
Abstract
The edible red seaweed Gracilariopsis chorda (Holmes) Ohmi is known for its extensive medicinal benefits and its use as a food ingredient in Korea, Japan, and China. In a previous study, an ethanol extract of G. chorda (GCE) showed potential neuroprotective effects in cultured hippocampal neurons. In this study, we further examined the ability of GCE to promote neurite extension in primary rat hippocampal neurons. Neurons were stained with the lipophilic dye DiO or immunostained to visualize the neuronal morphology. The results indicated that GCE concentration-dependently increased neurite outgrowth, with an optimal concentration of 30 μg/mL. GCE significantly promoted early neuronal differentiation (i.e., polarity and process number) and enhanced axonal and dendritic arborization in a time-responsive manner. In addition, arachidonic acid, which was previously identified and quantified as a major neuroprotective component of GCE, significantly accelerated neurite outgrowth similar to GCE. Our findings suggest that G. chorda and its active component, arachidonic acid, may be useful for developing medicinal food or pharmaceuticals in the prevention and treatment of neurological disorders.
Introduction
A
Extracellular neuritogenic substances of plant origin have attracted attention not only because of their effects in neuronal development, differentiation, and survival but also because of their efficacy in maintaining functional activity in the degenerating brain. Many researchers have reported neurotrophic medicinal plants and assessed their extracts for drug development.
3
Among the seaweeds examined, Porphyra yezoensis, Sargassum fulvellum, Gelidium amansii, and Undaria pinnatifida have multiple neurotrophic and neuroprotective activities that are also effective in the early development of primary hippocampal neurons and can promote axodendritic arborization,
4
–7
synaptogensis, and N-methyl-
Embryonic hippocampal neurons develop morphologically and functionally in a primary culture system; this system is advantageous because it includes several structural phenotypes and is appropriate for morphometric analyses. Previously, we reported that an ethanol extract of Gracilariopsis chorda (GCE) protected hippocampal neurons from oxidative stress induced by hypoxia/reoxygenation (H/R). 13 GCE had a neuromodulatory effect in hippocampal neurons. Therefore, we sought to identify the neuroactive substance(s) responsible for inducing neurotrophic factor-like activity in the seaweed extract. In this study, we evaluated the effects of GCE and its active component, arachidonic acid (AA), on axodendritic morphology in a rat primary hippocampal culture model.
Materials and Methods
Collection and processing of G. chorda
The red seaweed G. chorda (Holmes) Ohmi was collected along the south-eastern coastal region of the Korean peninsula. The sample was washed with fresh water to remove epiphytes and salts. After 1 week, the dried tissues were pulverized using a grinder (HMF-340; Hanil Co., Seoul, Korea) and stored in sealed plastic bags under dark conditions at −20°C until use. A voucher specimen (SW-23) was deposited in the laboratory of Dr. Y.K. Hong (Pukyong National University, Busan, Korea). The seaweed powder was extracted with 95% ethanol at a ratio of 50:1 (v/w). The mixture was kept on an orbital shaker (VS-202D; Vision Scientific Co. Ltd., Seoul, Korea) at 200 rpm at room temperature for 24 h under dark conditions. Then, the slurry was centrifuged at 10,000 g (Sorvall T6000D centrifuge; Thermo Fisher Scientific, Inc., Waltham, MA), and the supernatant was filtered through sterile cotton wool. The filtrate was concentrated in vacuo and dried completely under a stream of nitrogen gas. The extract yield was 0.22% (w/w). The extract was dissolved in dimethyl sulfoxide (DMSO) to make aliquots (8 mg/mL), which were stored in foil-wrapped vials at −20°C for future experiments. In addition, AA was identified as a major active component and quantified by reverse-phase high-performance liquid chromatography (RP-HPLC) according to Mohibbullah et al. 13
Primary culture of hippocampal neurons and extract treatment
This study was approved by the Institutional Animal Care and Use Committee of the School of Medicine, Dongguk University. All reagents used for the primary cell cultures were purchased from Invitrogen (Carlsbad, CA) unless otherwise stated. Pregnant Sprague-Dawley rats were received on the 13th day of pregnancy and housed in a controlled environment with access to food and water ad libitum. On the 19th day of pregnancy, rats were euthanized with isoflurane, and the fetuses were collected. Fetal hippocampi were excised from the brains, and hippocampal neuronal cultures were prepared as described previously. 14,15 The dissociated cells were plated at a density of 1.0 × 104 cells/cm2 for the morphometric study. The seaweed extracts, AA, and vehicle (DMSO, final concentration <1%) were added to the media before cell plating.
DiO staining
On day in vitro (DIV) 3, neurons were live stained with Vybrant™ DiO (Invitrogen) according to the manufacturer's protocol. The lipophilic nature of DiO, which binds uniformly to the plasma membrane of cultured cells, enables the visualization of entire neurons using epifluorescence microscopy.
Immunocytochemistry
Neurons on cover slips were rinsed briefly with Dulbecco's phosphate-buffered saline (D-PBS; Invitrogen) and fixed using a sequential paraformaldehyde/methanol fixation procedure 16 on DIV 3 and 5. For immunostaining, the following antibodies were used: primary antibodies against microtubule-associated protein 2 (mouse monoclonal MAP2, 1:500 dilution; Sigma-Aldrich Co., St. Louis, MO), tubulin α-subunit (mouse monoclonal 12G10, 1:1000 dilution; Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA), ankyrin G (rabbit polyclonal H-215, 1:50 dilution; Santa Cruz Biotechnology, Inc., Dallas, TX), and secondary antibodies (Alexa Fluor® 568-conjugated goat anti-mouse IgG, 1:1000, Alexa Fluor® 488-conjugated goat anti-mouse IgG, 1:1000, and Alexa Fluor® 568-conjugated donkey anti-rabbit IgG, 1:1000; Molecular Probes, Eugene, OR). Fixed neurons were incubated with a primary antibody, followed by a secondary antibody, and mounted on slides as described previously. 16
Image acquisition
A Leica DM IRE2 research microscope equipped with I3 S, N2.1S, and Y5 filter systems (Leica Microsystems AG, Wetzlar, Germany) was used for phase-contrast and epifluorescence microscopy. Images (1388 × 1039 pixels) were acquired using a high-resolution CoolSNAP™ CCD camera (Photometrics, Inc., Tucson, AZ) under the control of a computer running Leica FW4000 software (Leica Microsystems AG).
Image analysis and quantification
Morphometric analyses and quantification were performed using ImageJ (ver. 1.45) software with the neurite tracer (National Institutes of Health [NIH], Bethesda, MD) and Sholl plug-ins (
Data analysis
All data are expressed as mean ± standard error of the mean of at least three independent experiments. Statistical comparisons were made using Student's t-tests, χ2 tests, and a one-way analysis of variance with Duncan's post hoc multiple comparisons (SPSS Software, ver. 16.0; SPSS, Inc., Chicago, IL). P-values <.05 were considered to indicate statistical significance.
Results
GCE enhanced neurite tree structure
Initially, in the morphometric analysis, we confirmed that the neuronal cultures had been grown with no visible contamination by other brain cell types (e.g., glial cells) and were characterized by extensive processes and glossy cell bodies associated with strong light refraction. Hippocampal neurons stained positive for MAP2, a neuron-specific marker, at DIV 3 (Fig. 1A). To optimize the GCE concentration for neurite outgrowth-promoting activity, hippocampal neurons were incubated with GCE at different concentrations, from 7.5 to 60 μg/mL. Typical DiO-stained images were obtained at DIV 3 (Fig. 1B). Four different growth parameters (the number and total length of primary neurites, the length of the longest neurites, and the number of branching points) increased significantly and concentration-dependently when neurons were treated with GCE (Fig. 1C). The optimal concentration of GCE was 30 μg/mL. GCE was used at this concentration for further experiments.
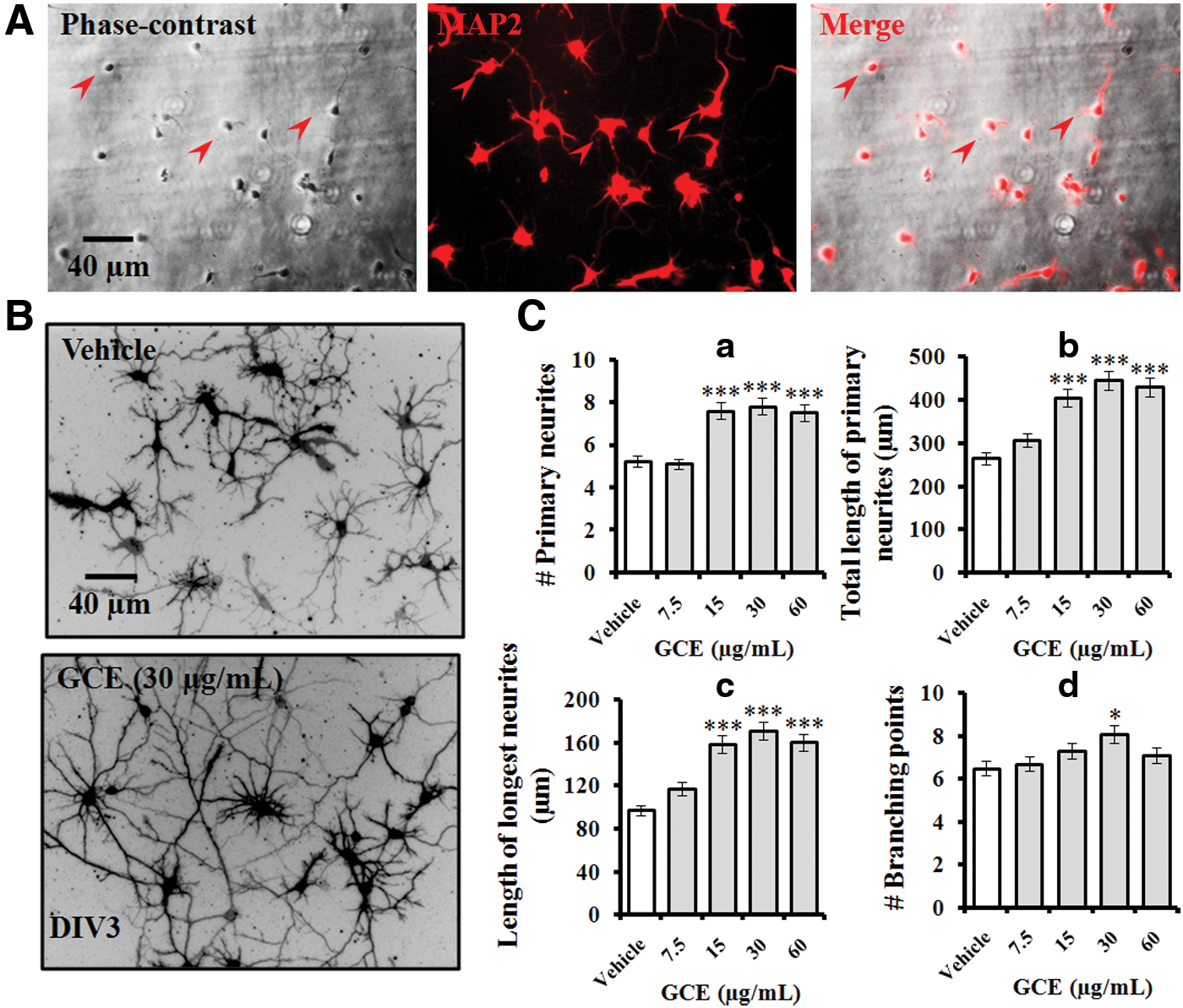
GCE enhanced neurite structure in a concentration-dependent manner. Embryonic rat hippocampal neurons were cultured on poly-
GCE promoted early neuronal differentiation
We investigated the effects of GCE on early neuronal differentiation in DIV 1 cultures. Neurons in different developmental stages, such as stage 1 (lamellipodia), 1.5 (fan-shaped), and 2 (minor process), were counted (Fig. 2A-a). A significant difference in the number of neurons in each developmental stage was observed between cells treated with GCE and those without. Significantly increased numbers of stage 2 cells (26%) were observed in GCE-treated cells compared with control cells (χ2 test: P < .001; Fig. 2B-a). In the same cultures, we counted neurons based on the number of processes and categorized them as unipolar, bipolar, or multipolar (Fig. 2A-b). About 27% more cells in the GCE-treated culture developed multipolar characteristics after 24 h of culture versus the controls (χ2 test: P < .01; Fig. 2B-b).
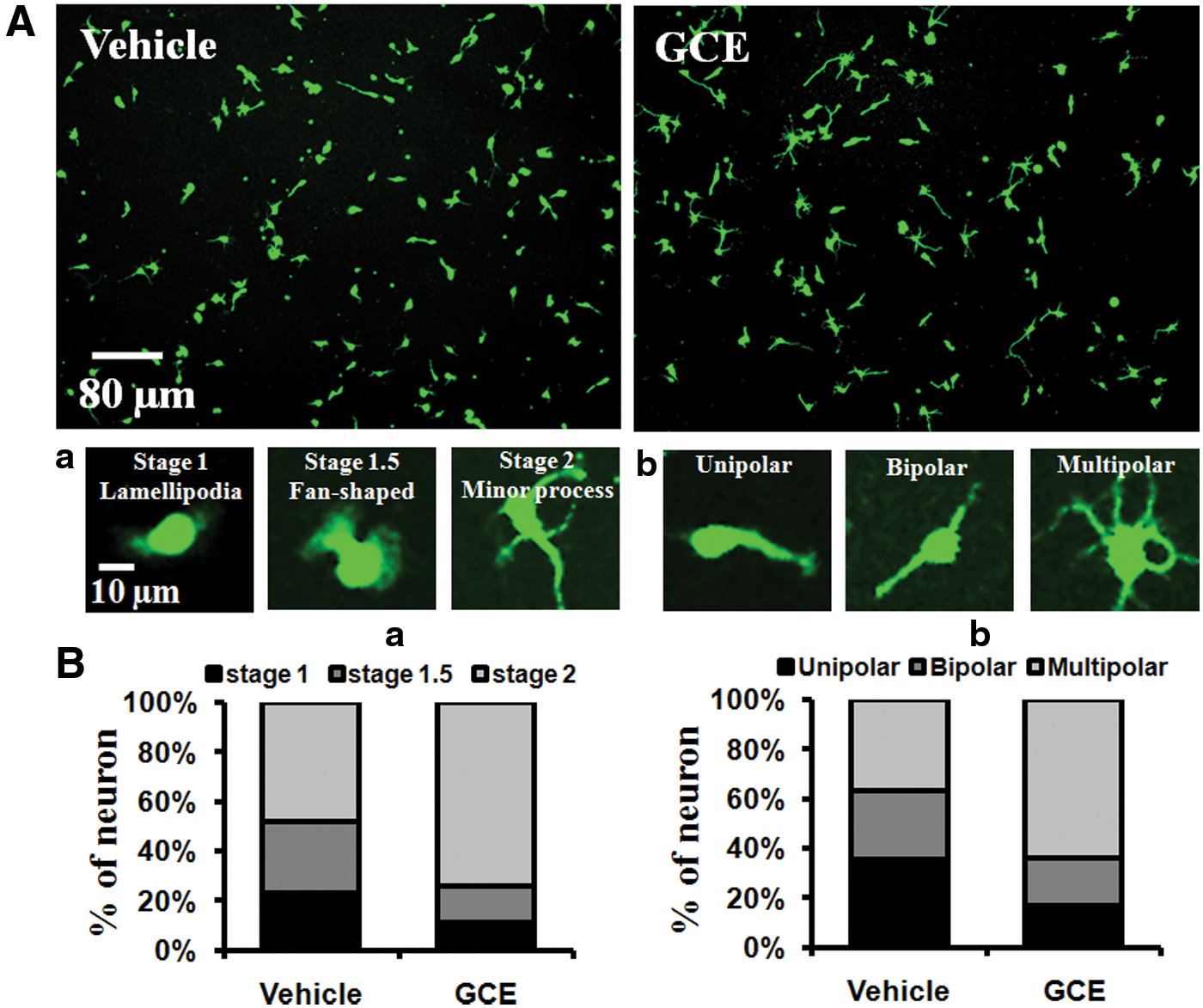
GCE promoted early neuronal maturation. Hippocampal neurons were grown in the same culture conditions described in Figure 1 with vehicle or GCE extract (30 μg/mL) for 24 h, fixed, and immunostained for MAP2.
GCE stimulated dendritic structural dynamics
Dendritic elongation and branching of cultured neurons were defined as developmental stage IV (dendritic outgrowth), which began on DIV 3 to 5 in typical cultures. As a part of the subsequent morphogenesis of neurons, dendritic structure and branching projections are required for proper externalization of the neuronal circuitry. To recognize axons from dendrites in culture, neurons were double immunostained for ankyrin G (red) and α-tubulin (green). Because ankyrin G is characteristically located at the axon initial segment, this segment was a reddish color in merged images. Neuronal cells were observed from DIV 3 and DIV 5 (Fig. 3A). Treatment with GCE significantly increased the number of primary dendrites (by 60% and 45% vs. the control at DIV 3 and DIV 5, respectively; Fig. 3B-a) and TLPD (by 64% and 100% vs. the control at DIV 3 and DIV 5, respectively; Fig. 3B-b) in a time-course study. The number (Fig. 3B-c) and total length of the dendritic branches (Fig. 3B-d) were also increased significantly by GCE treatment in cultures.
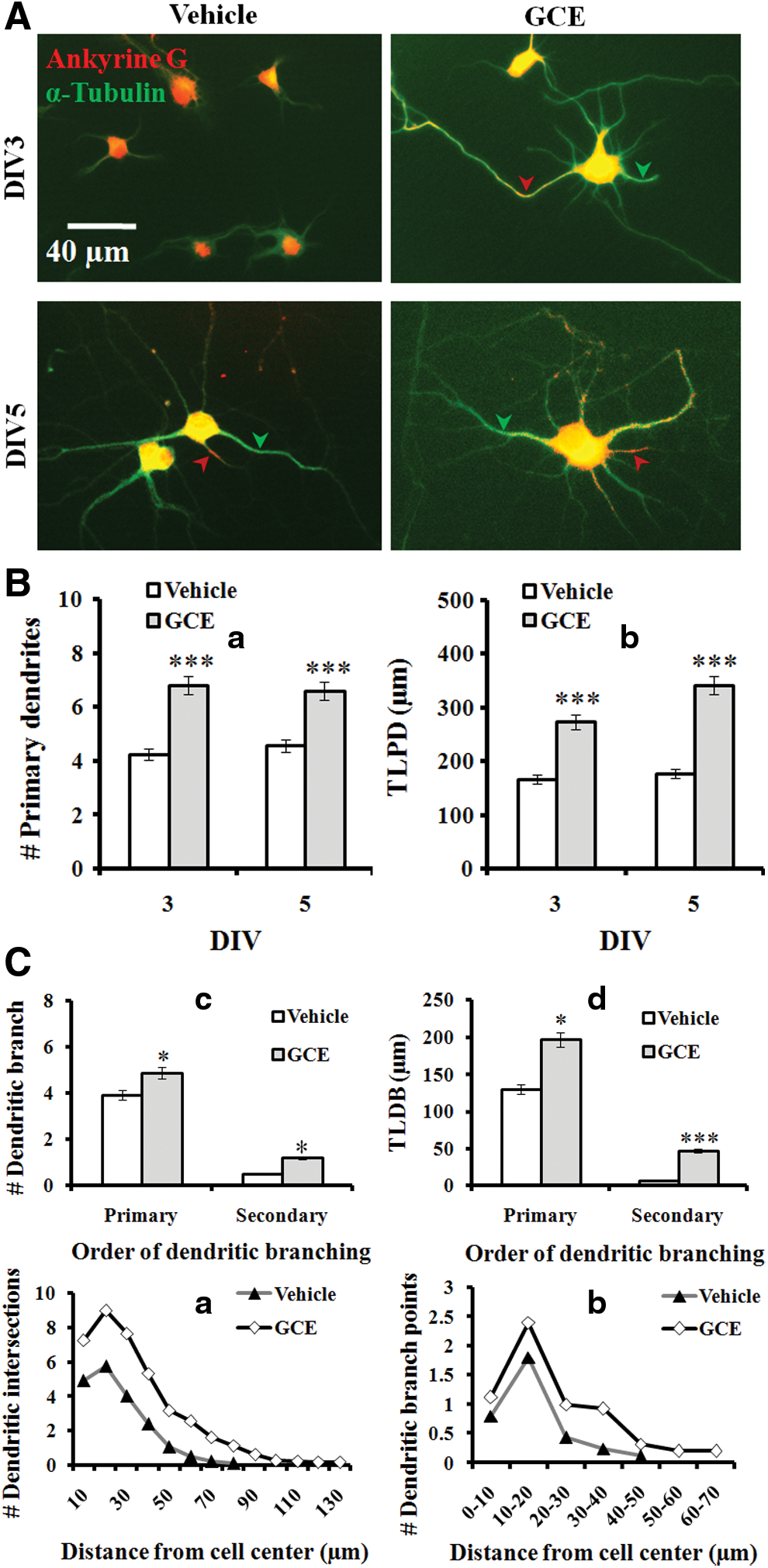
GCE stimulated dendritic structural dynamics. Neurons were grown in the same culture conditions indicated in Figure 1 for 3–5 days. Neurons were fixed and double immunostained for ankyrin G (red) and α-tubulin (green). Ankyrin G is characteristically located at the axon initial segment (red arrow in
For the quantitative assessment of dendritic projections, we used Sholl's analysis, which measures the dendritic intersections at different radical distances of Sholl's circle from the cell body. In addition to the intersections, we also counted branching points within two succeeding concentric Sholl circles. In GCE-treated neurons, the number of dendritic intersections and dendritic branching points were increased by 105% and 81%, respectively, compared with the controls. In addition, dendritic intersections and branching points were found up to the 130 and 60 μm circles, respectively, when cultures were treated with GCE. In contrast, no intersections or branching points were found in control neurons beyond the 80 and 40 μm circles, respectively (Fig. 3C-a, b). These findings indicate that GCE has a promising effect for promoting dendritic complexity in developing neurons.
GCE induced axonal sprouting
Axonal sprouting is an essential step in neuronal morphogenesis, which allows connecting adjacent neurons to form a robust neuronal network. Moreover, axonal sprouting may be valuable in the establishment and refinement of early neuronal cells during development and in damaged neurons during the onset of neurodegenerative events. Therefore, we examined the effect of GCE on axonal maturation in DIV 3 and DIV 5 cultures (Fig. 3A). Upon treatment with GCE in culture, there was a significant increase in axonal length in a time-dependent manner (by 76% and 47% vs. the control at DIV 3 and DIV 5, respectively; Fig. 4A-a). Moreover, GCE treatment significantly increased the number of axonal collateral branches (ACB; Fig. 4A-b) and the total length of axonal collateral branches (TLACB; Fig. 4A-c) for both primary and secondary orders, respectively, compared with the controls. We did not find any tertiary branches in control cultures, whereas GCE-treated neurons possessed branching orders up to the tertiary level.
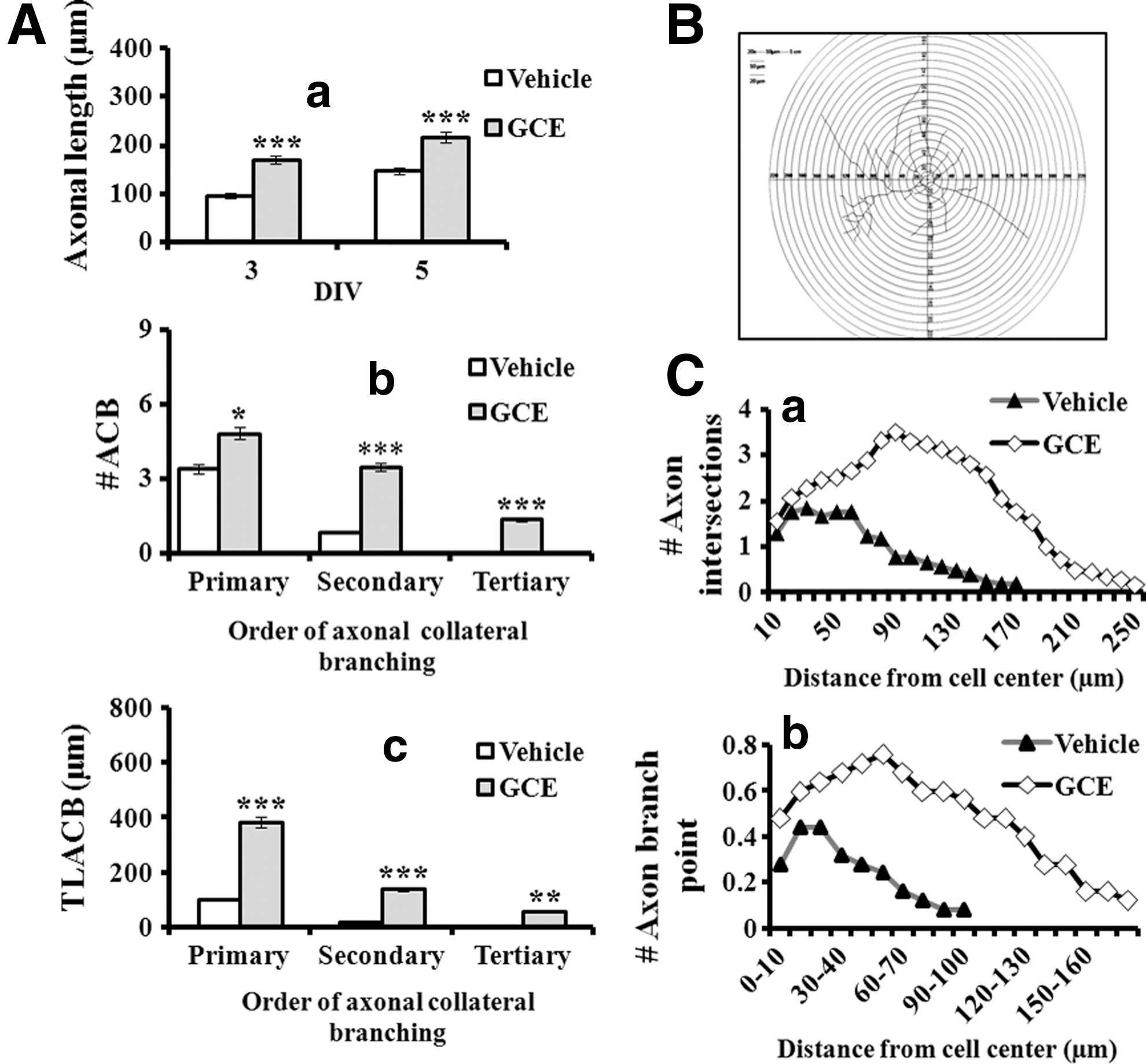
GCE induced axonal sprouting. This figure is based on the same representative photographs shown in Figure 3A.
In the Sholl analysis (Fig. 4B), the numbers of axonal intersections and branch points of GCE-treated neurons increased by 198% and 256%, respectively, compared with control neurons. In the Sholl circles, the axonal intersections and branch points extended up to 250 and 170 μm, respectively. However, in control cells, no axonal intersection or branch point was visible beyond the circles of 170 and 90 μm, respectively (Fig. 4C-a, b). Overall, GCE significantly enhanced axonal arborization.
AA potentiated neuronal outgrowth
Previously, we identified and quantified AA as a major active component having neuroprotective efficacy against H/R-induced cell death. In this study, we further examined the effects of AA and whether it could promote neuronal arbor structures in rat embryonic hippocampal neurons. At DIV 3, neurons were fixed and immunostained for α-tubulin (green) to investigate the effects of GCE and/or AA on morphogenesis (Fig. 5A). The results indicated that AA at an optimal concentration of 1 μM significantly increased the number of primary neurites (by 49% vs. the control; Fig. 5B-a), the total length of primary neurites (by 69% vs. the control; Fig. 5B-b), the length of the longest neurite (by 80% vs. the control; Fig. 5B-c), and the number of branching points (by 83% vs. the control; Fig. 5B-d). These effects of AA were comparable with the effects of GCE.
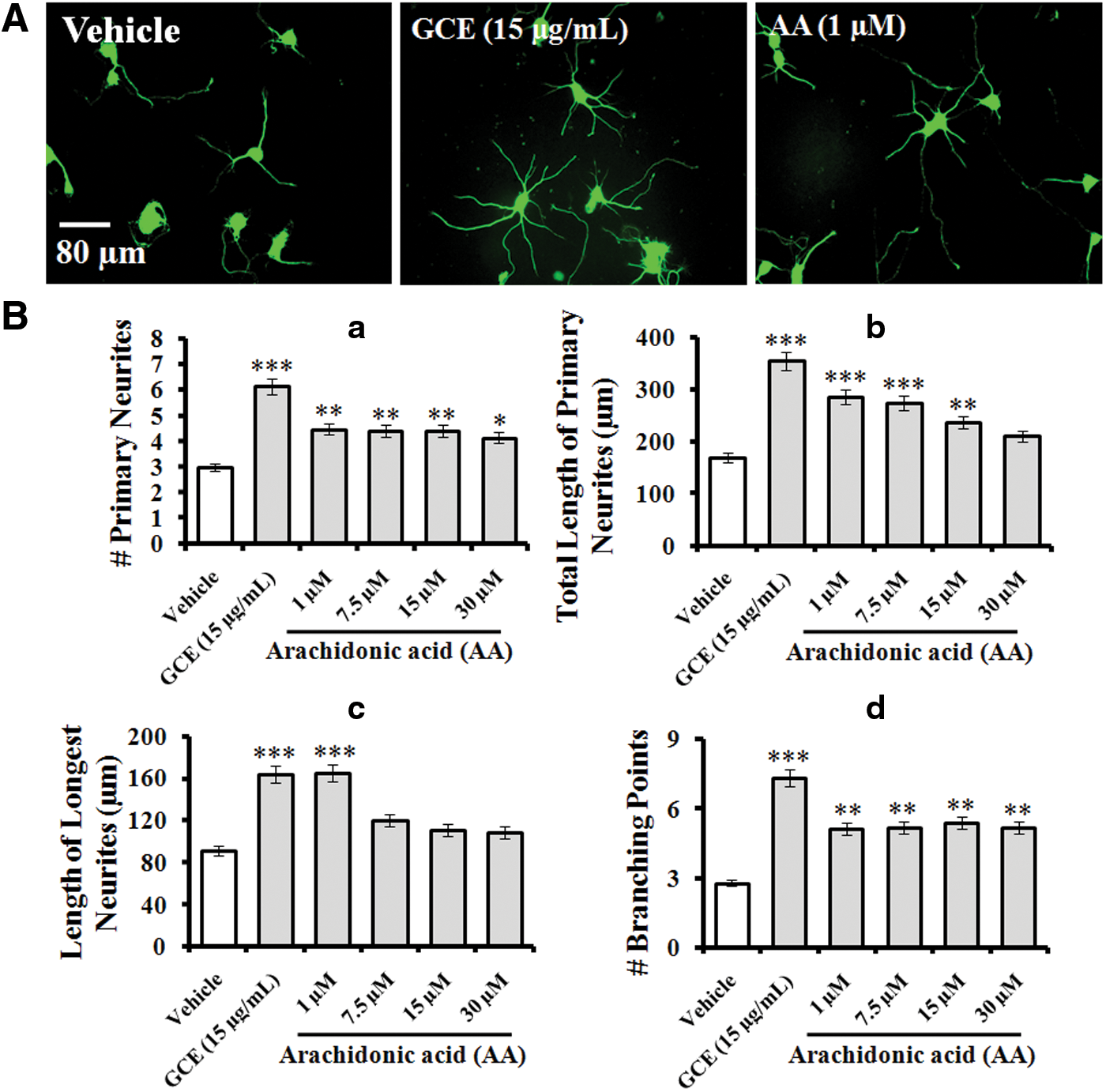
Effect of AA on neurite outgrowth activity. Neurons were grown in the same culture conditions indicated in Figure 1 in the presence of GCE (30 μg/mL) or AA at the indicated concentrations for 3 days. Neurons were fixed and immunostained for α-tubulin (green).
Discussion
An ethanolic extract from the seaweed G. chorda was examined for its ability to potentiate neurite outgrowth in primary hippocampal neurons in a serum-free medium. GCE provided potent trophic support to these neurons because the axon and dendrites were robust in form. Our results are consistent with a recent report showing that GCE and its identified active compound, AA, protected hippocampal neurons in H/R-induced neuronal injury. 13 Treatment of neurons with the polar lipid AA can enhance the outgrowth activity by stimulation of a plasma membrane protein, syntaxin 3 (STX3), in rapidly growing cells, 19 and by the activation of phospholipase A2, which facilitates the cognitive activity at an early stage of Alzheimer's disease. 20 During neuronal development in vitro, stereotypical morphological differentiation stages that lead to fully polarized neurons in culture are essential. The early differentiation of neuronal processes that finally become axons and dendrites can establish structural and functional plasticity in developing neurons. 21,22 In this study, GCE supported early developmental events and promoted the maturation of growing neurons, suggesting that GCE may contain several functional molecules that are important for generating robust neuronal networks. In future studies, the bioassay-guided isolation of other active compounds and their modes of action should be investigated.
Brain aging is a critical risk factor in most neurodegenerative disorders characterized by memory impairment and disorientation, particularly in Alzheimer's disease, as a result of loss of neuronal structure and function. Selective elimination of axons, dendrites, and their branches, even without the death of the parent neurons, are common scenarios in acute and chronic neurodegeneration. 23 Evidence suggests that exogenous neurotrophic factors may be potent therapeutics capable of trophic support by extending axonal sprouting and dendritic remodeling in the degenerating brain. 24 The addition of GCE to neurons resulted in the extension of axonal and dendritic arbors with robust branching; thus, it may be effective for further stabilization of synaptic plasticity in neurons. Consequently, this marine alga might be useful for developing medicinal food or drugs that have an effect on restoring neuronal cells from age-associated neurodegeneration. However, further studies are needed to establish whether trophic effects of GCE could potentiate spinogenesis and synaptogenesis of hippocampal neurons in later stages of culture in vitro and in vivo.
G. chorda has been considered an economically important seaweed species in agar-producing countries, including Korea, Japan, and China. The amount of Gracilaria collected from natural populations in Korea during 2013 was 13 tons (wet weight), but the amount produced by aquaculture was not recorded. 25 As a rich source of phytochemicals and because of its nontoxic nature, G. chorda may be useful as a dietary supplement for people who are prone to age-associated disorders involving memory and learning deficits.
Footnotes
Acknowledgment
This work was supported by a research grant of Pukyong National University (2016).
Author Disclosure Statement
No competing financial interests exist.