
Abstract
We have provided a protocol for establishing an atopic dermatitis (AD) in vitro model, and evaluated the effects of Costaria costata (CC) extracts on AD in an in vitro model using keratinocytes and splenocytes from AD-induced mice and mast cells. HaCaT cells were each treated with 200 μg/mL of CC water extract (CCW), CC 10% ethanol extract (CCE10%), and CC 70% ethanol extract (CCE70%), immediately followed by stimulation with 20 ng/mL tumor necrosis factor (TNF)-α, and 20 ng/mL interferon (IFN)-γ for inflammation. The splenocytes from AD-induced mice were each treated with 200 μg/mL of CCW, CCE10%, and CCE70%, followed by stimulation with 5 μg/mL ConA or lipopolysaccharide (LPS), to induce T cell or B-cell activation, and 5 μg/mL LPS and 50 ng/mL interleukin-4, to induce immunoglobulin (Ig) E production. We investigated the effects of CCW, CCE10%, and CCE70% on the production of histamine in PMA, and A23187-stimulated MC/9 cells. We found that treatments with CC extracts decreased the production of proinflammatory cytokines in TNF-α and IFN-γ-stimulated HaCaT cells, and the suppression of the imbalance of Th1/Th2 cytokines and IgE production on primary splenocytes. In addition, CC extracts resulted in a decrease in histamine release in the PMA and A23187-simulated MC/9 cells. According to our present results, we can conclude that CC extracts may be effective for the treatment of other allergy diseases, and AD, via the attenuation of allergic reactions.
Introduction
T
Keratinocytes, the major cells of the epidermis, can produce proinflammatory cytokines and chemokines, such as CCL17/thymus, activation-regulated chemokine (TARC), and Eotaxin (CCL11) to attract leukocytes during the development of AD. 5 The persistent leukocytes infiltrate the skin tissue, inducing an inflammatory response, including T-cell activation by the dendritic cells (Langerhans cells) of the skin. 6 The activated CD4+ T cells (T helper cells, Th cells) secrete Th 2 cytokines, including interleukin (IL)-4 and IL-10, which help B cells to produce large amounts of immunoglobulin (Ig) E in the initiation phase of AD. 7,8 IgE can prime mast cell to increase their degranulation response by binding to receptors for IgE (FcɛRI), which can in turn release histamines and mediators, further potentiating the inflammation response in the development of AD. 8,9 Several reports have described that most of blood samples from AD patients displayed higher concentrations of IgE and histamine, compared with nonallergic controls. 10 Medicines used to treat AD, such as topical glucocorticoids and immunomodulators, are not always sufficiently responsive, and they can carry a risk of side effects; as such, many studies on the prevention of AD development have focused on herbal medicines as an alternative. 11
Recently, it was established that the biochemical molecules, dietary fiber, and minerals from marine algae can provide essential biologically active components, and holds antioxidative, antiobesity, antitumor, and anti-inflammatory potential. 12 –14 Costaria costata (CC), a type of benthic brown algae species, in the genus Costaria and the family Costariaceae is found in the seas of Korea, China, and Japan. 15 Although various marine algae are being studied, a few studies have reported that CC has effect on liver injury and cancer. 16,17 In this study, we investigated the protective effects of CC extracts on an AD in vitro model. Various in vivo model systems, including Picryl chloride (PiCl) or 1-Chloro-2,4-dinitrobenzene (DNCB)-induced, and an air-uncontrolled convention-induced NC/Nga mouse, to elucidate the pathogenesis of AD, have been used. 18,19 A few in vitro model systems have already studied the pathogenesis of AD, although our models have novel advantages, such as the reduction of animal usage, time savings, and identification of specific mechanisms. As such, we have provided a protocol for establishing an in vitro AD model, and investigated the effects of CC extracts on it.
Materials and Methods
Preparation of the extracts
CC was obtained from the Marine Biotechnology Research Center (Wando, Korea). CC was extracted using 60 volumes of water or 10% ethanol, 20 volumes of 70% ethanol at 80–100°C for 3 h in a reflux apparatus, and filtered. The extracts obtained after filtering (Whatman No. 6; Whatman, Maidstone, United Kingdom) were concentrated and lyophilized by rotary evaporator in vacuo. The dried CC water extract (CCW), CC 10% ethanol extract (CCE10%), and CC 70% ethanol extract (CCE70%) were kept in a tightly sealed, light-protected container at −20°C, until use.
Cell culture and treatments
HaCaT cells (keratinocytes) and MC/9 cells (mast cells) were obtained from the American type culture collection (ATCC, Manassas, VA, USA). The cells were cultured in Dulbecco's minimal essential medium (Hyclone Laboratories, Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS; Hyclone Laboratories), 100 mg/L of penicillin-streptomycin, and 2 mmol/L of glutamine (Hyclone Laboratories) at 37°C in a humid atmosphere of 5% CO2.
Determination of mRNA expression in HaCaT cells
HaCaT cells were seeded at a concentration of 1 × 106 cells/well in six-well tissue culture dishes, and incubated for 24 h. After 24 h incubation, the cells were each treated with 200 μg/mL of CCW, CCE10%, and CCE70%, immediately followed by stimulation with 20 ng/mL tumor necrosis factor-α (TNF-α) and 20 ng/mL interferon-γ (IFN-γ) for 24 h. Following this, the cells were lysed in the presence of RLT buffer (Qiagen, Valencia, CA, USA) including 1% β-mercaptoethanol. Total RNA was extracted from the cells using the RNeasy Mini kit (Qiagen). Two hundred nanograms of purified total RNA was to synthesize complementary DNA (cDNA) in 20 μL of reaction buffer, using the iScript™ cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA, USA). We used 1 μL cDNA with the SYBR Green PCR Master Mix (iQ SYBR Green Supermix, Bio-Rad Laboratories) for Real-Time PCR (Applied Biosystems, Foster City, CA, USA) assays. We then amplified the cDNA with 40 cycles of denaturation (95°C for 30 s), annealing (55°C for 30 s), and extension (72°C for 60 s), using the following primers: GAPDH forward primer 5′-ATG GAA ATC CCA TCA CCA TCT T-3′, reverse primer 5′-CGC CCC ACT TGA TTT TGG-3′; TARC forward primer 5′-GAA GAC GTG GTA CCA GAC ATC TGA-3′, reverse primer 5′-CCC TGC ACA GTT ACA AAA ACG A-3′; Eotaxin forward primer 5′-GCG ACT AGA GAG CTA CAG GAG AAT C-3′, reverse primer 5′-GGT CTT GAA GAT CAC AGC TTT CTG-3′; TNF-α forward primer 5′-CCA CTT CGA AAC CTG GGA TTC-3′, reverse primer 5′-TTA GTG GTT GCC AGC ACT TCA-3′; IL-6 forward primer 5′-AGG GCT CTT CGG CAA ATG TA-3′, reverse primer 5′-GAA GGA ATG CCC ATT AAC AAC AA-3′. The 7500 System SDS software version 1.3.1 (Applied Biosystems) was used to process the data calculation of relative quantitation.
Splenocytes primary culture
The present experimental protocols were approved by the Institutional Animal Care and Use Review Committee of Kyung Hee University (KHUASP(SE)-15-011). NC/Nga mice (22–25 g, 6 weeks, male) and age-matched Balb/c mice (22–25 g, 6 weeks, male) were purchased from SLC, Inc. (Kotoh-cho, Hamamatsu, Japan), and NC/Nga mice were maintained under conventional laboratory conditions, but without air filtration, to induce AD spontaneously. We measured the number of scratching episodes, and then the AD-induced mice were classified by scratching when over 50 in 20 min period. At the end of 10 weeks, all mice were killed, and the spleens of all the mice were dissected and gently placed in to a 40 mesh strainer in culture medium (RPMI 1640, 10% FBS, 2 mmol/L glutamine, 100 mg/L of penicillin-streptomycin). We resuspended the cell pellet in 2 mL of red blood cell lysing buffer (Sigma, St. Louis, MO, USA) at 37°C for 3 min, and washed twice with phosphate-buffered saline.
Determination of T and B-cell proliferation in primary splenocytes
The splenocytes were cultured at a concentration of 1 × 106 cells/well in 96-well tissue culture dishes, and then each treated with 200 μg/mL of CCW, CCE10%, and CCE70%, immediately followed by stimulation with 5 μg/mL Con A to induce T-cell proliferation, or lipopolysaccharide (LPS) (5 μg/mL) to induce B-cell proliferation. After 48 h incubation (37°C, 5% CO2 incubator), proliferation was determined by EZ-CyTox (Daeil Lab Service, Seoul, Korea) according to the manufacturer's instructions.
Determination of cytokine production in primary splenocytes
The splenocytes were cultured at a concentration of 1 × 106 cells/well in 96-well tissue culture dishes, and then was each treated with 200 μg/mL of CCW, CCE10%, and CCE70%, immediately followed by stimulation with 5 μg/mL Con A to induce Th1/Th2 type cytokines production. After 72 h incubation, the supernatants were collected and analyzed for IL-2, IFN-γ, IL-10, and IL-4 production. We measured the concentrations of cytokines using the R&D Duoset ELISA development kit (R&D Systems, Minneapolis, MN, USA).
Determination of IgE production in primary splenocytes
The splenocytes were cultured at a concentration of 1 × 106 cells/well in 96-well tissue culture dishes, and then each was treated with 200 μg/mL of CCW, CCE10%, and CCE70%, immediately followed by stimulation with 5 μg/mL LPS and 50 ng/mL IL-4 to induce IgE production, for 7 days. We collected the supernatants to assay IgE production. IgE concentration was measured using an IgE ELISA kit, from BD Biosciences Pharmingen (San Diego, CA, USA).
Determination of histamine production in MC-9 cells
MC/9 cells were cultured at a concentration of 1 × 104 cells/well in 96-well tissue culture dishes and incubated for 24 h. After 24 h incubation, the cells were each treated with 200 μg/mL of CCW, CCE10%, and CCE70%, immediately followed by stimulation with 50 nM phorbol 12-myristate 13-acetate (PMA) and 1 μM calcium ionophore A23187, to induce histamine release for 24 h. The supernatants were collected and analyzed for histamine levels. We measured the concentration of histamine using a mouse histamine ELISA kit (LDN, Nordhorn, Germany).
Statistical analysis
Data were expressed as mean ± standard deviation (SD), and were analyzed using Duncan's multiple range tests, after one-way analysis of variance (ANOVA), using SPSS statistical procedures for Windows (SPSS PASW Statistic 20.0; SPSS, Inc., Chicago, IL, USA). Statistical significance was considered at P < .05.
Results
mRNA expression of Eotaxin and TARC in HaCaT cells
The mRNA expressions of Eotaxin and TARC were significantly increased in the TNF-α and IFN-γ-stimulated HaCaT cells (control group), when compared with the nonstimulated normal control group cells. The treatments with CCW, CCE10%, and CCE70% in TNF-α and IFN-γ-stimulated HaCaT cells caused a marked increase in the mRNA expression of Eotaxin, compared with the TNF-α and IFN-γ-stimulated control group cells. However, the treatments with CCW and CCE70% caused significant decreases in mRNA expression of TARC, compared with the TNF-α and IFN-γ-stimulated control group cells (P < .05) (Fig. 1).
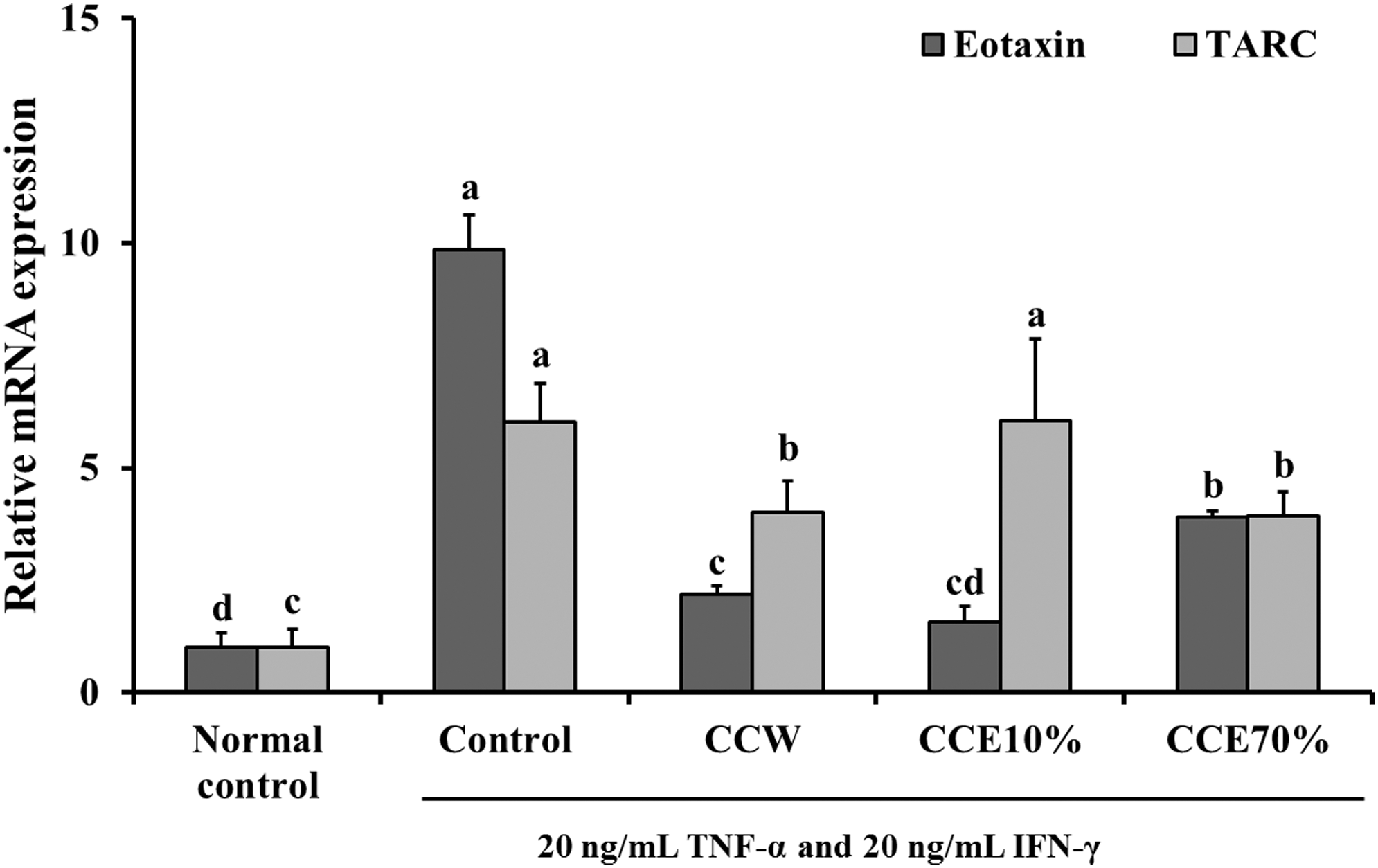
The effects of CC extracts on mRNA expression of chemokines (Eotaxin and TARC) in the TNF-α and IFN-γ-simulated HaCaT cells. Normal control, nonsimulated HaCaT cells; Control, TNF-α and IFN-γ-simulated HaCaT cells; CCW, treatment of 200 μg/mL CC water extract in the TNF-α and IFN-γ-simulated HaCaT cells; CCE10%, treatment of 200 μg/mL CC 10% ethanol extract in the TNF-α and IFN-γ-simulated HaCaT cells; CCE70%, treatment of 200 μg/mL CC 70% ethanol extract in the TNF-α and IFN-γ-simulated HaCaT cells. Values are expressed as the mean ± SD (n = 4). The different letters show a significant difference at P < .05, as determined by Duncan's multiple-range test. CC, Costaria costata; CCW, CC water; IFN, interferon; TARC, thymus, activation-regulated chemokine; TNF, tumor necrosis factor.
mRNA expression of TNF-α and IL-6 in HaCaT cells
The TNF-α and IFN-γ-stimulated HaCaT cells (control group) showed a marked increase in the mRNA expressions of TNF-α and IL-6, compared with the normal HaCaT cells (normal control group). When TNF-α and IFN-γ-stimulated HaCaT cells were treated with CCW, CCE10%, and CCE70%, the mRNA expressions of TNF-α production decreased significantly. The treatments with CCE10% and CCE70% caused significant decreases in mRNA expression of IL-6, compared with the TNF-α and IFN-γ-stimulated control group. There was not, however, a significant difference in the mRNA expression of IL-6 in the treatment with CCW, compared with the TNF-α and IFN-γ-stimulated control group (P < .05) (Fig. 2). These data in the TNF-α and IFN-γ-stimulated HaCaT cells suggest that CC can prevent the development of skin inflammation via suppression of production of chemokines and cytokines.

The effects of CC extracts on the mRNA expression of proinflammatory cytokines (TNF-α and IL-6) in the TNF-α and IFN-γ-simulated HaCaT cells. Normal control, nonsimulated HaCaT cells; Control, TNF-α and IFN-γ-simulated HaCaT cells; CCW, treatment of 200 μg/mL CC water extract in the TNF-α and IFN-γ-simulated HaCaT cells; CCE10%, treatment of 200 μg/mL CC 10% ethanol extract in the TNF-α and IFN-γ-simulated HaCaT cells; CCE70%, treatment of 200 μg/mL CC 70% ethanol extract in the TNF-α and IFN-γ-simulated HaCaT cells. Values are expressed as the mean ± SD (n = 4). The different letters show a significant difference at P < .05, as determined by Duncan's multiple-range test. IL, interleukin.
T and B-cell proliferation in primary splenocytes
ConA-stimulated primary splenocytes from the AD-induced mice (control group) showed significantly decreased T-cell proliferation, when compared with those of the normal control group (Fig. 3A). On the other hand, B-cell proliferation of the LPS-stimulated primary splenocytes from the AD-induced mice (control group) increased significantly, compared with those of the normal mice control group (Fig. 3B). Treatments of CCW, CCE10%, and CCE70% caused significant changes in the T cell and B-cell proliferation levels, compared with control group levels (P < .05) (Fig. 3).
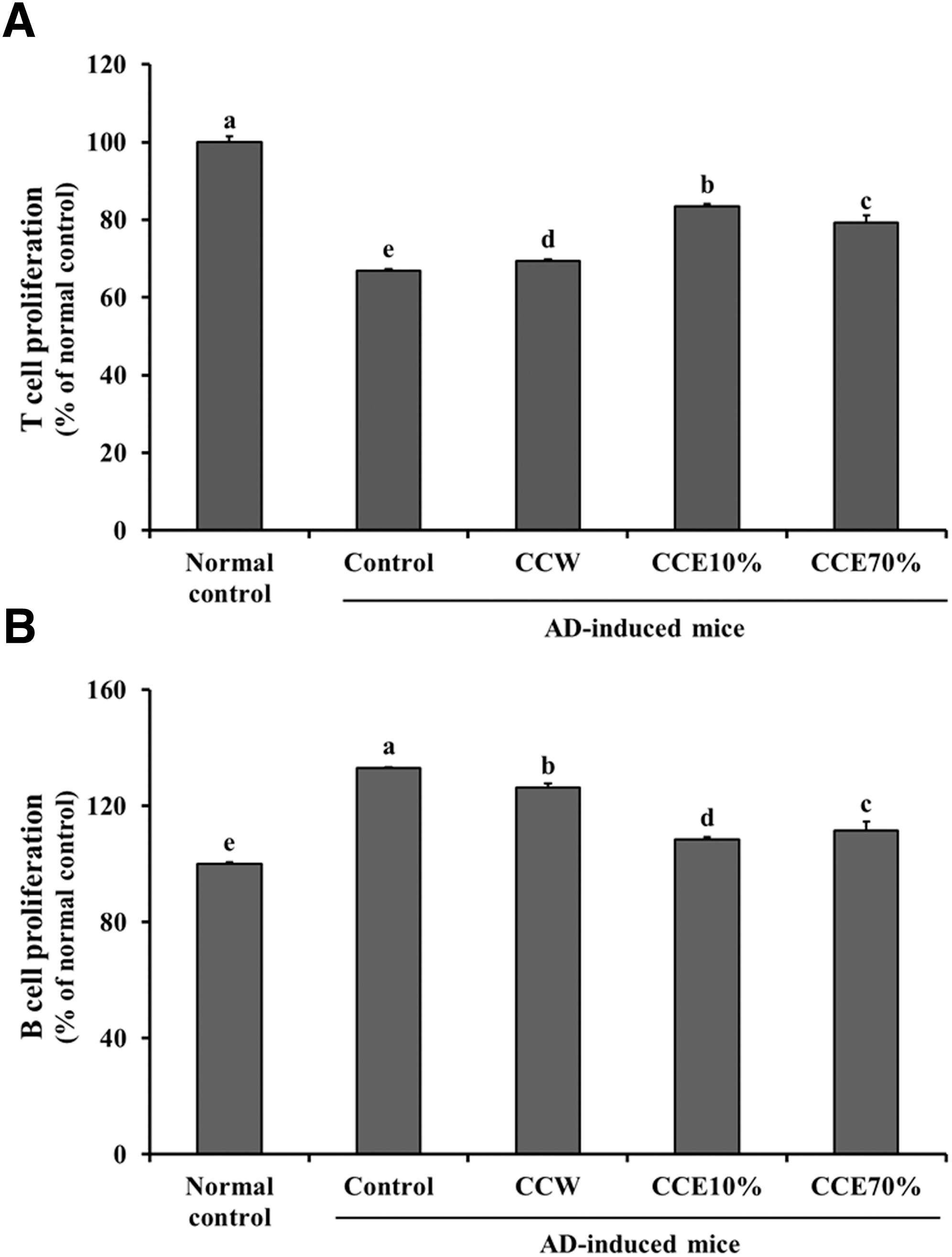
The effects of CC extracts on
Th1 and Th2 type cytokine production in primary splenocytes
The production of Th1 type cytokines (IL-2 and IFN-γ) in ConA-stimulated primary splenocytes (control group) from the AD-induced mice were significantly decreased, compared with that seen in the normal mice control group. The ConA-stimulated primary splenocytes treated with CCW, CCE10%, and CCE70% showed significant increases in the production of IFN-γ, compared with that of the control group. IL-2 production results, however, showed a significant increase compared with the control group in the CCE70% group, but not in the CCW or CCE10% groups (P < .05) (Fig. 4). ConA-stimulated primary splenocytes (control group) from AD-induced mice resulted in a significant increase in the production of Th2 type cytokines (IL-10 and IL-4), compared with that in the normal control group. Compared with the control group, however, the productions of IL-10 and IL-4 were significantly decreased in the CCW, CCE10%, and CCE70% treatment groups (P < .05) (Fig. 5). In summary, CC extracts suppressed the imbalance of Th1/Th2 cytokines and T/B cell proliferation, suggesting that CC may have benefits for improvement of AD or other allergy.
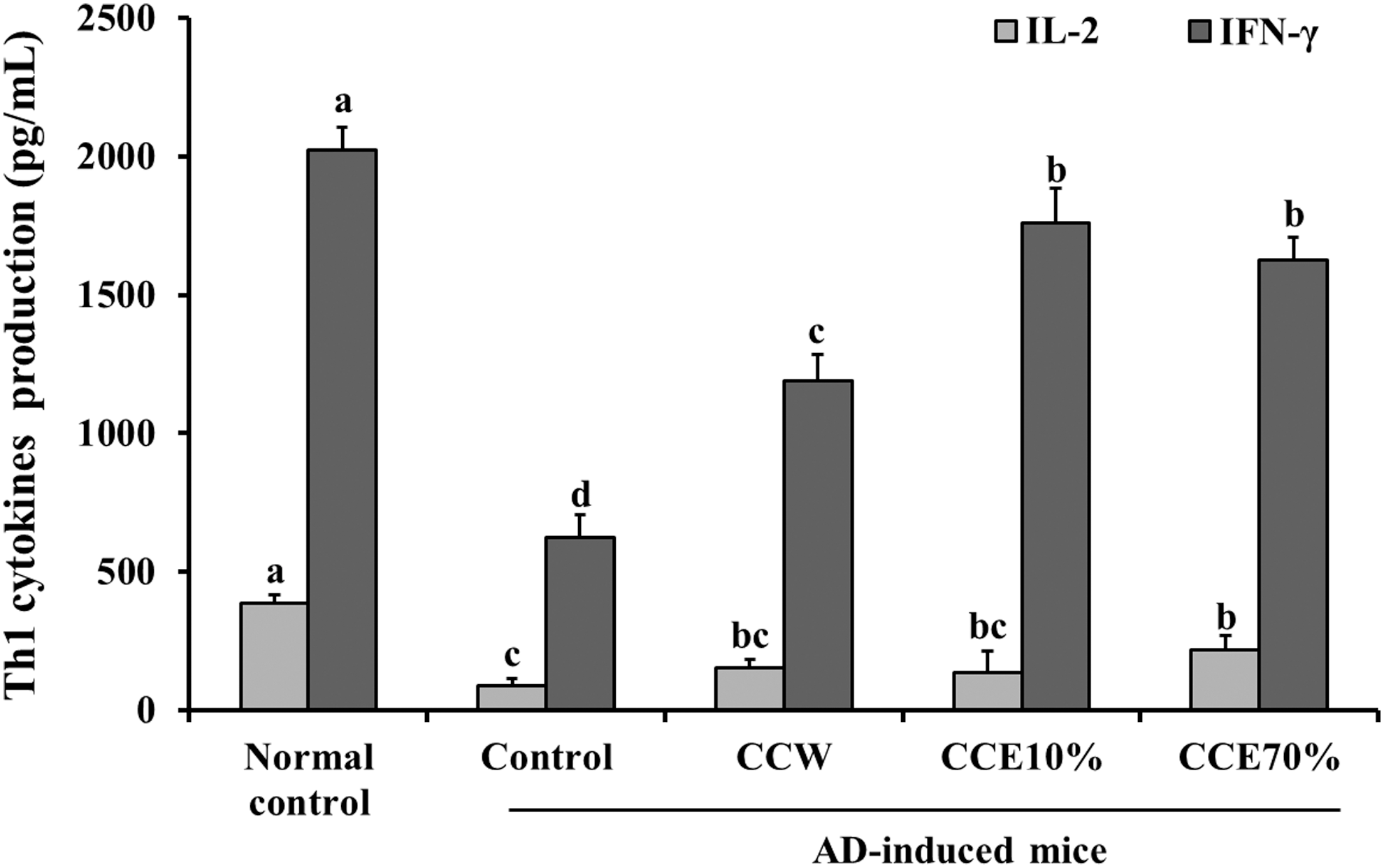
The effects of CC extracts on Th1 cytokines (IL-2 and IFN-γ) in ConA-stimulated splenocytes from normal mice and AD-induced mice. Normal control, splenocytes from normal Balb/c mice; Control, splenocytes from AD-induced mice; CCW, treatment of 200 μg/mL CC water extract in splenocytes from AD-induced mice; CCE10%, treatment of 200 μg/mL CC 10% ethanol extract in splenocytes from AD-induced mice; CCE70%, treatment of 200 μg/mL CC 70% ethanol extract in splenocytes from AD-induced mice. Values are expressed as the mean ± SD (n = 4). The different letters show a significant difference at P < .05, as determined by Duncan's multiple-range test.
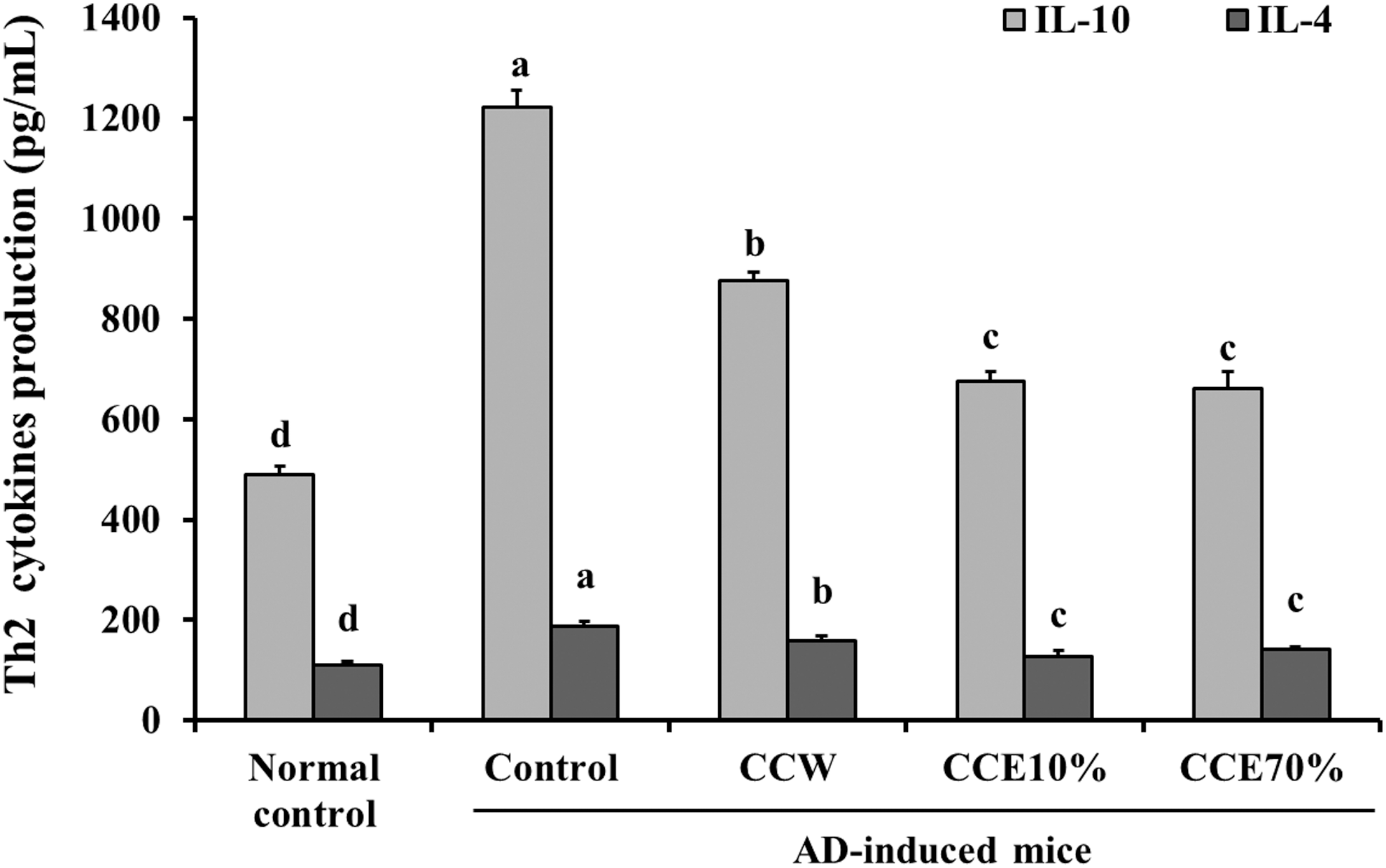
The effects of CC extracts on Th2 cytokines (IL-10 and IL-4) in ConA-stimulated splenocytes from normal mice and AD-induced mice. Normal control, splenocytes from normal Balb/c mice; Control, splenocytes from AD-induced mice; CCW, treatment of 200 μg/mL CC water extract in the splenocytes from AD-induced mice; CCE10%, treatment of 200 μg/mL CC 10% ethanol extract in the splenocytes from AD-induced mice; CCE70%, treatment of 200 μg/mL CC 70% ethanol extract in the splenocytes from AD-induced mice. Values are expressed as the mean ± SD (n = 4). The different letters show a significant difference at P < .05, as determined by Duncan's multiple-range test.
IgE production in primary splenocytes
The LPS and IL-4-stimulated splenocytes (control group) from the AD-induced mice showed a marked increase in the expression of IgE production, compared with the normal control group. In contrast, IgE production decreased significantly in the LPS and IL-4-stimulated splenocytes treated with CCW, CCE10%, and CCE70%, compared with that in the control group (P < .05) (Fig. 6). According to this data, we can suggest that CC may prevent IgE isotype switching in B cell.

The effects of CC extracts on IgE production in LPS and IL-4-stimulated splenocytes from normal mice and AD-induced mice. Normal control, splenocytes from normal Balb/c mice; Control, splenocytes from AD-induced mice; CCW, treatment of 200 μg/mL CC water extract in the splenocytes from AD-induced mice; CCE10%, treatment of 200 μg/mL CC 10% ethanol extract in the splenocytes from AD-induced mice; CCE70%, treatment of 200 μg/mL CC 70% ethanol extract in the splenocytes from AD-induced mice. Values are expressed as the mean ± SD (n = 4). The different letters show a significant difference at P < .05, as determined by Duncan's multiple-range test. Ig, immunoglobulin E.
Histamine production in MC/9
The production of histamine in PMA and A23187-stimulated MC/9 cells (control group) was increased significantly, when compared with that in the nonstimulated MC/9 cells (normal control group). Treatment with CCW, CCE10%, and CCE70%, however, caused a significant decrease in the production of histamine, when compared with the control group cells (P < .05) (Fig. 7). The results suggest that CC may attenuate the activation of mast cells and production of histamine.
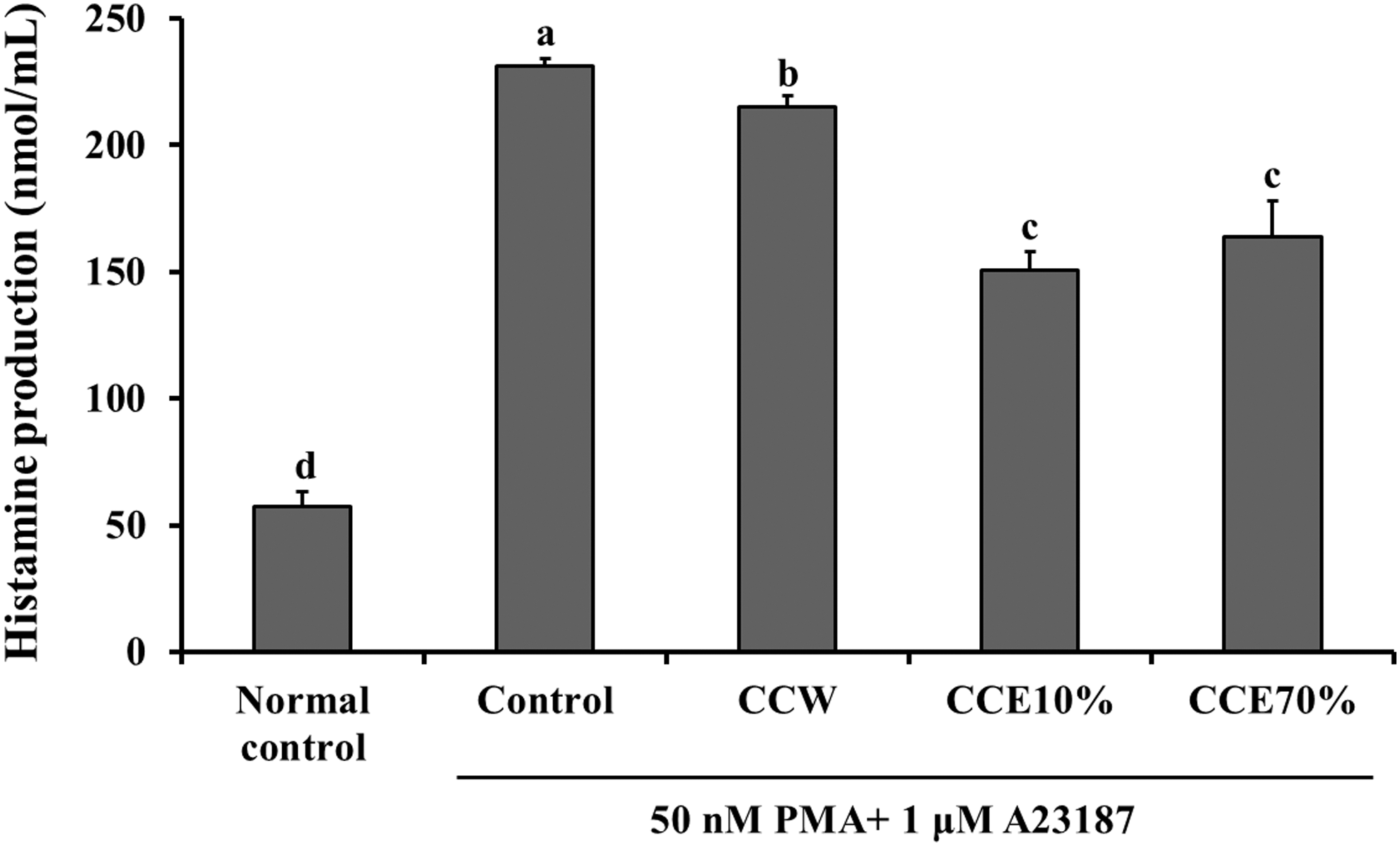
The effects of CC extracts on histamine production in the PMA and A23187-simulated MC/9 cells. Normal control, non-simulated MC/9 cells; Control, PMA and A23187-simulated MC/9 cells; CCW, treatment of 200 μg/mL CC water extract in the PMA and A23187-simulated MC/9 cells; CCE10%, treatment of 200 μg/mL CC 10% ethanol extract in the PMA and A23187-simulated MC/9 cells; CCE70%, treatment of 200 μg/mL CC 70% ethanol extract in the PMA and A23187-simulated MC/9 cells. Values are expressed as the mean ± SD (n = 4). The different letters show a significant difference at P < .05, as determined by Duncan's multiple-range test.
Discussion
The keratinocytes on the outer surface of the epidermis work as a protective layer against environmental damage using its immunological reaction, such as expression and secretion of immune mediators in response to inflammation. 5 Several studies have shown that TNF-α and IFN-γ induce abnormal expression of proinflammatory cytokines and specific chemokines in keratinocytes, which can play a role in the infiltration of leukocytes into the skin during the development of AD. 20,21 As such, decreasing the production of proinflammatory cytokines in keratinocytes may be an effective target for AD therapy.
In this study, we investigated the effects of CC on TNF-α and IFN-γ-induced activation of inflammatory response in HaCaT cells, which are a substitute for human primary keratinocytes. Vestergaard et al. 22 found that the level of TARC production was significantly higher when stimulated with both TNF-α and IFN-γ, compared with when stimulated with either TNF-α or IFN-γ alone. In addition, Kong et al. 23 reported that stimulation with TNF-α and IFN-γ caused abnormal expression of TNF-αR1 and IFN-γR1, and abnormal production of proinflammatory cytokines, including IL-8 and IL-1β in HaCaT cells. We also found that treatment with TNF-α and IFN-γ induced an increase in mRNA expression of Eotaxin, TARC, and proinflammatory cytokines, including TNF-α and IL-6 in HaCaT cells (Figs. 1 and 2). Treatment with CC extracts in TNF-α and IFN-γ-stimulated HaCaT cells, however, only partially induced a marked increase in the mRNA expression of chemokines and proinflammatory cytokines, compared with the TNF-α and IFN-γ-stimulated control group. In particular, CCE70% caused a significant decrease in the expression of all the chemokines and proinflammatory cytokines tested (Figs. 1 and 2).
In the early stages of AD, Th cells are activated and produce increasing Th2 type cytokines, such as IL-4, IL-5, IL-10, and IL-13 from Th2 cells, while decreasing Th1 type cytokines, such as IFN-γ and IL-2, from Th1 cells. 7 Th2 type cytokines can activate class switching to IgE in B cells, which play a role in AD pathology by mast cell activation. 24 Punnonen et al. 25 investigated the role of IL-4 in IgE synthesis induced by cytokines derived from activated Th cells, and they found that IL-4 contributes to IgE synthesis induced by all Th cell subsets. As such, this Th2-associated pathology induces abnormal immunological reactions, which subsequently lead to epidermal barrier defects during the development of AD. In addition, there is evidence that Th1 type cytokines, especially IFN-γ, play a key role in chronic AD by switch to a Th1 cells response. 26 Endo et al. 24 discussed that balance of Th1/Th2 cells and cytokines is associated with the pathogenesis of autoimmune diseases or allergic diseases.
We found that Th2 type cytokines increased, and Th1 type cytokines decreased, in ConA-stimulated primary splenocytes from the AD-induced mice, although T-cell proliferation also decreased. The results suggest that the experimental design of this study does not appear to correlate with chronic AD. We treated the primary splenocytes from the AD-induced mice with LPS and IL-4 to stimulate IgE production, and found a marked increase in the expression of IgE production in the stimulated group, compared with the nonstimulated group. In contrast, treatment with CC extracts resulted in a significant increase in T-cell proliferation and production of IFN-γ, compared with the control group (Figs. 3 and 4). In addition, the production of Th2 type cytokines and IgE were significantly decreased in the groups treated with CC extracts (Figs. 5 and 6).
Many previous studies have suggested that serum IgE levels in AD patients correlate with the degree of AD, and, as such, high serum IgE level is a strong factor in AD diagnosis. 9 One of the functions of IgE in AD pathogenesis is the sensitization of mast cells, through the high affinity FcɛRI at the cell surface, which mediates the inflammation responses. 8,27 Once mast cells are activated by IgE binding, diacylglycerol (DAG) and inositol 1,4,5-trisphosphate (IP3) activate protein kinase C (PKC), and elevate intracellular calcium levels, which promote the movement of granule including histamines, and fusion with the plasma membrane. 28,29 Histamine is well known for its role in itching, which is the most common symptom associated with AD. 30 In addition, recent reports found that the histamine 4 receptor is highly expressed on keratinocytes in the pathogenesis of AD, which can induce the epidermal hyperplasia by keratinocyte proliferation. 30 For this reason, antihistamines are often used to treat itching in AD patients, but it has side effects, including headaches and abdominal pain. 31 In this study, we used PMA to activate PKC and A23187 to increase intracellular Ca2+ levels for mast cell activation and stimulate histamine release. We found that PMA and A23187 induced an increase in the release of histamine. Groups treated with CC extracts, however, showed a significant decrease in the release of histamine, compared with control groups.
In summary, we investigated the effects of CC extracts on AD in an in vitro model of AD, for which we provided a protocol. We found that CC extracts resulted in a decrease in the production of proinflammatory cytokines in keratinocytes, and the suppression of the imbalance of Th1/Th2 cytokines and IgE production of primary splenocytes. In addition, CC extracts decrease the release of histamine from PMA and A23187-simulated MC/9 cells. These results suggest that CC extracts may be effective for treating other allergy diseases, and AD, through the attenuation of the allergic reaction response.
Footnotes
Acknowledgment
This research was supported by a grant from the Marine Biotechnology Program (PJT200672), funded by the Ministry of Oceans and Fisheries, Korea.
Author Disclosure Statement
No competing financial interests exist.