
Abstract
Even though rice hull has various physiological functions with high antioxidant potential, the molecular mechanism(s) underlying the effects of rice hull on benign prostatic hyperplasia (BPH) have not been evaluated. The aim of this study was to determine the protective effect of rice hull water extract (RHE) against BPH, which is a common disorder in elderly men and involves inflammation that induces an imbalance between cell proliferation and cell death. In this study, RHE-treated mice exhibited lower prostate weights and ratios of prostate weight to body weight compared to those for the BPH-induced group. In addition, RHE-treated mice had lower serum levels of dihydrotestosterone, mRNA expression of 5α-reductase2, and protein expressions of proliferating cell nuclear antigen (PCNA). Furthermore, RHE treatment significantly decreased cell proliferation by regulating the expression levels of inflammatory-related proteins (iNOS and COX-2) and apoptosis-associated proteins (Fas, FADD, procaspase-8, −3, and Bcl-2 family proteins). These results suggest that RHE could protect against the development of BPH through its anti-inflammatory and apoptotic properties and has good potential as a treatment for BPH.
Introduction
B
Although aging and androgens are the two established risk factors for the development of BPH, recent novel findings have also highlighted the key role of inflammation and apoptosis-related mediators. 5 The inflammation present in BPH may contribute to tissue injury, and the cytokines produced by inflammatory cells may serve to drive local growth factor production and angiogenesis in the tissues as a wound healing response. 6 This complex proinflammatory microenvironment is closely related to BPH stromal hyperproliferation. All of the available data seem to suggest that the inflammation-induced damage of the prostatic tissue leads to a chronic process of wound healing that activates hyperproliferative processes resulting in the cyclic reactivation of prostatic inflammation. In addition, cellular hyperplasia and changes in tissue consistency in BPH are accompanied by a decrease in the levels of cell apoptosis, which play a key role in the maintenance of the size of the prostate. As demonstrated by immunohistochemistry analysis of biopsies from BPH patients, apoptotic cells, as assessed by terminal deoxynucleotidyl transferase dUTP nick-end labeling, are rarely observed in the stroma, while stromal cells express high levels of proliferation markers. 7 Downregulation of the apoptotic factor transforming growth factor-beta, in combination with increased levels of Bcl-2, a potent apoptosis suppressor, contributes to hyperproliferation of the prostate epithelium. 8 As a result of the disruption in apoptosis and proliferation dynamics within the prostate, the gland becomes hypertrophic and eventually BPH symptoms develop.
Rice (Oryza sativa L.) is a major source of nourishment for the world's population, especially in Asia. Worldwide production of rice is estimated at around 680 million tons, equivalent to that of wheat. Rice is composed of a white starchy rice kernel tightly coated with bran enclosed in a tough siliceous hull. 9 Rice hulls consist mainly of lignin, hemicellulose, cellulose, and hydrated silica. 10 Rice hull extractives have been proven to possess scavenging effects on the free electrons of reactive oxygen species, metal-chelating activity, and a protective effect against oxidative DNA damage in human lymphocytes. 11 In addition, rice hull has been demonstrated to have antimicrobial, antibacterial, antiallergic, and anti-inflammatory properties. 10 Furthermore, rice hull extractives exhibited cytotoxicity against human cancer cells and protective effect against type 2 diabetes in mice induced by a high-fat diet.
Although studies on the physiological functions of rice hull have been performed, the molecular mechanism(s) underlying the effects of rice hull on BPH have not been evaluated. In this study, we evaluated the effect of rice hull on BPH and demonstrated that rice hull water extract (RHE) could modulate the imbalance between prostatic cell growth and apoptosis in a rat model of testosterone-induced BPH.
Materials and Methods
Materials
Testosterone, phenylmethylsulfonylfluoride, Triton X-100, propidium iodide, Nonidet P-40, and protein inhibitor cocktail were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Dimethyl sulfoxide was purchased from Junsei Chemical Co., Ltd. (Tokyo, Japan) and finasteride (≥98%) was obtained from Merck & Co., Inc. Oligonucleotide primers for 5α-reductase 2 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were purchased from Bioneer (Daejeon, Republic of Korea), and SYBR Premix Ex Taq was purchased from Takara (Shuzo, Shiga, Japan). Antibodies for inducible nitric oxide synthase (iNOS; M-19), cyclooxygenase-2 (COX-2; C-20), Fas (B-10), caspase-8 (C-20), caspase-3(E-8), Bcl-2 (C-2), Bcl-xL (H-5), Bax (B-9), and β-actin (ACTBD11B7) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibodies for proliferating cell nuclear antigen (PCNA) and Fas-associated protein with death domain (FADD) were purchased from BD Biosciences, Pharmingen (San Diego, CA, USA). Phosphate-buffered saline (PBS) was obtained from Life Technologies, Inc. (Grand Island, NY, USA).
Preparation of the water extract of rice hull
Rice hulls (300 g) were weighed and soaked with 3 L water and left overnight. Then, the rice hull was boiled for 1 h and the water extract was filtered through Whatman No. 1 filter paper. The filtrate was evaporated to dryness under reduced pressure on an EYELAN-1000 rotary evaporator (EYELA Riakikai Co., LTD., Tokyo, Japan) at 40°C. Finally, 30 g of water extract (at a concentration of 1 g/mL) was obtained from 300 g of rice hull and stored at −20°C for later use.
High-performance liquid chromatography analysis
The chromatographic system consisted of a Jasco HPLC-LC-2000 Plus (Tokyo, Japan) equipped with a Jasco MD-2018 Plus Photodiode Array Detector, using the INNO Column C18 (5 μm, 4.6 × 250 mm; Young Jin Biochrom Co. Ltd.). The column temperature was set to 30°C. The mobile phase consisted of acetonitrile (solvent A) and water containing 0.05% acetic acid (solvent B). The mobile phase protocol specified 20% of solvent A for 60 min. The total running time was 60 min and the flow rate was 0.5 mL/min. Data acquisition was performed in the range of 190–650 nm. The retention times of 4-hydroxybenzoic acid were obtained at 12.34 min.
Gas chromatograph–mass spectrometer analysis
The gas chromatograph–mass spectrometer (GC-MS) analysis was performed on an Agilent 6890N GC system interfaced with a 5975i MS spectrometer. The ion source temperature was 200–290°C. Each sample was injected in split mode (30:1) at 280°C and separated through a capillary column of DB-5MS UI (30 m, 0.25 mm, 0.25 μm). The initial oven temperature was 80°C, which was increased to 300°C at 10°C/min. Carrier gas (He) flow was 1 mL/min.
Animals and treatment
Ten-week-old male Sprague-Dawley rats (200 ± 20 g) were purchased from Daehan Biolink (Daejeon, Korea). The animals were housed under conditions that were in accordance with the guidelines for the care and use of laboratory animals, adopted and promulgated by the Institutional Animal Care Committee of Sangji University (Reg. No. 2014-20). The rats were acclimatized to the laboratory conditions for 2 weeks before starting the experiment. They were provided free access to food and water for 4 weeks and were maintained under a 12-h light/12-h dark cycle at a constant temperature (22°C ± 2°C) and relative humidity (55% ± 9%) throughout the experiment. The rats were randomly assigned to one of four groups (n = 6): CON group (the normal control group with normal prostate and treated with a vehicle solution of PBS 200 μL; p.o.); BPH group (BPH-induced group treated with vehicle solution of PBS 200 μL; p.o.); Fina group (BPH-induced group treated with finasteride, 5 mg/kg/day; p.o.); and RHE group (BPH-induced group treated with RHE, 200 mg/kg/day; p.o.). The rats in the control group were operated upon and then sewn up without removing the testicles after anesthetization with Zoletil 50 (20 mg/kg, i.p.); the rats in the other groups were castrated. After 3 days of recovery, BPH-induced groups were injected with testosterone propionate (10 mg/kg/day; i.p.; dissolved in corn oil) alone or along with RHE or finasteride daily for 4 weeks. Twenty-four hours after the last administration, all rats were sacrificed after anesthetization with Zoletil 50 (20 mg/kg, i.p.). Blood samples were drawn from the caudal vena cava and serum was separated by centrifugation and stored at −80°C. The entire prostate gland was removed and weighed. 12
Prostate weight to body weight ratio
Prostatic tissues were excised, rinsed, and weighed immediately. Prostate weight to body weight ratio (PW/BW ratio) was calculated as described below.
PW/BW ratio = (prostate weight of each animal from experimental group/body weight of each animal from experimental group) × 1000.
Growth inhibition of prostate weight
On study completion, the prostates were removed and weighed. The percentage of growth inhibition was calculated as follows: 100− [(treated group − control group)/(BPH group − control group) × 100].
Serum DHT analysis
Serum concentrations of DHT were determined using a commercial enzyme-linked immunosorbent assay (ELISA; ALPCO Diagnostics, Salem, NH, USA). Assays were performed according to the manufacturer's instructions.
Histological analysis
The prostatic tissue from the rats in each group was fixed in 4% buffered formalin, embedded in paraffin, and cut into 4-μm-thick sections. The sections were stained with hematoxylin and eosin before histological examination. Images were acquired using an SZX10 microscope. Histological analysis was performed and the thickness of epithelium tissue from prostate (TETP) was measured by professional pathologists (Seo-Na Jang and Jae-Hak Park) from Korean Experimental Pathology, Inc. (Seoul, Republic of Korea) (sample number: XP13-0087).
Quantitative real-time polymerase chain reaction analysis
Prostatic tissue from each animal was homogenized, and total RNA was isolated using Easy-Blue Reagent (Intron Biotechnology Inc., Gyeongi-do, Republic of Korea) according to the manufacturer's instructions. Total RNA was quantified using an Epoch micro-volume spectrophotometer system (BioTek Instruments, Inc., Winooski, VT, USA). Total RNA from the prostate sample was converted to cDNA using a high-capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA, USA). PCR amplification was performed in the presence of SYBR Green (Applied Biosystems). The oligonucleotide primers for rat5α-reductase2 were ATG GGG ACC CTG ATC CTG TG (forward) and CGA CAC AAA GGA AGG CA (reverse), and those for rat GAPDH, analyzed as a housekeeping gene, were TGA TTC TAC CCA CGG CAA GT (forward) and AGC ATC ACC CCA TTT GAT GT (reverse). Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using a thermocycler (Gene Amp PCR system 9700; Applied Biosystems); the results have been expressed relative to those of GAPDH.
Western blot analysis
Prostatic tissue from each animal was homogenized in a commercial lysis buffer (PRO-PREP; Intron Biotechnology, Inc.) and incubated for 25 min on ice to induce cell lysis. Tissue extracts were centrifuged at 16,000 ×g at 4°C for 20 min, and the supernatants were transferred to clean tubes. The protein concentration was determined using the Bio-Rad protein assay reagent according to the manufacturer's instructions (Bio-Rad, Hercules, CA). Aliquots of each protein sample (30 μg) were separated on a sodium dodecyl sulfate–polyacrylamide gel and transferred onto a polyvinylidene fluoride membrane. Membranes were blocked with 5% skimmed milk at 4°C for 1 h. The membranes were incubated overnight with primary antibodies, which included the anti-PCNA, anti-iNOS, anti-COX-2, anti-Fas, anti-FADD, anti-caspase-8, anti-caspase-3, anti-Bcl-xL, anti-Bax, and anti-β-actin. Blots were washed thrice with Tween-20/Tris-buffered saline (TTBS), followed by incubation with the corresponding secondary antibody (Santa Cruz Biotechnology) for 1 h at room temperature. Blots were again washed thrice with TTBS, and immunoreactive protein bands were visualized using enhanced chemiluminescence and exposure to X-ray film (Amersham, Piscataway, NJ, USA).
Statistical analysis
All values are expressed as means ± the standard errors of the means (SEM) for data from six rats. The data were analyzed using analysis of variance with Dunnett's test. Statistical analysis was performed using the Statistical Package for the Social Sciences (SPSS, version 19.0; IBM Corporation, Armonk, NY, USA).
Results
High-performance liquid chromatography and GC-MS analysis of RHE
To confirm the constituents of RHE, high-performance liquid chromatography (HPLC) with a pulsed amperometric detector (HPLC-PAD) and GC-MS analyses were performed. HPLC-PAD measurement of RHE demonstrated various chromatographic peaks. Comparing the chromatographic peaks of RHE with reference chromatographic peaks, 4-hydroxybenzoic acid was identified (Fig. 1A). When RHE was analyzed using a GC-MS and compared with a database, 2-methoxyphenol, 4-vinylphenol, 2-methoxy-4-vinylphenol, 4-hydoxybenzaldehyde, and 2,6-dimethoxyphenol were detected (Fig. 1B).
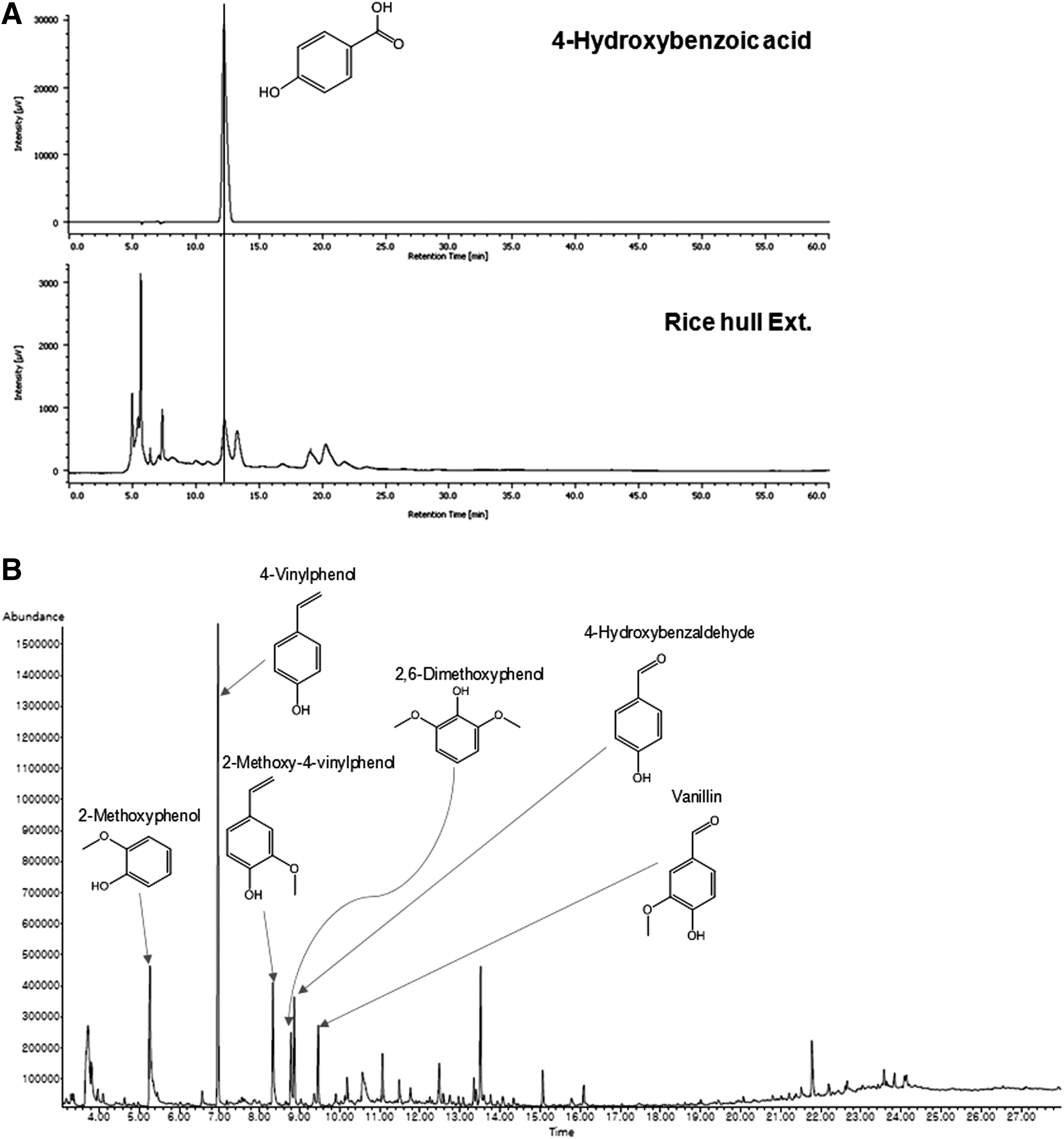
HPLC-PAD and GC-MS analysis of RHE.
Effects of RHE on prostate weight in the BPH-Induced rats
As shown in Figure 2A, the testosterone-induced BPH group showed a significant increase in prostate weight compared with the control group, which suggested that testosterone successfully induced BPH in the castrated rats. However, the finasteride- and RHE-treated groups showed a significant trend for reduction in the prostate weight (Fig. 2B).
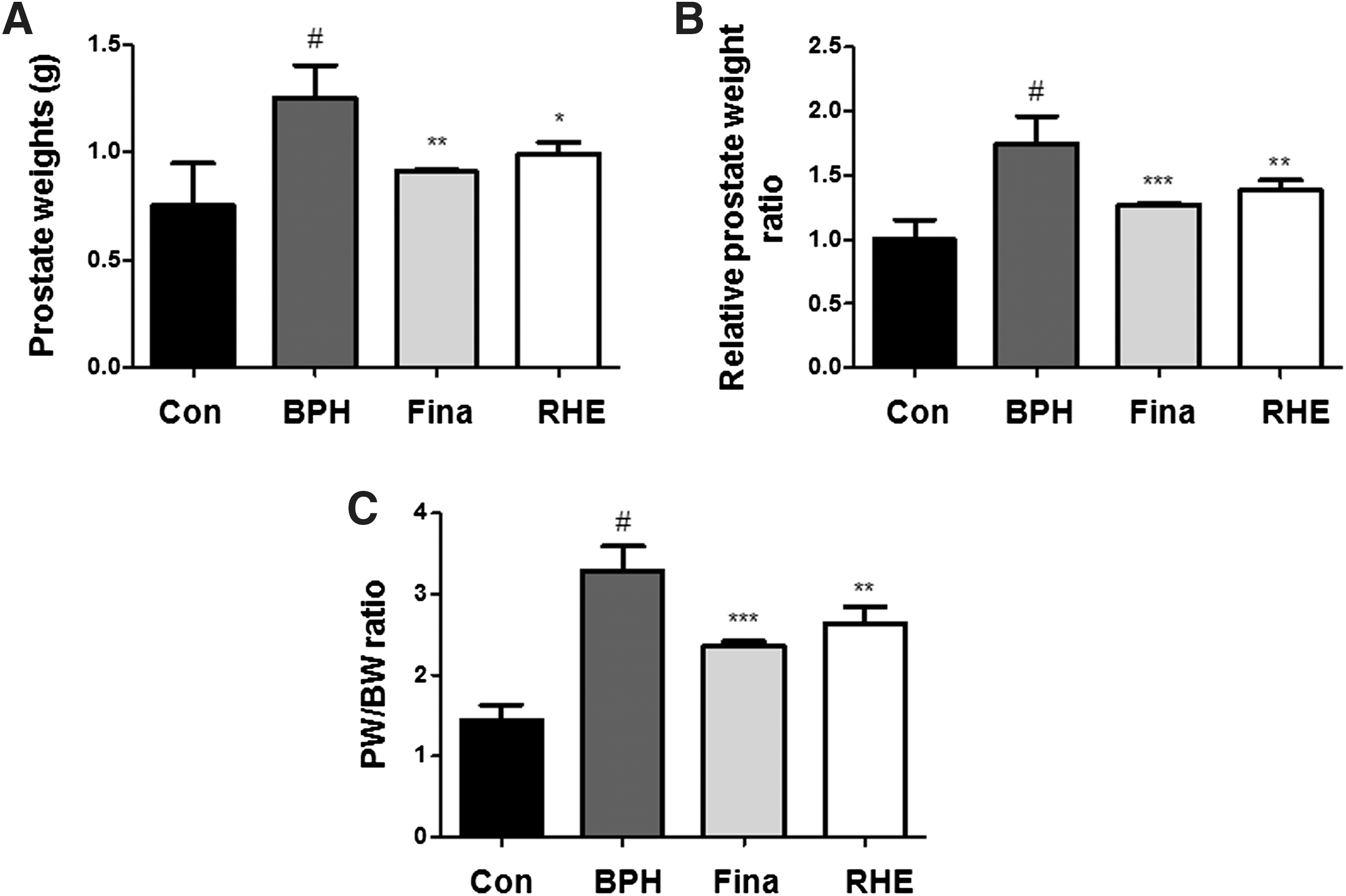
Effect of RHE administration on prostate weight in BPH-induced rat models. Changes in prostate total weight
Prostate weights and prostate weight to body weight ratios (PW/BW) are important indicators in the development of BPH. The PW/BW ratio in BPH-induced group was 2.27 times higher than that in the control groups (Fig. 2C). In the finasteride- and RHE-treated groups, the PW/BW ratio was 1.63 and 1.82 times higher, respectively, compared with the control group. The growth inhibition ratio of prostates in each group is shown in Table 1. In the finasteride- and RHE-treated groups, the growth inhibition ratio of prostates was 63.98% ± 1.86% and 48.45% ± 1.26%, respectively.
P < .05 versus Con group.
P < .05.
P < .01 versus BPH group.
Effects of RHE on serum DHT levels and 5α-reductase 2 mRNA expression in BPH-Induced rats
Prostatic luminal secretory cells require androgens, particularly the intracellular metabolite of testosterone, DHT, for terminal differentiation and secretory functions. DHT is generated from testosterone by 5α-reductase, which is present in fibroblasts of the stroma and in basal epithelial cells age. 13,14 As shown in Figure 3A, serum DHT levels in the testosterone-induced BPH group were significantly higher than those in the control group. However, serum DHT levels in the finasteride-treated group and the RHE-treated group were significantly decreased compared with the BPH-induced group. To determine whether this effect of RHE was associated with altered levels of the DHT from testosterone by 5α-reductase in prostatic tissue, the expression levels of 5α-reductase 2 mRNA were examined. As shown in Figure 3B, similar to the results in the finasteride-treated group, RHE treatment significantly decreased testosterone-induced 5α-reductase 2 mRNA levels in the prostate tissue of BPH-induced rats. These findings suggest that the inhibition of 5α-reductase 2 mRNA by RHE might be caused by the suppression of 5α-reductase 2 activity.

Effect of RHE administration on the serum DHT production and mRNA level of 5α-reductase 2 in prostate tissues of BPH-induced rat models.
Effects of RHE on histological analysis and cell proliferation in BPH-Induced rats
In histological analysis, one layer of low columnar epithelial cells formed a secretory lumen that was filled with thin acidophilic materials in the control group (Fig. 4A). In contrast, the epithelial cells in the BPH-induced rats were arranged in several uneven layers, and the gland was excessively developed. However, finasteride and RHE treatment suppressed these typical hyperplastic patterns, which represent the histologic change of normal prostatic tissue into tissue with prostatic hyperplasia. The luminal volume was increased and the glandular epithelial height was markedly reduced in these groups. In addition, as shown in Figure 4B, TETP analysis showed that the most thickened epithelium tissue was observed in BPH-induced rats and the finasteride- and RHE-treated groups showed a significant reduction in the thickness of the epithelium tissue of the prostate.

Effect of RHE administration on the prostatic cell proliferation. Hematoxylin and eosin staining of prostatic tissue from BPH-induced rat models
To evaluate the effects of RHE on the proliferation of prostatic epithelial cells, we examined the mRNA and protein expression levels of PCNA, which is a proliferation marker, in the prostatic tissue of BPH-induced rats. As shown in Figure 4C and D, qRT-PCR and Western blot analysis indicated an increase in PCNA mRNA and protein levels in the BPH-induced group compared to the levels in the control group. Compared to the BPH-induced group, however, the finasteride- and RHE-treated groups exhibited a lesser increase in the mRNA and protein levels of PCNA, indicating antiproliferative effects in BPH.
Effects of RHE on inflammatory proteins in BPH-induced rats
Inflammation can influence the tissue microenvironment through the COX activity and nitric oxide synthesis, all linked to the cellular proliferation in prostate tissue. 15 Inflammatory-related factors can alter protein structure and function, induce gene changes, cause posttranslational modifications, including those involved in the DNA repair and apoptosis processes, and provoke prostatic cellular proliferation. As shown in Figure 5, treatment with testosterone markedly increased the protein expression levels of iNOS and COX-2 in the BPH-induced group compared to the control group. The finasteride- and RHE-treated groups, however, exhibited reduced expression levels of these inflammatory proteins.

Effect of RHE administration on the expression of inflammation proteins in prostate tissues of BPH-induced rat models. The expression levels of iNOS proteins and COX-2 proteins were determined by Western blotting using specific antibodies. β-actin was used as internal controls. Densitometric analysis was performed using Bio-Rad Quantity One Software. The data shown represent mean ± SEM of six rats per group. # P < .05 versus Con group; *P < .05, **P < .01, ***P < .001 versus BPH group.
Effects of RHE on caspase-8 and Bcl-2-mediated apoptosis in BPH-Induced rats
Apoptosis is a selective process of physiological cell deletion that plays an important role in the balance between cellular replication and death. 16 In this study, treatment with testosterone induced downregulation of the protein expression levels of Fas and FADD and upregulation of procaspase-8 and procaspase-3 in the BPH-induced group (Fig. 6A). Treatment with RHE, however, markedly restored Fas and FADD protein expression and suppressed procaspase-8 and −3 protein expression, suggesting the involvement of caspase-8-mediated extrinsic apoptotic pathways. In addition, the RHE-treated group showed decreased protein levels of the Bcl-2 and Bcl-xL (antiapoptotic proteins), but increased expression of Bax (proapoptotic protein) compared to that in the BPH-induced group (Fig. 6B). Thus, the ratio of Bcl-2 to Bax significantly decreased after treatment with RHE suggesting the involvement of the Bcl-2 family of proteins (Fig. 6C), which is an important mediator of the intrinsic apoptotic pathway, in RHE-induced apoptosis during BPH. These findings suggested that RHE-induced apoptosis was mediated by the regulation of the expression levels of the death receptor-related proteins, such as FADD, procaspase-8, and Bcl-2 family of proteins.

Effect of RHE administration on the formation of DISC and regulation of Bcl-2 family proteins in prostate tissues of BPH-induced rat models.
Discussion
BPH is thought to be a disease with multiple etiologies, including hormone signaling, disruption of proliferation and apoptosis dynamics, and chronic inflammation with changes in the morphology and phenotype of the prostate stroma. 17 BPH arises in the periurethral and transitional zones of the prostatic gland and represents an inescapable phenomenon for the ageing male population. BPH is uncommon before the age of 40 years; approximately 50% of men develop BPH-related symptoms by 50 years of age. The incidence of BPH increases by 10% per decade and reaches 80% at approximately 80 years of age. 18 Changes in the concentration of androgens in the prostate during development, puberty, adult life, and ageing may contribute to the development of BPH. Higher levels of DHT are associated with the occurrence of BPH and a higher testosterone to DHT ratio is associated with a decreased risk of BPH. 19 DHT is the product of conversion of testosterone by the enzyme 5α-reductase and is produced in the tissues of the liver, skin, and organs that originate from the mesonephric duct, such as the prostate. Within the prostate, locally produced DHT acts in a paracrine manner to stimulate growth. Inhibitors of 5α-reductase decrease production of DHT within the prostate, which results in decreased prostate volume, increased peak urinary flow rates, and improvement in symptom scores. 1 Our investigation was performed on a BPH model in rats induced by subcutaneous injection of testosterone after castration. The model rats exhibited enlargement of the prostate as a consequence of progressive hyperplasia of glandular and stromal tissues around the urethra, clearly confirming the influence of androgens on prostate growth. Treatment with RHE decreased the prostate weight by inhibiting the growth of prostatic cells. Moreover, the administration of RHE reduced serum DHT levels and mRNA levels of 5α-reductase 2.
In fact, hormonal imbalance has been proposed to provoke intraprostatic inflammation that may trigger a compensatory cellular proliferation with the risk of hyperplastic growth. 20 The prostate is normally populated by small numbers of T cells, B lymphocytes, macrophages, and mast cells. 21 Interestingly, several studies have shown that the prostatic tissue in BPH patients contains disseminated infiltration of T and B lymphocytes and numerous colonies of macrophages. The immune response in the prostate is primarily T-cell mediated, with regulatory T cells (CD4) in the stroma and cytotoxic T cells (CD8) in the epithelium. 22 In addition, by analyzing the expression of T-cell activation markers, it was demonstrated that such inflammation mediators remain chronically activated. 21 Because local accumulation of activated lymphocytes can cause tissue destruction, the high concentrations of cytokines and consequent tissue rebuilding might contribute to the pathogenesis of BPH. Indeed, upregulation of a set of proinflammatory cytokines has been reported in BPH tissue, particularly interleukin-15 (IL-15) in stromal cells, IL-17 in infiltrating T cells, interferon-γ in basal and stromal cells, and IL-8 in epithelial cells. 23,24 Based on this inflammatory response, the microenvironment also induces iNOS and COX-2 expression in the epithelial cells in BPH. 25 In this study, testosterone increased the level of iNOS and COX-2 protein expression in BPH-induced rats. However, the increased level was reversed after treatment with RHE. Therefore, the anti-BPH effects of RHE may be related to its anti-inflammatory properties.
The development of BPH is associated with the enhanced proliferation and suppression of apoptosis in prostatic cells 7 . Apoptosis, a morphologically and biochemically defined form of cell death, plays a key role in the regulation of growth of normal and neoplastic tissues in the prostate. Recent studies have amply documented that two major pathways are involved in the regulation of apoptosis. The extrinsic apoptotic pathway involves death-inducing signal complex, including cell surface death receptors such as Fas/CD95 and tumor necrosis factor receptors 1 and 2 that, upon activation, upregulate downstream signaling cascades leading to the accumulation of FADD and the activation of caspase-8. 26 The intrinsic pathway is dependent on various cell stress stimuli leading to an altered ratio of Bcl-2 family members, affecting the release of cytochrome c and apoptotic protease activating factor-1 that leads to caspase-9 activation. The active forms of caspase-8 and caspase-9 may activate the downstream effector caspase-3, resulting in the cleavage of poly-ADP ribose polymerase-1 and leading to DNA fragmentation. In this study, we showed that RHE markedly increased the protein expression of Fas and FADD, whereas it decreased the protein expression of procaspase-8 and procaspase-3 in BPH-induced rats. Moreover, the ratio of Bcl-2 to Bax significantly decreased after treatment with RHE, suggesting the involvement of the Bcl-2 family of proteins, important mediators of RHE-induced apoptosis during BPH. Based on these data, we concluded that RHE-induced apoptosis plays a role in the effect of this extract on BPH and that this was mediated through the extrinsic and intrinsic apoptotic pathways.
In summary, our results demonstrate that RHE treatment prevents BPH development (based on prostate weight and determination of DHT production) and it significantly reduces 5α-reductase and PCNA expressions in BPH-induced animal models. In addition, RHE treatment inhibits the expression of iNOS and COX-2, and procaspases and Bcl-2 family proteins. Taken together, these results suggest that RHE treatment had a protective effect on BPH development by regulating inflammatory-related proteins and apoptosis-modulated proteins. RHE treatment provides a promising therapeutic means for treating BPH.
Footnotes
Acknowledgment
The GC-MS analysis was performed with the 6890N GC/5975i MS system at Korea Basic Science Institute.
Author Disclosure Statement
No competing financial interests exist.