tissue, and PepT1 protein was hardly detected in the adipose tissue, whereas PepT2 protein was highly expressed at levels almost equal to those measured in lung tissue (Fig. 4A, B). We also confirmed that PepT2 mRNA and protein were expressed in 3T3-L1 adipocytes, and the expression of PepT2 was upregulated by the obesity-related inflammatory factors, lipopolysaccharide (LPS) and TNFα (Fig. 4C): LPS caused a 25-fold increase in PepT2 mRNA, and TNFα a 1.5-fold increase in PepT2 protein. Moreover, we observed that transcript levels of the peptide transporters were higher in the epididymal adipose tissue from obese mice fed a high-fat diet (HFD) than in tissue from lean mice (Fig. 4D).

Peptide transporters are expressed in adipose tissue and 3T3-L1 adipocytes.
Peptide transporter inhibitor disrupts the anti-inflammatory action of soy peptide FLV
To test whether peptide transporters function in adipocytes, we first tested the uptake of Ala–Lys–AMCA, which is a specific fluorescent substrate for peptide transporters. 25 The uptake of Ala–Lys–AMCA occurred in a concentration-dependent manner in adipocytes (data not shown). Moreover, the uptake was diminished by GlySar, which is a competitive substrate commonly used for peptide transporters 26 as well as by FLV (Fig. 5A), suggesting that FLV shares the peptide transporters with GlySar in the adipocytes.

The anti-inflammatory effect of soy peptide FLV is blocked by a peptide transporter inhibitor.
Next, we examined whether the peptide transporters participated in FLV uptake. FLV decreased the release of MCP-1 and IL-6 from adipocytes, and this decrease was inhibited by GlySar. GlySar alone did not affect release of the inflammatory cytokines from the adipocytes (Fig. 5B). These results suggest that FLV is transported into adipocytes through the peptide transporters, where it inhibits TNFα-induced inflammatory signaling.
Discussion
Obesity-induced adipose inflammation is characterized by hypertrophy of adipocytes, and the accumulation and activation of adipose macrophages. In obesity both hypertrophic adipocytes and adipose macrophages release large quantities of inflammatory cytokines, such as TNFα, IL-6, and MCP-1, which augment adipose inflammation and increase insulin resistance, resulting in the development of metabolic diseases such as insulin resistance. 3,27,28 In particular, TNFα (a representative inflammatory cytokine) plays a critical role in triggering and amplifying the inflammatory signaling cascade in adipose macrophages, and also inhibits insulin signaling in hypertrophic adipocytes in obese adipose tissue. 12,13 In this study, we confirmed that TNFα enhances the production of inflammatory cytokines (MCP-1, IL-6, and TNFα) from lipid-containing mature adipocytes. Importantly, we found that soy peptide FLV reduced the release of TNFα-induced inflammatory cytokines from both adipocytes and cocultured adipocytes/macrophages. Moreover, FLV also restored TNFα-induced insulin resistance in the adipocytes. Given that TNFα plays a major role in obesity-induced adipose inflammation and insulin resistance, these findings suggest that the soy peptide FLV has the potential to protect against metabolic disorders linked to obesity-induced inflammation.
In most cases, dietary protein-derived substrates are transported from the intestinal lumen by peptide transporters, which are membrane proteins that mediate the cellular uptake of di- or tripeptides. The peptide transporter PepT1 is mainly expressed on the apical membrane of epithelial cells in the small intestine 23 and it mediates the cellular uptake of di- and tripeptides, 18,29,30 whereas PepT2 is found in a variety of tissues, including lung and kidney as well as in nonepithelial cells. 24,31 Interestingly, recent studies have shown that the tripeptide Lys–Pro–Val (KPV) from the C-terminal end of α-melanocyte-stimulating hormone reduces intestinal inflammation, such as colitis (which is accompanied by enhanced recruitment of various activated immune cells) in a PepT1-mediated manner. 18,29 Moreover, our previous study showed that the soy peptide VPY elicited PepT1-mediated anti-inflammatory activity in vitro, and ameliorated chemically induced intestinal inflammation in vivo. 30 Moreover, PepT1 and PepT2 genes and proteins are detected in murine macrophages and lymphocytes, 32 and also in human monocytes/macrophages and T cells, 33 indicating that the soy peptide VPY reduces immune cell-mediated inflammatory responses during colitis through the peptide transporters. However, the involvement of the peptide transporters in controlling the inflammatory responses of adipocytes and/or macrophages in obesity-induced adipose inflammation has not yet been explored.
Intriguingly, we detected PepT1 and PepT2 mRNA in epididymal adipose tissue, and their mRNA transcript levels were upregulated in HFD-fed obese mice, indicating an association with obesity. PepT1 protein was highly expressed in jejunum, but was only present in low levels in epididymal adipose tissue. PepT2 protein, however, was predominantly expressed in adipose tissue, and both mRNA transcript and protein were well-represented in adipocytes. More interestingly, in adipocytes, the transcript and protein levels of PepT2 were markedly upregulated in the presence of obesity-related inflammatory factors such as TNFα and LPS. This indicates that PepT2 has a major role in facilitating the uptake of the soy peptide FLV in adipocytes in obesity, and that the soy peptide FLV may be a useful treatment for advanced adipose inflammatory conditions. Furthermore, we found that the anti-inflammatory effect of soy peptide FLV in adipocytes as well as in cocultured adipocytes/macrophages was blocked by GlySar, which is a competitive substrate for peptide transporters. 26 Given that PepT2, which has a higher affinity for tripeptide than PepT1, 33 is also highly expressed on macrophages, which are increased in obese adipose tissue, the beneficial effect of the soy peptide FLV against obesity-induced adipose inflammation may probably be attributed to PepT2.
It is well established that the TNFα-induced inflammatory signaling pathway includes phosphorylation of JNK and IKK, and degradation of IκBα. 13,22 In this study, we found that FLV suppressed the phosphorylation of JNK and IKK and degradation of IκBα in adipocytes, indicating that the decrease in inflammatory cytokines in its presence may be due to its inhibitory action on inflammatory signaling. Additionally, TNFα is known to suppress tyrosine phosphorylation of IRS-1 and, thus, to interfere with downstream insulin signaling. 13 Indeed, we confirmed that TNFα reduced the phosphorylation of both IRS-1 and Akt in adipocytes. Of note, FLV restored insulin responsiveness in a dose-dependent manner, and directly enhanced the phosphorylation of Akt. Taken together, these data indicate that FLV protects against TNFα-mediated insulin resistance by inhibiting inflammatory signaling, and might also have the potential to improve insulin sensitivity in adipocytes. Further studies are required to clarify the in vivo effects of FLV in obesity-induced adipose inflammation and insulin resistance.
In conclusion, we have demonstrated for the first time that FLV can be transported into adipocytes by PepT2, where it both decreases inflammatory responses, by blocking TNFα-induced inflammatory signaling, and improves insulin responsiveness (Fig. 6). Soy peptide FLV, therefore, may prevent obesity-induced adipose inflammation and insulin resistance.
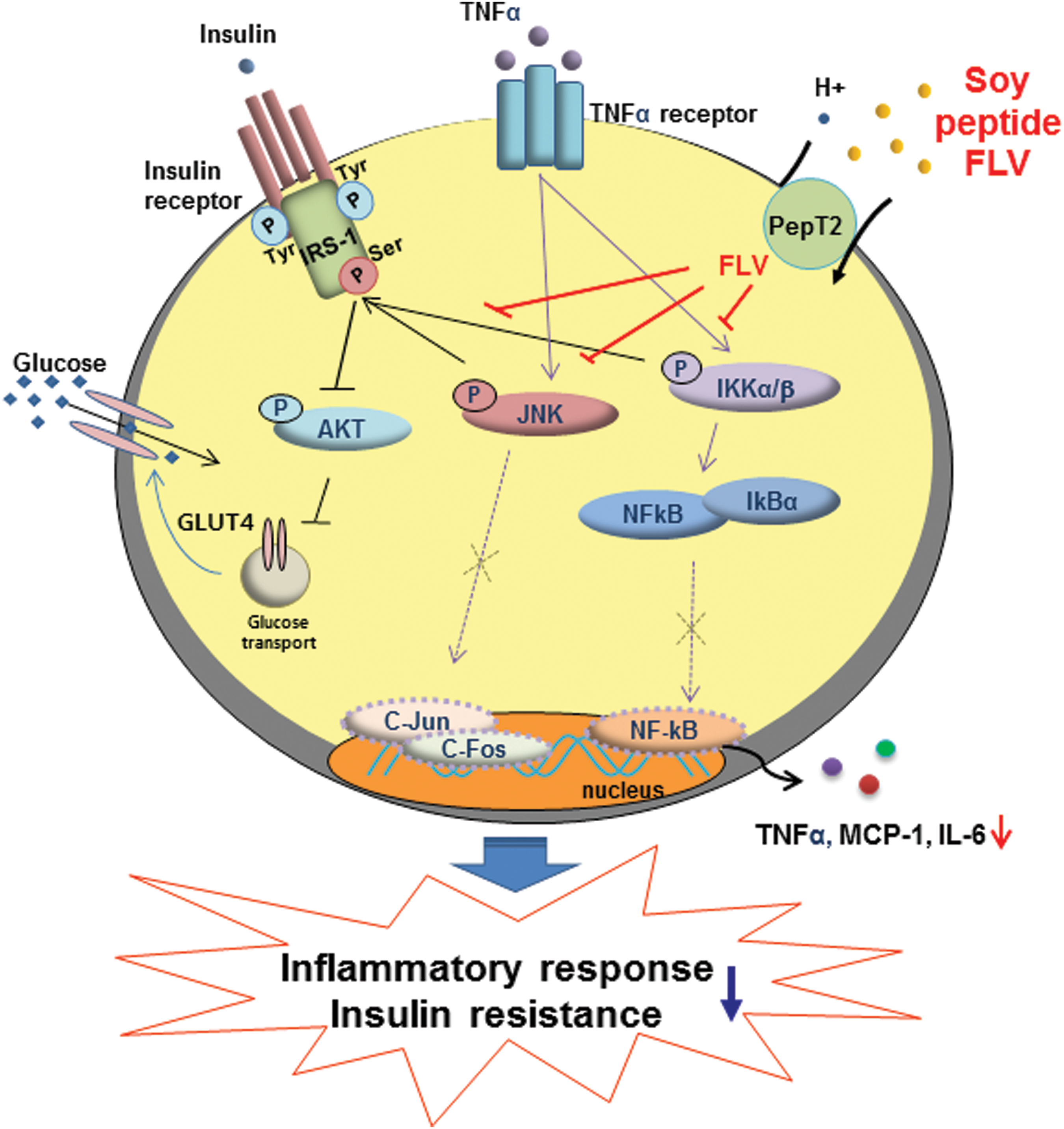
Schematic presentation of soy peptide FLV action on adipocytes. Soy peptide FLV reduces TNFα-induced inflammatory signaling pathway (JNK, IKK, and IκBα degradation) in adipocytes, and protects against TNFα-mediated insulin resistance. Color images available online at
Footnotes
Acknowledgments
This work was supported by the Science Research Center program (Center for Food & Nutritional Genomics Research: Grant no. 2015R1A5A6001906) of the National Research Foundation of Korea funded by the Ministry of Education, Science, and Technology and by Fuji Foundation for Protein Research.
Author Disclosure Statement
No competing financial interests exist.