
Abstract
The aim of this study was to evaluate the antidiabetic properties of collagen hydrolysates (CHs). CHs exhibited dipeptidyl peptidase-IV inhibitory activity and stimulated glucagon-like-peptide-1 (GLP-1) secretion in vitro. We also determined whether CHs improve glucose tolerance in normal mice. Oral administration of CHs suppressed the glycemic response during the oral and intraperitoneal glucose tolerance tests (OGTT and IPGTT), but the effects were weaker in IPGTT than in OGTT. CHs had no effect on the gastric emptying rate. A pretreatment with the GLP-1 receptor antagonist, exendin 9–39 (Ex9), partially reversed the glucose-lowering effects of CHs, but only when coadministered with glucose. CHs administered 45 min before the glucose load potentiated the glucose-stimulated insulin secretion. This potentiating effect on insulin secretion was not reversed by the pretreatment with Ex9, it appeared to be enhanced. These results suggest that CHs improve glucose tolerance by inhibiting intestinal glucose uptake and enhancing insulin secretion, and also demonstrated that GLP-1 was partially involved in the inhibition of glucose uptake, but not essential for the enhancement of insulin secretion.
Introduction
T
After the ingestion of food, the intestines secrete a range of hormones with important local and distant actions, including the control of insulin secretion and appetite. 6 Glucose-dependent insulinotropic polypeptide (GIP) and glucagon-like peptide-1 (GLP-1) are two primary incretin hormones that increase insulin secretion from islet β cells in a glucose-dependent manner. 7,8 Both incretin hormones are rapidly inactivated by the ubiquitously expressed serine protease, dipeptidyl peptidase-IV (DPP-IV), which cleaves the N-terminal dipeptide from these hormones. 7,8 Long-acting GLP-1 analogues and DPP-IV inhibitors are now being extensively used to treat T2DM. 9
The enzymatic hydrolysates of gelatin are called collagen peptides or collagen hydrolysates (CHs), and have been used in diverse fields, including the food, cosmetic, biomedical, and pharmaceutical industries. 10,11
CHs have been shown to exert beneficial biological effects in the treatment of joint disorders 12 and skin damage 13 ; however, limited information is available on their effects on glucose metabolism. 14,15 Protein hydrolysates are known to have insulinotropic actions 16,17 and stimulate the release of GLP-1. 18,19 Therefore, we hypothesized that CHs may be useful as dietary supplements for the prevention of T2DM.
In the present study, we examined the antidiabetic properties of CHs in vitro and in vivo. We showed that CHs exerted inhibitory effects on DPP-IV activity and potentiated the secretion of GLP-1 in vitro. We also demonstrated that the oral administration of CHs improved glucose tolerance in normal mice. Furthermore, we investigated the involvement of GLP-1 in the glucose-lowering mechanisms of CHs.
Materials and Methods
Materials
The CHs used in this study (Wellnex® Type D), which were commercially available, were kindly gifted by Nitta Gelatin, Inc. (Osaka, Japan). These CHs were enzymatic hydrolysates of fish (Oreochromis sp.) scale gelatin and of food grade. Recombinant human DPP-IV was purchased from Enzo Life Sciences (Farmingdale, NY, USA). Gly-Pro-βNA was obtained from Bachem AG (Bubendorf, Switzerland). Liraglutide and Exendin 9–39 (Ex9) were purchased from Novo Nordisk (Copenhagen, Denmark) and AnaSpec (San Jose, CA, USA), respectively.
Determination of DPP-IV inhibitory activity
DPP-IV activity was determined in 96-well microplates using Gly-Pro-βNA as a surrogate substrate. CHs were reconstituted in 20 mM Tris–HCl buffer (pH 7.0). The mixture (90 μL), consisting of 70 μL of 20 mM Tris–HCl buffer (pH 7.0), 10 μL of DPP-IV (diluted with the same Tris buffer to 0.0125 U mL), and 10 μL of the test sample, was preincubated at 37°C for 5 min. After the preincubation, 10 μL of the substrate solution was added to initiate the reaction, and the reaction mixture was incubated at 37°C for 20 min. The enzymatic reaction was terminated by the addition of 50 μL Fast Garnet GBC (1 mg/mL) solution, dissolved in 1 M sodium acetate buffer (pH 4.0) containing 10% (v/v) Triton X-100. Absorbance was measured at 550 nm with a microplate reader.
Cell culture and GLP-1 secretion studies
Human enteroendocrine NCI-H716 cells were obtained from the American Type Culture Collection (Manassas, VA, USA). Two days before the experiment, cells were seeded on 96-well culture plates precoated with poly-
Mice
Male C57BL/6J mice (8 weeks) were purchased from Japan SLC (Hamamatsu, Japan) and acclimated to the facility for 1 week before experimental use. Mice were fed standard chow and maintained under controlled environmental conditions (12-h light–12-h dark cycle, 23°C ± 2°C, and 55% ± 10% humidity). All experiments were conducted on mice between 9 and 13 weeks of age. Animal procedures were approved by the Animal Care and Use Committee of Setsunan University and were conducted in accordance with the guidelines of the Japanese Pharmacological Society.
Glucose tolerance test
The oral glucose tolerance test (OGTT) and intraperitoneal glucose tolerance test (IPGTT) were performed on mice fasted for 16 h. Blood glucose levels were monitored in single drops of blood obtained over the time course from the tip of the tail at 0, 15, 30, 60, 90, and 120 min after a 2 or 4 g/kg glucose load using a glucose meter (Sanwa Kagaku Kenkyusho, Aichi, Japan). The area under the glucose curve between 0 and 120 min (AUC0–120 min) was calculated for each group of mice.
Gastric emptying
We used a modification of the techniques described earlier for gastric emptying measurement. 21 Briefly, mice were administered glucose solution containing 0.1 mg/mL phenol red and then sacrificed 20 min later by cervical dislocation. The cardia and pylorus of the stomach were clamped to prevent the leakage of phenol red. The isolated stomach was transferred to 10 mL of 0.1 N NaOH in a 50-mL Falcon tube and cut into small fragments. This mixture was homogenized for 30 sec and allowed to stand at room temperature for 1 h. Tissue debris was removed by centrifugation at 2300 g for 15 min. Five milliliters of supernatant was placed into a 15-mL Falcon tube and precipitated with 0.5 mL of 20% trichloroacetic acid, then centrifuged at 2300 g for 30 min. Two milliliters of the supernatant was transferred to another test tube and mixed with 2 mL of 0.5 M NaOH to develop the maximum intensity of the color. The solutions were assayed with a spectrometer at 560 nm. The percentage of gastric emptying was calculated as (infusion-remained/infusion) × 100%.
Measurement of plasma insulin and active GLP-1
Blood was collected 15 min after the administration of glucose by intracardiac puncture under deep anesthesia with sodium pentobarbital (70 mg/kg, i.p.). Blood samples were immediately transferred into ice-cooled tubes containing EDTA-2K (final concentration, 1 mg/mL) and a DPP-IV inhibitor (10 μL per milliliter of blood; DPP4-010: EMD Millipore, Billerica, MA, USA) for the measurement of insulin and active GLP-1 concentrations. Plasma was separated by centrifugation at 4°C and stored at −80°C until measurements. Plasma concentrations of insulin and active GLP-1 were measured using the Mouse Insulin Assay Kit (Morinaga, Yokohama, Japan) and the GLP-1, Active Form Assay Kit (IBL), respectively.
Administration of CHs or liraglutide
CHs (2–3 g/kg) or vehicle (water) were orally coadministered with glucose or administered 45 min before the ingestion of glucose. In some experiments, we compared the effects of CHs with those of liraglutide, a GLP-1 receptor agonist. Liraglutide (50 μg/kg) or vehicle (saline) was subcutaneously administered 45 min before the ingestion of glucose.
Administration of Ex9 in OGTT
In some experiments, the GLP-1 receptor antagonist, Ex9, was used to investigate the possible involvement of GLP-1 in the glucose-lowering effects of CHs. Ex9 (1 nmol/mouse, i.p.) was administered 30 min before the ingestion of glucose in the case of the coadministration of CHs with glucose, or 60 min before the ingestion of glucose in the case of the administration of CHs or liraglutide 45 min before the ingestion of glucose.
Statistical analysis
All values are expressed as the mean ± SEM. Statistical analysis was performed using Student's t-test for single comparisons and a one-way ANOVA followed by Tukey's HSD test for multiple comparisons. Differences were considered significant at P < .05.
Results
Effects of CHs on DPP-IV activity
DPP-IV inhibitory activities at various concentrations of CHs were shown in Figure 1a. The IC50 value was determined and found to be 0.77 mg/mL.

Effects of CHs on DPP-IV activity
Effects of CHs on GLP-1 secretion in NCI-H716 cells
CHs also stimulated the secretion of GLP-1 from NCI-H716 cells in a dose-dependent manner, with the maximum level of secretion reaching 258% ± 3.23% of the control value in the presence of 1% CHs (Fig. 1b).
Effects of CHs on the glycemic response during OGTT and IPGTT
The oral administration of CHs markedly attenuated the glycemic response during OGTT (Fig. 2a–d). The administration of CHs 45 min before the ingestion of glucose significantly decreased the AUC0–120 min values at 3 g/kg in the case of a 2 g/kg glucose load (Fig. 2a), and at 2–3 g/kg in the case of a 4 g/kg glucose load (Fig. 2b). Likewise, the coadministration of CHs with glucose significantly decreased the AUC0–120 min values at 2–3 g/kg in the case of a 2 g/kg glucose load (Fig. 2c), and at 3 g/kg in the case of a 4 g/kg glucose load (Fig. 2d). Since the inhibition of glucose uptake attenuated the glycemic response during OGTT, we conducted IPGTT to eliminate possible effects on glucose absorption. The administration of CHs 45 min before the ingestion of 2 g/kg glucose significantly attenuated the glycemic response during IPGTT; however, the degree of inhibition (18.2%) was lower than that in the case of OGTT (28.4%) (Fig. 2a, e).
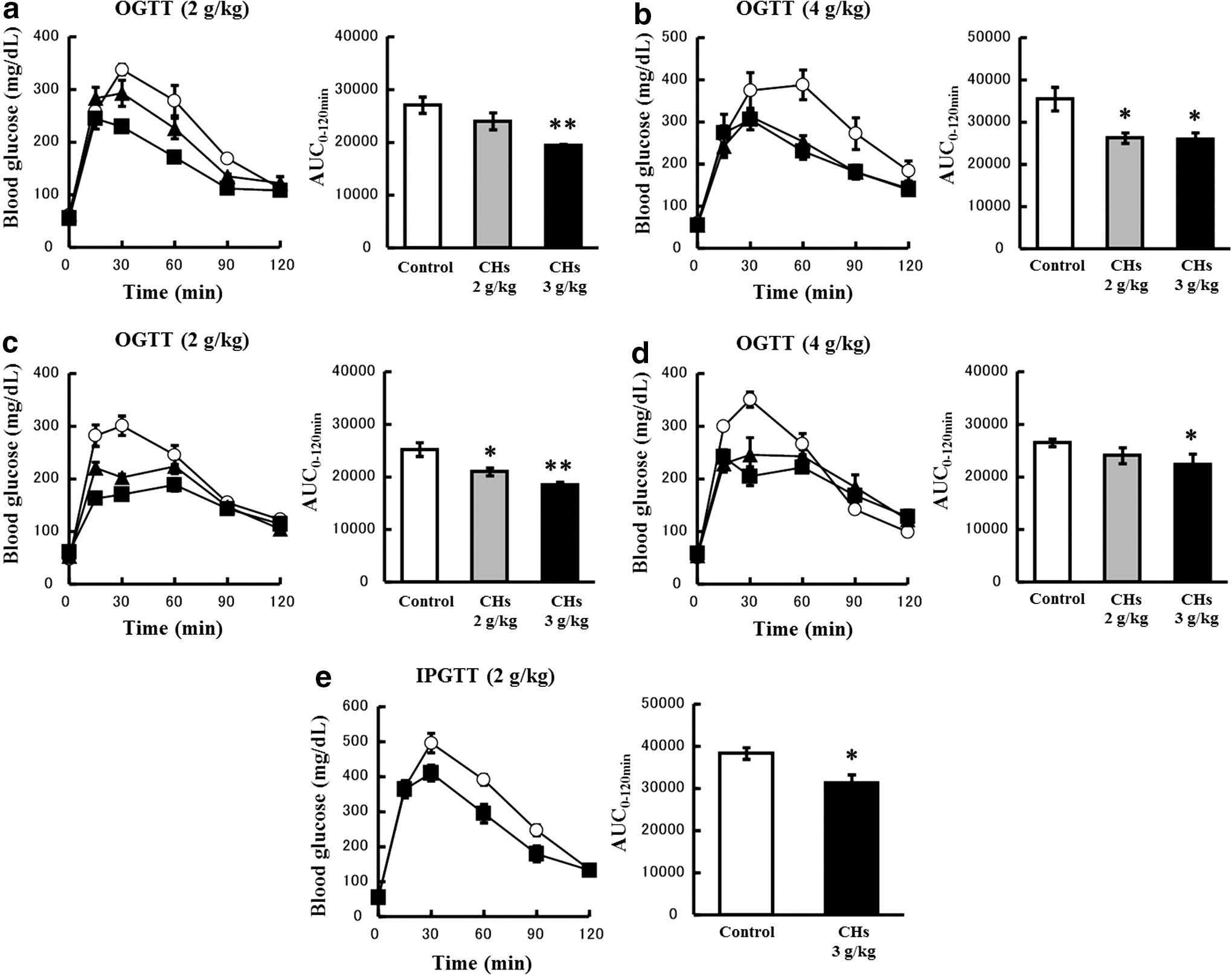
Effects of CHs on the glycemic response during OGTT and IPGTT in mice. Mice were fasted for 16 h and orally loaded with 2 g/kg glucose
Effects of CHs on the gastric emptying rate
No significant differences were noted between mice administered CHs and control mice, regardless of the timing of the administration of CHs and the loading dose of glucose (Fig. 3a–d).
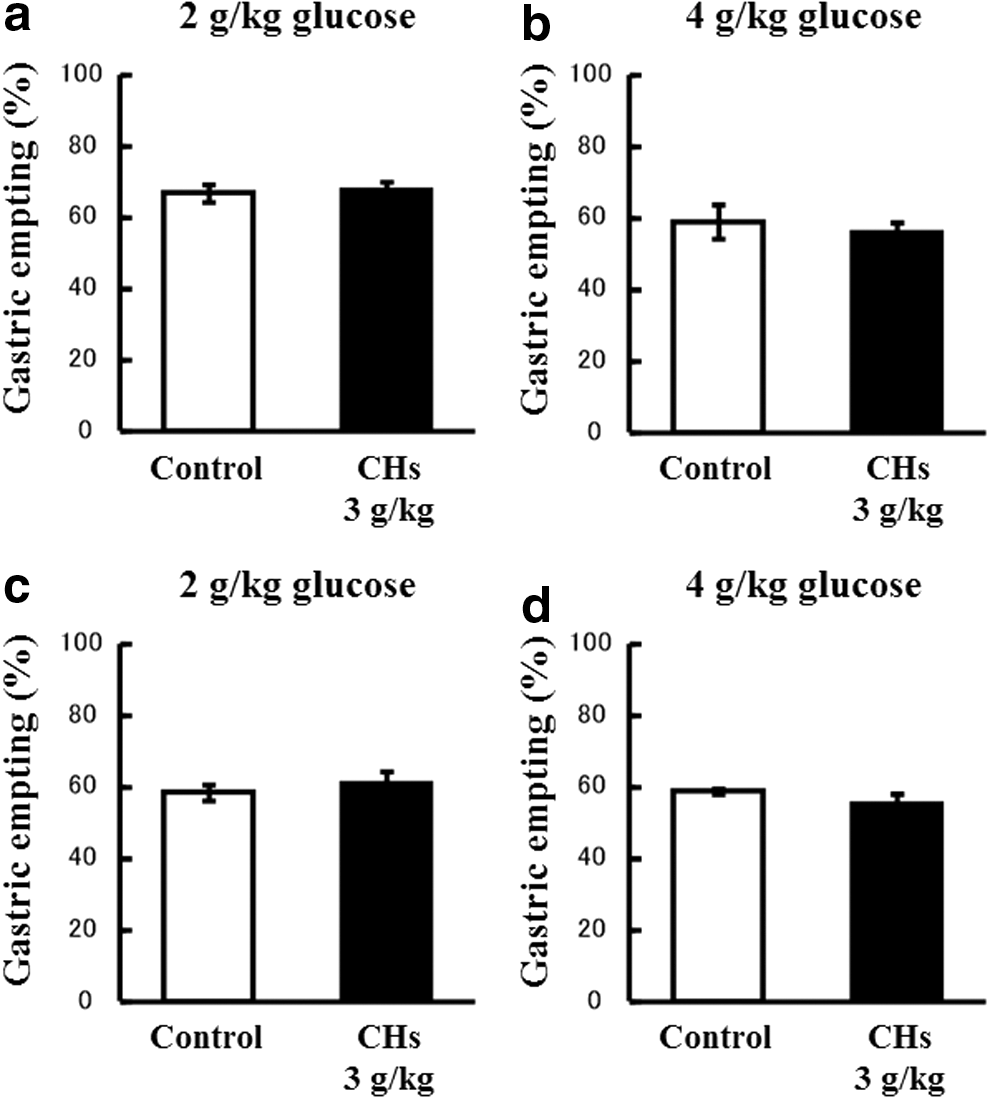
Effects of CHs on the gastric emptying rate. The percentage of gastric emptying was calculated 20 min after the 2 g/kg
Effects of CHs on glucose-stimulated active GLP-1 secretion
The administration of CHs (3 g/kg) 45 min before the ingestion of glucose did not affect active GLP-1 levels in the plasma in the case of a 2 g/kg glucose load or a 4 g/kg glucose load (Fig. 4a). On the other hand, the coadministration of CHs (3 g/kg) with glucose significantly elevated active GLP-1 levels in the plasma after 15 min in the case of a 4 g/kg glucose load, but not in the case of a 2 g/kg glucose load (Fig. 4b).

Effects of CHs on the glucose-stimulated secretion of active GLP-1 and insulin in mice. Mice were fasted for 16 h and orally loaded with glucose (2 or 4 g/kg). CHs (black bars, 3 g/kg) or vehicle (white bars, water) were coadministered with glucose
Effects of CHs on glucose-stimulated secretion of insulin
The administration of CHs (3 g/kg) 45 min before the ingestion of glucose significantly increased plasma insulin levels 15 min after the ingestion of glucose (Fig. 4c). On the other hand, the coadministration of CHs (3 g/kg) with glucose significantly elevated plasma insulin levels after 15 min in the case of a 2 g/kg glucose load, but not in the case of a 4 g/kg glucose load (Fig. 4d).
Involvement of GLP-1 in glucose-lowering effects of CHs
We then examined the glucose-lowering effects of liraglutide, a GLP-1 receptor agonist, and CHs in the presence or absence of the GLP-1 receptor antagonist, Ex9. The pretreatment with Ex9 enhanced the glycemic response to a 2 g/kg oral glucose load (Fig. 5b, d), but not to a 4 g/kg oral glucose load (Fig. 5a, c, e). The subcutaneous administration of liraglutide 45 min before the administration of 4 g/kg glucose markedly attenuated the glycemic response (Fig. 5a). The glucose-lowering effect of liraglutide was almost completely reversed by the pretreatment with Ex9 (Fig. 5a). However, in the case of the administration of CHs 45 min before the ingestion of glucose, the glucoregulatory actions of CHs were not affected by the pretreatment with Ex9, regardless of the glucose doses (2 or 4 g/kg) (Fig. 5b, c). On the other hand, in the case of the coadministration of CHs with glucose, the glucose-lowering effects of CHs were significantly reversed by the pretreatment with Ex9 (Fig. 5d, e). This phenomenon was marked 60 min after the coadministration, but was hardly observed 30 min after the coadministration (Fig. 5d, e).
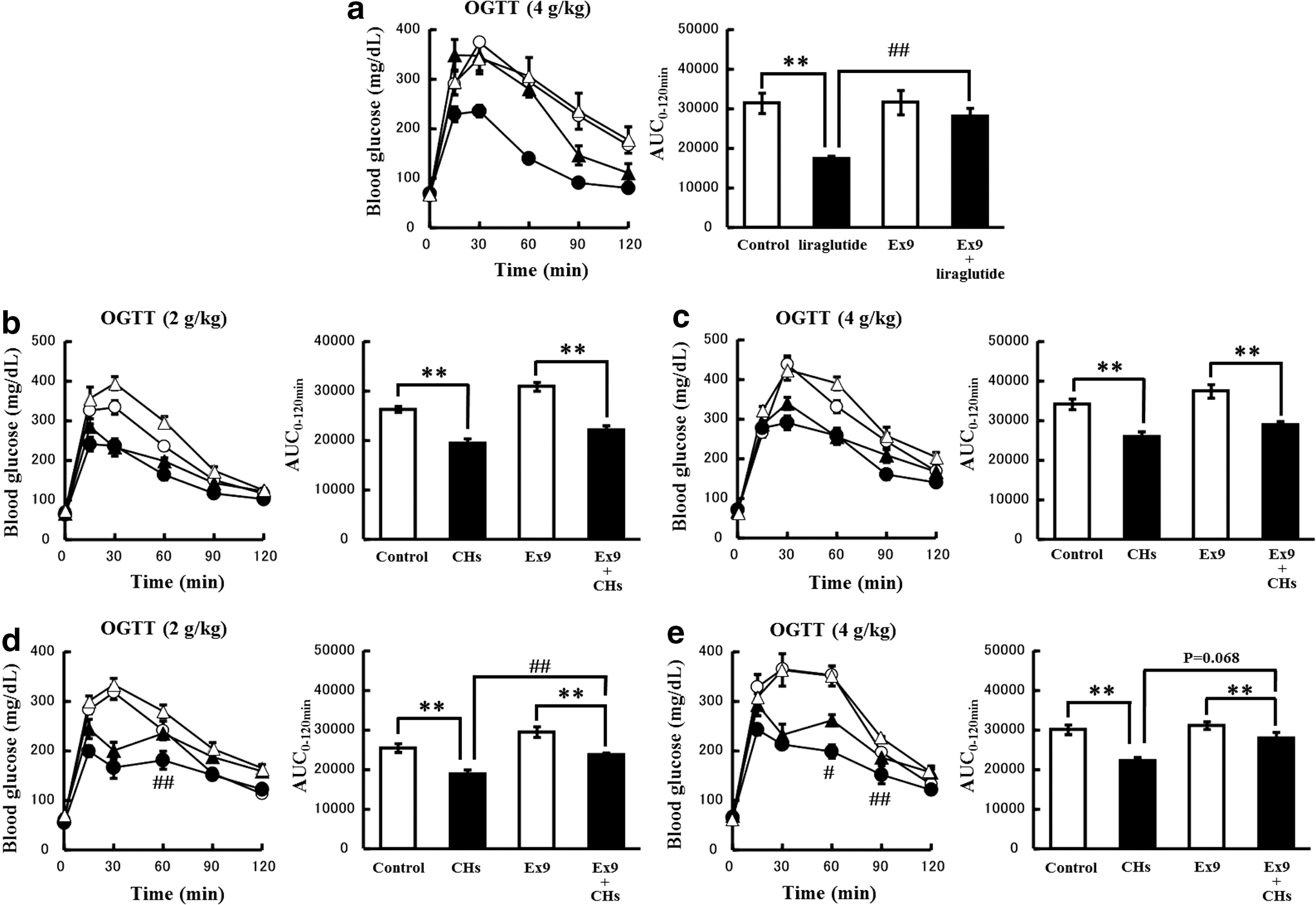
Involvement of GLP-1 receptors in the glucose-lowering effects of CHs. Mice were fasted for 16 h and orally loaded with 2 g/kg glucose
Involvement of GLP-1 in CHs-enhanced insulin secretion
Liraglutide, regardless of whether administered with or without Ex9 pretreatment, did not induce any significant effects on the insulin secretion response to a 4 g/kg oral glucose load (Fig. 6a). On the other hand, as described above, CHs administered 45 min before the 4 g/kg glucose load potentiated the glucose-stimulated insulin secretion. This potentiating effect on insulin secretion was not reversed by the pretreatment with Ex9, it appeared to be enhanced (Fig. 6b).
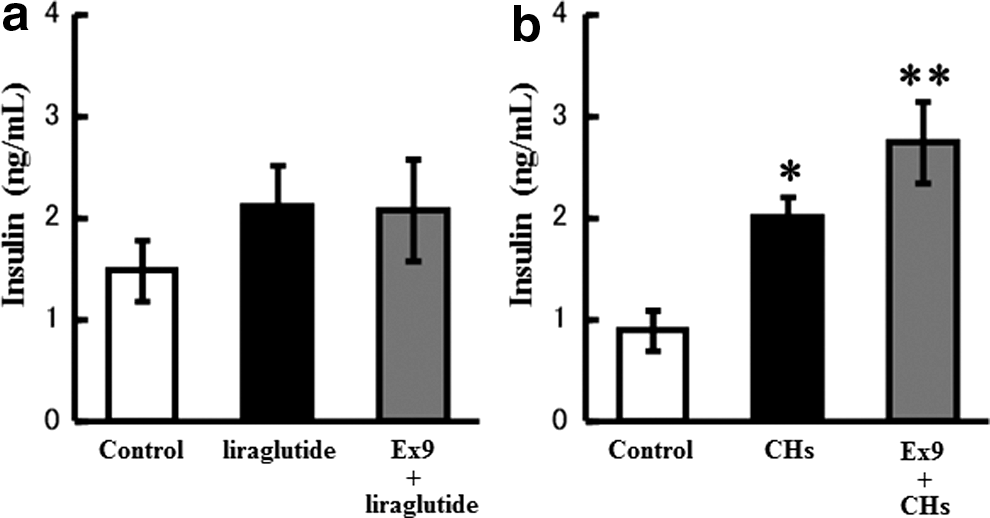
Involvement of GLP-1 in CH-enhanced insulin secretion. Mice were fasted for 16 h and orally loaded with 4 g/kg glucose. Liraglutide (50 μg/kg, s.c.,
Discussion
Our in vitro results (Fig. 1) were consistent with the findings of previous studies, indicating that the protein hydrolysates derived from various protein sources exhibited inhibitory activity toward DPP-IV 5, 22 –25 and directly affected the GLP-1 stimulation. 18,19,26,27 These results suggested that CHs have the potential to be antidiabetic supplements.
In the present study, we demonstrated that the administration of CHs improved glucose tolerance in normal mice (Fig. 2). This is consistent with previous findings in which the coadministration of some proteins or protein hydrolysates with glucose improved glucose tolerance in rodents. 28 –30 On the other hand, circulating active GLP-1 levels significantly increased under restricted conditions only (coadministration of 3 g/kg CHs with 4 g/kg glucose, Fig. 4b). Gunnarsson et al. reported that the coadministration of 75 mg whey protein with 75 mg glucose to normal C57BL/6J mice (equal to 3–3.75 g/kg each) led to significantly higher plasma insulin and active GLP-1 levels than those with glucose alone. 28 These results suggested that a high glucose dose may be needed for a significant increase in active GLP-1 levels when CHs are coadministered with glucose.
We also compared the effects of CHs with those of liraglutide to reveal the involvement of GLP-1 in the glucose-lowering mechanisms of CHs. Liraglutide markedly delayed the gastric emptying rate (data not shown), but had no effect on insulin secretion 15 min after the glucose load in the present study (Fig. 6a). When glucose is administered orally, short-acting GLP-1 agonists have been shown to predominantly lower postprandial glucose excursions by inhibiting gastric emptying and intestinal glucose uptake, with negligible effects on insulin secretion. 31 Since we used liraglutide, which is a long-acting agonist, in single administration studies, our results were consistent with previous findings. In the present study, the significant increase observed in active GLP-1 levels was not followed by a significant increase in insulin levels (Fig. 4d). The increase in plasma levels of GLP-1 may delay the absorption of glucose into the blood from the intestines and subsequently reduce glucose-stimulated secretion of insulin.
The glucose-lowering effects of CHs were weaker in IPGTT than in OGTT (Fig. 2a, e). CHs also had no effect on the gastric emptying rate (Fig. 3a–d). These results suggest that the majority of the glucose-lowering effects of CHs were due to the inhibition of intestinal glucose uptake. The glucose-lowering effects of CHs when coadministered with glucose were partially reversed by the pretreatment with Ex9 (Fig. 5d, e), suggesting that the inhibition of intestinal glucose uptake by CHs was due not only to GLP-1, but also to other factors.
When CHs were administered 45 min before the glucose load, the glucose-lowering effects of CHs were not altered by the pretreatment with Ex9. Furthermore, the potentiating effects of CHs on insulin secretion were not reversed by the pretreatment with Ex9, they appeared to be enhanced. Although the reasons for this remain obscure, one possible mechanism is that the blockade of the GLP-1 receptor increased intestinal glucose uptake and compensatory insulin secretion, resulting in no change in blood glucose levels. In any case, our results suggest that GLP-1 is not essential for the effects of CHs on the enhancement of insulin secretion. Higuchi et al. 29 showed that the oral administration of Zein hydrolysates (ZeinH) 15 min before a glucose injection (1 g/kg, i.p.) reduced the glycemic response 15 and 30 min after the glucose injection, and that the glucose-lowering effects of ZeinH were attenuated by the pharmacological blockade of either GIP or GLP-1 receptors. Glucose-induced insulin secretion (GSIS) is known to be modulated by many factors, including hormones such as GLP-1 and GIP, neurotransmitters, including acetylcholine, and nutrients such as fatty acids. 32,33 Further studies are needed to explore the mechanisms underlying the GLP-1-independent stimulatory effects of CHs on GSIS.
Conclusion
We demonstrated that CHs possess potent antidiabetic properties as illustrated in Figure 7. CHs inhibited DPP-IV activity and stimulated the secretion of GLP-1 in vitro. The oral administration of CHs improved glucose tolerance by inhibiting intestinal glucose uptake and enhancing insulin secretion in mice. Furthermore, GLP-1 was partially involved in the inhibition of glucose uptake, but was not essential for the enhancement of insulin secretion.

Glucose-lowering mechanisms of CHs during OGTT.
Footnotes
Acknowledgment
This work was supported in part by a grant from Nitta Gelatin, Inc., Japan.
Author Disclosure Statement
No competing financial interests exist.