
Abstract
Krill oil is a novel, commercially available marine oil rich in long-chain polyunsaturated omega-3 fatty acids, particularly eicosapentaenoic acid and docosahexaenoic acid. Compared with fish oil, the effects of krill oil supplementation on human health and its underlying action mechanisms are currently poorly understood. In the present study, we examined the effect of krill oil supplementation on metabolic parameters of mice fed a high-fat diet (HFD). Krill oil supplementation in mice fed a HFD for 10 weeks resulted in an ∼15% lower body weight gain and a dramatic suppression of hepatic steatosis. These effects were associated with significantly lower serum triglyceride and low-density lipoprotein-cholesterol levels. We further uncovered a novel underlying mechanism, showing that AMP-activated protein kinase, a master regulator of glucose and lipid metabolism, mediates the beneficial effects of krill oil.
Introduction
M
Evidence accumulating over the past several decades suggests that dietary intake of long-chain omega-3 polyunsaturated fatty acids (n-3 PUFAs) improves health by reducing the risk of cardiovascular disease, obesity, inflammation, and neurodegeneration. 6 –10 Among n-3 PUFAs, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) are well known bioactive compounds that are highly enriched in marine-derived food sources, including fatty fish, shellfish, and krill. 6 –10 The American Heart Association dietary guideline suggests the intake of at least 400–500 mg of DHA and EPA per week, which can be achieved by consuming two servings of fish weekly; they suggest even higher intake of n-3 PUFAs for patients with coronary heart diseases. 11,12 Fish oil extracted from salmon, sprat, and anchovies has been widely consumed as a source of DHA and EPA. In addition, krill oil (krill oil), extracted from Antarctic krill (Euphausia superba), a marine crustacean with the most abundant biomass on earth, has recently been recognized as a new source of n-3 PUFAs. 13 –16 Krill oil differs from most fish oils available on the market in that a majority of its n-3 PUFAs are esterified in phospholipids, whereas n-3 PUFAs in fish oil are incorporated into the triglycerol (TG). 13,17 –19 Several studies have reported that DHA and EPA levels in tissues are higher when they are delivered in phospholipid form than in TG form. 19,20 Krill oil was also found to be Generally Recognized As Safe (GRAS) by the American Food and Drug Administration. 9 Many preclinical and clinical studies have confirmed the safety of krill oil and its lack of adverse effects and shown that consumption of krill oil increases plasma EPA and DHA levels in normal, as well as obese, individuals. 21 These beneficial effects of krill oil on health have motivated intensive research efforts, but the mechanisms of action of krill oil are still poorly understood.
In the present study, we demonstrated that krill oil exerts antiobesity effects on mice fed a high-fat diet (HFD) by lowering the levels of TGs and low-density lipoprotein (LDL)-cholesterol. In particular, hepatic steatosis was efficiently prevented by krill oil supplementation. We further revealed the novel mechanism for krill oil action. AMP-activated protein kinase (AMPK) is a master regulator of lipid and glucose metabolism 22 –25 and has been recognized as a therapeutic target for the treatment of metabolic syndrome. 26 –29 Therefore, we tested whether AMPK is involved in the action of krill oil and demonstrate that AMPK mediates the beneficial effect of krill oil.
Materials and Methods
Preparation of krill oil
Krill (Euphausia Superba) was crushed and mixed with water and a protease isolated from Bacillus subtilis Y-108, reported to be highly effective in deproteinizing crustacean wastes, 30 and further incubated at 40°C for 3 h. After removing proteins, the liquid was filtered to remove krill shells, and the pH was adjusted to a neutral 7.5, yielding a water-insoluble fraction. This fraction was further filtered to remove small proteins and distilled to reduce water content to less than 1%. No organic solvents were used at any point in the process. The resulting viscous red krill oil was analyzed for lipid content according to the US Pharmacopeial Convention (USP). The lipid composition of krill oil, expressed as a percentage of total lipid, was as follows: phospholipid, 42.3%; TG, 30.6%; free fatty acids, 12.2%; sterol esters, 6.2%; cholesterol/sterols, 5.8%; and unknown neutral lipids, 2.9%. The composition of fatty acids is summarized in detail in Table 1. UV spectrophotometry analyses detected the antioxidant, astaxanthin, at a concentration of 191 mg/kg. Escherichia coli, Salmonella, coliform group, molds, and yeast were not detected. Levels of heavy metals, including mercury (<0.005 mg/kg), cadmium (<0.05 mg/kg), and lead (<0.05 mg/kg), were far below lower limit values.
Animal models
All animal experiments were approved by the Institutional Animal Care and Use Committee of Kyung Hee University. Male C57BL/6N mice were obtained from Taconic Farms (Hudson, NY) at 3 weeks of age and housed at 21 ± 2°C, 50% ± 5% relative humidity, with a 12-h light/12-h dark cycle. After mice consumed a commercial diet for 1 week, they were divided into three weight-matched groups (n = 10/group): a normal diet (ND), which provides 10% of calories as fat (D12450B normal fat diet [NFD]; Research Diets, New Brunswick, NJ); a HFD, which provides 40% of calories as fat (D12492 HFD; Research Diets); and HFD + krill oil, added at a 2% (w/w) ratio, which also provides 40% of calories as fat. Daily food intake was restricted to 2.5 g per mouse, and body weights of mice in the three groups were recorded daily. At the end of the experimental period, mice from each group were anesthetized with diethyl ether after an initial 12-h fasting period. Blood was collected from the aorta into an EDTA-coated tube. Thereafter, mice were dissected and their tissue weights were measured. The specimens were immediately frozen in liquid nitrogen and stored at −80°C.
Micro CT image
At the end of the experimental period, micro-CT images were taken. Transverse micro-CT images were taken from L2 lumbar spine section, and 3D scan images were reconstructed by accumulating visceral and subcutaneous fat areas of transverse micro-CT images from L2 to L5 lumbar spine sections.
Antibodies and reagents
Antibodies against phospho(Thr172)-AMPKα, phospho(Ser79)-ACC (acetyl-CoA carboxylase), total ACC, and FAS (fatty acid synthase) were purchased from Cell Signaling Technology (Danvers, MA). Antibodies against SREBP-1c, GAPDH, lamin B, and horseradish peroxidase (HRP)-conjugated secondary antibodies were purchased from Santa Cruz Biotechnology (Dallas, TX). All cell culture media and supplements were obtained from Thermo Fisher Scientific (Waltham, MA). Transfection reagents were purchased from Intron Biotechnology (Gyeonggi-do, Korea).
Cell culture
The Chang liver cell line was maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and antibiotics and cultured at 37°C in 95% air and 5% CO2.
Immunoblot analysis
Total proteins from liver tissue and Chang cells were extracted in RIPA lysis buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.1% SDS, 0.5% sodium deoxycholate, 5 mM sodium fluoride, 2 mM sodium orthovanadate, 1 mM PMSF, and protein inhibitor cocktail). Nuclear and cytosolic fractions were prepared using a mild lysis buffer (10 mM HEPES pH 7.9, 1.5 mM MgCl2, 10 mM KCl, 0.5 mM DTT, and 0.05% NP40) and RIPA lysis buffer. The protein concentration of the lysate was determined by Bradford assay. For immunoblotting, total cellular protein or subcellular fractions were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). After transferring proteins to nitrocellulose membranes, membranes were blocked with tris-buffered saline (TBS) containing Tween 20 (10 mM Tris-HCl pH 7.4, 150 mM NaCl, and 0.1% Tween 20) and 5% nonfat dry milk and incubated overnight at 4°C with primary antibodies diluted in blocking buffer. The membranes were washed and incubated with the appropriate secondary antibodies for 1 h at room temperature. HRP-conjugated secondary antibodies were detected using enhanced chemiluminescence reagents.
Reverse transcription-polymerase chain reaction
Total RNA was extracted from liver tissue or Chang cells using TRIzol reagent, and cDNA was synthesized by reverse transcription (RT) from 1 μg of total RNA using a cDNA Synthesis Kit, according to the manufacturer's instructions (Enzynomics, Daejeon, Korea). The primer pairs used for polymerase chain reaction (PCR) are as follows: AMPKα, 5′-TGT CAC AGG CAT ATG GT-3′ (forward) and 5′-ATT CAC CAT CTG ACA TCA T-3′ (reverse); ACC1, 5′-GTC AGC GGA TGG GCG GAA TG-3′ (forward) and 5′-CGC CGG ATG CCA TGC TCA AC-3′ (reverse); ACC2, 5′-GCT GCG GTC AAG TGT ATG CG-3′ (forward) and 5′-CAC TGA TGC ATT TGC CCT GG-3′ (reverse); SREBP-1c, 5′-GCG CTA CCG GTC TTC TAT CA-3′ (forward) and 5′-TGC TGC CAA AAG ACA AGG-3′ (reverse); FAS, 5′-GAT CCT GGA ACG AGA ACA C-3′ (forward) and 5′-AGA CTG TGG AAC ACG GTG GT-3′ (reverse); and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), 5′-ACG GCC GCA TCT TCT TGT GC-3′ (forward) and 5′-CGG CAG AAG GGG CGG AGA TG-3′ (reverse). The following thermocycling conditions were used for PCR: 2 min at 95°C, followed by 25 cycles of 95°C for 30 sec, 55°C for 30 sec, 72°C for 30 sec, and a final 2-min incubation at 72°C. PCR products were analyzed by agarose gel electrophoresis using 1% gels containing ethidium bromide. RT-PCR data were normalized to GAPDH mRNA levels in each sample.
Histological analysis
Liver tissues from three groups of mice fed ND, HFD, or HFD + krill oil were perfused with phosphate-buffered saline (PBS), frozen in liquid nitrogen immediately, and stored at −80°C. Tissue sections (4-μm thick) were prepared with a cryostat microtome (Leica, Wetzlar, Germany) and mounted on gelatin-coated glass slides. After fixing with 4% paraformaldehyde, sections were stained with hematoxylin–eosin (H–E) for microscopic assessment (Olympus, Tokyo, Japan). The frozen liver sections were stained with Oil Red O for the histological examination of fat droplets.
Biochemical analysis
The concentrations of blood glucose, high density lipoprotein (HDL)-cholesterol, LDL-cholesterol, and TGs were determined using an Accu-Chek Active Kit (Roche Diagnostics, GmbH, Germany) and automated analyzer (SMARTLAB, Mannheim, Germany). The concentration of adiponectin was measured using an Enzyme-linked Immunosorbent Assay (ELISA) Kit (Otsuka Pharmaceutical Co., Ltd. Tokyo, Japan).
Statistical analysis
Results are expressed as mean ± standard deviation. Differences between groups were examined for statistical significance using Student's t-test (RStudio, version 0.99.893). Differences were considered to be significant for P values <.05.
Results
Krill oil reduces the body weight and abdominal fat content of HFD-fed mice
To determine the physiological effects of krill oil on body weight, we divided animals into three weight-matched groups (n = 10/group) and maintained each group on one of three diets: a ND, which provided 10% of calories as fat; a HFD, which provided 40% of calories as fat; and HFD + krill oil added at a 2% (w/w) ratio, which also provided 40% of calories as fat. When allowed free access to food, mice in the HFD + krill oil group tended to consume a significantly larger amount of food daily (∼3.8 g) than those in ND or HFD groups, which consumed 2.5 g on average (Fig. 1A, left panel), a difference in consumption that may reflect the distinctive odor and taste of krill oil. Because the unequal amount of food intake among three groups might interfere with data interpretation, we chose to supply an equal amount of food for each group, restricting the daily food supply to 2.5 g per mouse and then monitored daily food consumption during the experimental period. Under these conditions of equal daily food consumption (Fig. 1A, right panel), the body weights of HFD mice markedly increased compared with those of ND mice, whereas weights of mice administered HFD + krill oil increased significantly and were by almost 15% lower at the end of 10 weeks (Fig. 1B). An anatomical examination of HFD + krill oil mice showed a noticeably lower body fat mass compared with HFD mice (Fig. 1C), and transverse micro-computed tomography (CT) and three-dimensional (3D) images revealed that visceral and subcutaneous fat content in the abdominal region were lower by 20% in HFD + krill oil mice compared with mice in the HFD group (Fig. 1D, E). These results show that krill oil possesses strong antiobesity properties.

The effects of krill oil on body weight and abdominal fat content in HFD-fed mice.
Krill oil reduces fat content in the liver and lowers serum levels of TG and LDL-cholesterol in HFD-fed mice
An examination of the effects of krill oil on major organs revealed the most dramatic effects in the liver, showing that krill oil lowers liver masses of HFD-fed mice by 35% when normalized for body weight (Fig. 2A, B). Notably, a histological examination of liver sections stained with both H–E and Oil Red O showed that consumption of krill oil considerably suppressed hepatic steatosis in mice with diet-induced obesity (Fig. 2C).
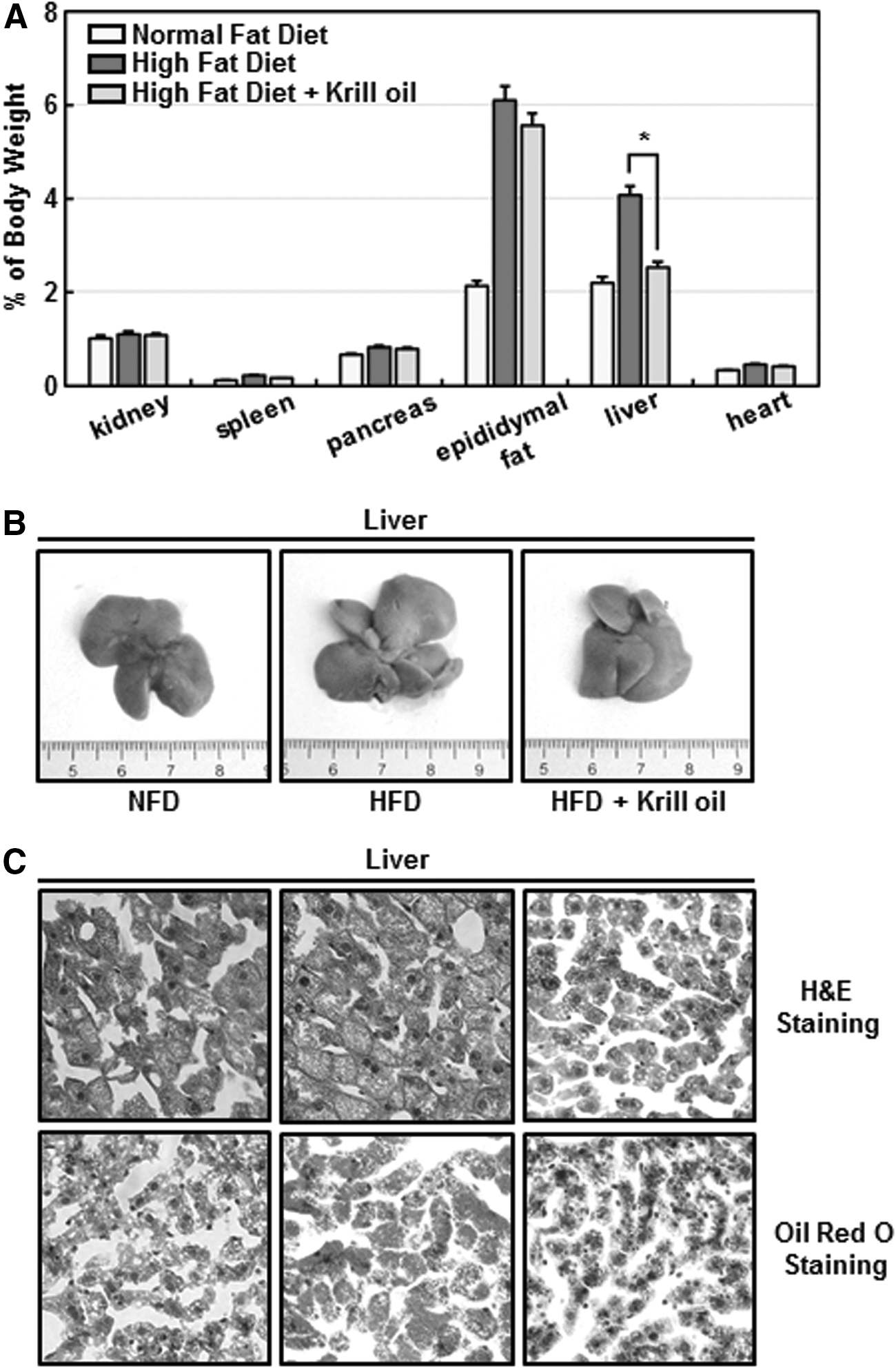
Krill oil reduces the weight and fat content of mouse livers.
To investigate glucose and lipid homeostasis in krill oil-fed mice, we performed biochemical analyses of various parameters of glucose and lipid metabolism in the sera of mice from each diet group. Consumption of krill oil did not significantly change fasting blood glucose (Fig. 3A) or total-cholesterol (Fig. 3B) or HDL-cholesterol (Fig. 3C) levels between HFD mice and mice in the HFD + krill oil group, but did significantly decrease LDL-cholesterol (Fig. 3D) and TG (Fig. 3E) levels. Adiponectin concentrations were increased in HFD mice compared with mice in the NFD group, but no significant decrease was observed in HFD + krill oil mice (Fig. 3F). These results indicate that krill oil consumption markedly lowers TG and LDL-cholesterol levels in mice with diet-induced obesity.
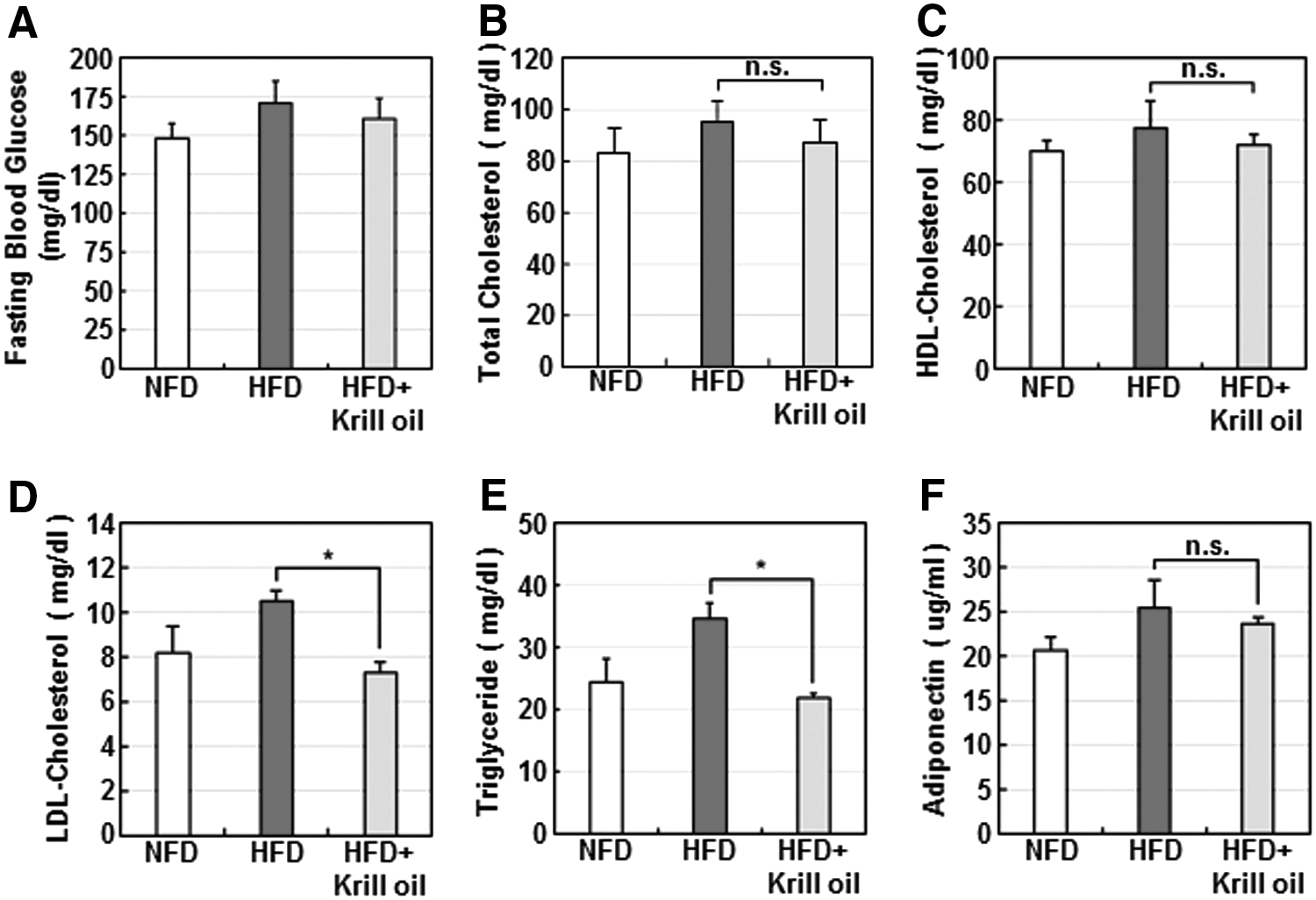
The effects of krill oil on blood glucose and lipid metabolism in mice. At the end of 10 weeks, fasting blood glucose
Krill oil inhibits fatty acid synthesis in Chang liver cells by activating AMPK
Next, to gain further insight into the molecular mechanisms underlying the inhibitory effects of krill oil on fat content, we examined the effects of krill oil on biochemical parameters in Chang liver cells. Cell viability profiles observed in the presence of the indicated concentrations of krill oil (25–100 μg/mL) for 24 h were similar to that of the vehicle-treated control, suggesting that krill oil did not exert significant cytotoxicity or affect the growth rate of Chang liver cells within the range of concentrations tested (Fig. 4A). To investigate the effects of krill oil on fatty acid synthesis, we assessed the expression of AMPK, ACC, sterol regulatory element-binding protein-1c (SREBP-1c), and FAS by immunoblot analysis. ACC and FAS play key roles in fatty acid synthesis, and SREBP-1c is a major transcription factor for lipogenic genes. Krill oil activated AMPK in a concentration-dependent manner, as evidenced by the increased levels of Thr172-phosphorylated AMPKα and Ser79-phosphorylated ACC, an intracellular substrate of AMPK 31 ; at a concentration of 100 μg/mL, krill oil induced a 2–4-fold activation of AMPK (Fig. 4B). The total amount of ACC, SREBP-1c, and FAS was also decreased by ∼50% compared with controls under these experimental conditions. In line with changes in protein levels, mRNA levels of ACC1, ACC2, SREBP-1c, and FAS were also significantly decreased by krill oil in a concentration-dependent manner, whereas the relative level of AMPKα mRNA was not changed.
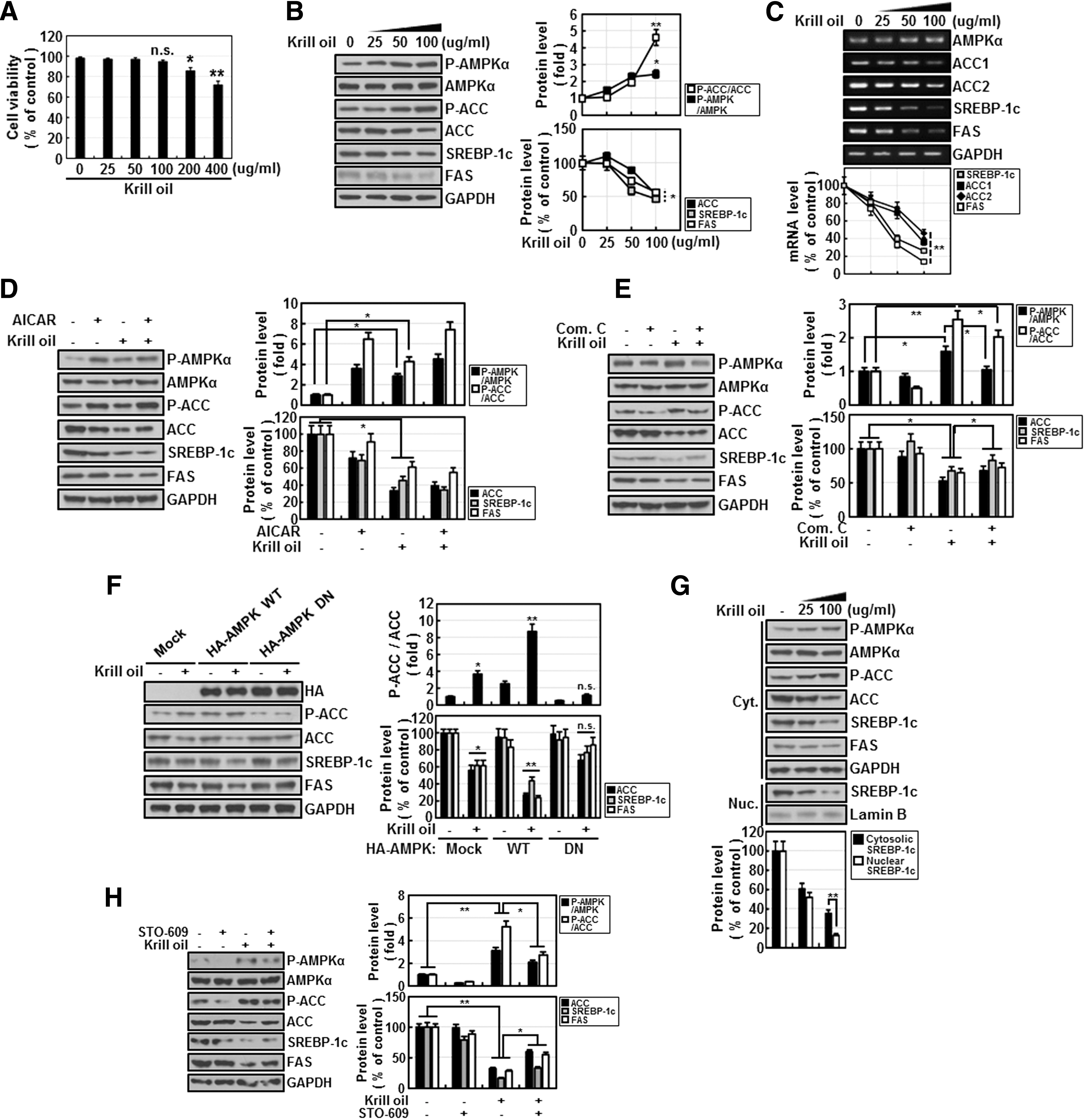
The effects of krill oil on AMPK signaling pathway in Chang liver cells.
A comparison of the effect of krill oil with that of AICAR, a well-known pharmacological activator of AMPK, showed that krill oil is as effective as AICAR in activating AMPK and suppressing its downstream targets, including ACC, SREBP-1c, and FAS (Fig. 4D). Next, to demonstrate the direct role of AMPK, we treated cells with krill oil in the presence of compound C, an AMPK inhibitor (Fig. 4E). Inhibition of krill oil-induced AMPK activation also slightly diminished the effects of krill oil on ACC, SREBP-1c, and FAS (Fig. 4E). Molecular approaches more clearly demonstrated the distinctive role of AMPK, showing that overexpression of wild-type (WT) AMPK synergistically enhanced the effect of krill oil, whereas overexpression of a dominant-negative (DN) form of AMPK effectively blocked the effect of krill oil (Fig. 4F). We also observed that krill oil treatment caused a more rapid decrease in nuclear SREBP-1c than cytosolic SREBP-1c (Fig. 4G). We further investigated the upstream of AMPK under krill oil treatment, and the inhibitor of CaMKKβ (Calmodulin-dependent protein kinase kinase), STO-609, significantly blocked AMPK activation by krill oil (Fig. 4H). Collectively, these results indicate that krill oil activates AMPK and, thereby, inhibits fatty acid synthesis, by downregulating genes involved in fatty acid synthesis.
Krill oil improves lipid metabolism in the livers of HFD-fed mice
Finally, we examined the profile of these biochemical parameters in the livers of mice in each diet group (Fig. 5). AMPK in the livers of HFD + krill oil mice was distinctively activated compared with that in HFD mice; moreover, krill oil effectively decreased protein (Fig. 5A) and mRNA (Fig. 5B) levels of ACC, SREBP-1C, and FAS in vivo. These results are consistent with in vitro data and suggest that krill oil inhibits fatty acid synthesis by suppressing SREBP-1c, ACC, and FAS in an AMPK-dependent manner.
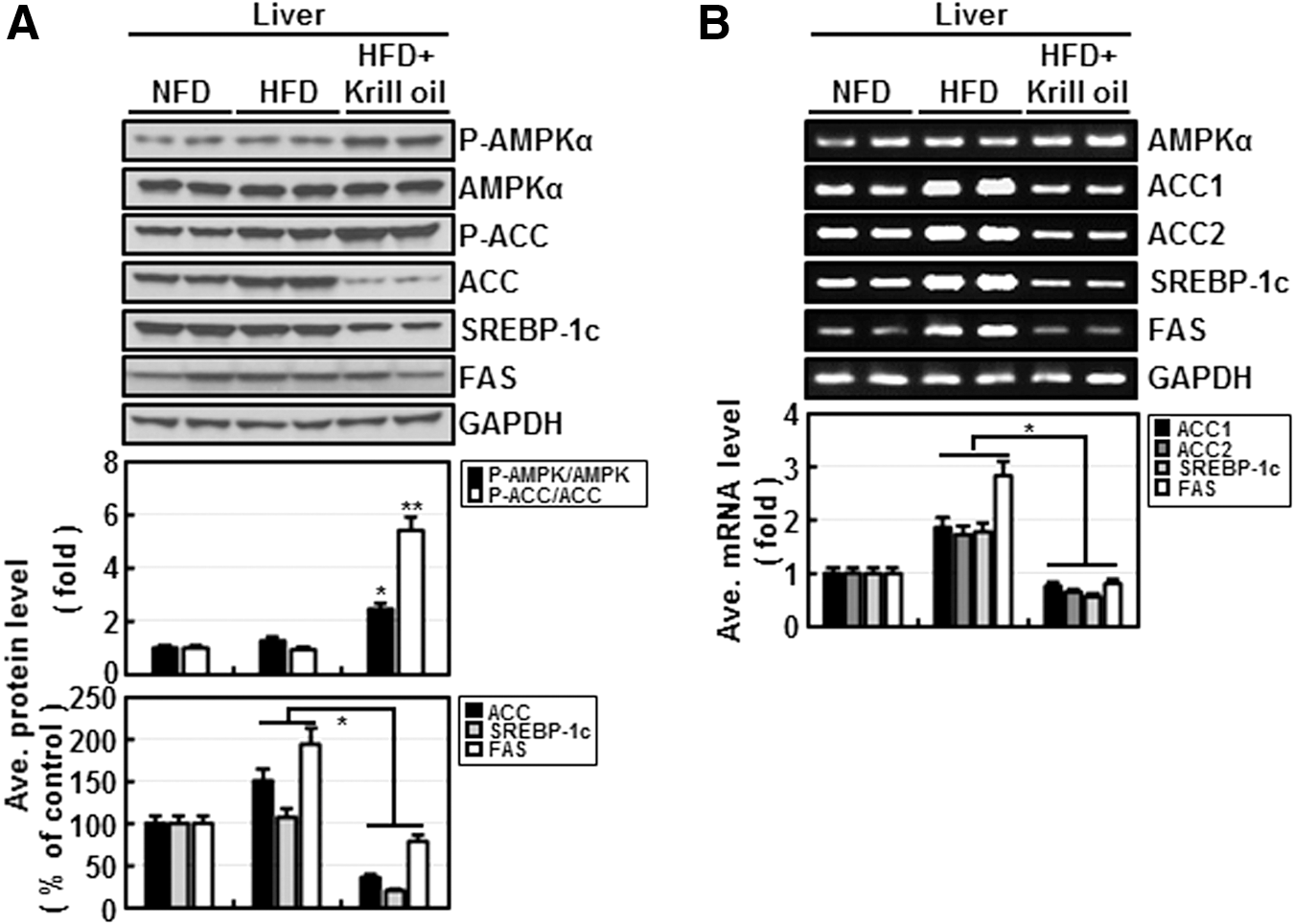
The effects of krill oil on mouse liver tissue levels of protein and mRNA levels of genes involved in fatty acid synthesis.
Discussion
Energy homeostasis is maintained through coordinated regulation of two parameters: energy intake and energy expenditure. Therefore, changes in either of these parameters can affect numerous physiological factors that contribute to the control of body weight. To examine the effect of krill oil on glucose and lipid metabolism, we initially allowed animals free access to ND or HFD without and with krill oil (2%, w/w) and then monitored food intake of each group for 1 week. Surprisingly, animals in the HFD + krill oil group consumed a significantly larger amount of food (3.8 g/day) than those in HFD or ND groups (2.5 g/day), presumably because the distinctive odor or taste of krill oil serves as an appetite stimulant. To eliminate these large differences in food intake among animal groups, which might interfere with data interpretation, we performed subsequent experiments under conditions of restricted intake in which the daily food supply was limited to 2.5 g, the average amount consumed by mice in ND and HFD groups. Under these conditions, krill oil exerted beneficial effects on several metabolic parameters in HFD-fed mice. Krill oil supplementation attenuated the body weight gain caused by HFD by ∼15% at the end of 10 weeks. These changes were accompanied by significantly lower visceral and subcutaneous fat content and suppression of hepatic steatosis. Krill oil also lowered the level of serum TGs and LDL-cholesterol.
Intriguingly, the liver showed the most dramatic response to krill oil supplementation among several examined tissues. Krill oil almost completely blocked the increase in liver mass, as well as hepatic steatosis, induced by HFD (Fig. 2). The liver is a central metabolic organ that regulates both circulating blood lipids and glucose levels, and our subsequent studies suggest that the beneficial effects of krill oil are associated with AMPK activation, showing that krill oil supplementation activated hepatic AMPK in vitro (Fig. 4), as well as in vivo (Fig. 5). AMPK has been recognized as a therapeutic target for treatment of obesity, type 2 diabetes, and cardiovascular disease because of its central role in the regulation of lipid and glucose metabolism. 26,27 AMPK inhibits fatty acid synthesis and cholesterol synthesis by directly phosphorylating and inhibiting ACC and HMG-CoA reductase, respectively. 22 In keeping with the activation of hepatic AMPK, the HFD-induced increase in the levels of circulating TGs and LDL-cholesterol was significantly decreased by krill oil supplementation (Fig. 3). Krill oil simultaneously suppressed the expression of genes involved in fatty acid synthesis, including ACC, FAS, as well as SREBP-1, a key lipogenic transcription factor (Figs. 4 and 5). This effect of krill oil appears to be dependent on AMPK activation, because pharmacological inhibition of AMPK or overexpression of DN-AMPK significantly attenuated the action of krill oil (Fig. 4E, F). These results are summarized schematically in Figure 6. Similar to our observations, metformin, a widely used drug for treatment of type 2 diabetes, activates hepatic AMPK and results in downregulation of hepatic lipogenic genes in vivo. 32

Schematic depiction of krill oil effects on AMPK and lipid synthesis signaling. On the basis of our results, we propose that krill oil, which is rich in DHA and EPA, decreases expression levels of SREBP-1c, FAS, and ACC by activating AMPK, leading to inhibition of fatty acid synthesis. DHA, docosahexaenoic acid; EPA, eicosapentaenoic acid; FAS, fatty acid synthase.
DHA and EPA, the major bioactive compounds in krill oil or fish oil, are known to regulate lipid and glucose metabolisms by acting as ligands for several transcription factors, including peroxisome proliferator-activated receptors (PPARs) 33 and sterol regulatory element-binding protein 1 (SREBP-1). 34 However, DHA is also known as an endogenous ligand of G protein-coupled receptor 120 (GRP120). 35 Moreover, a recent study demonstrated that DHA increases intracellular calcium (Ca2+) levels through GRP120, resulting in AMPK activation and glucose uptake in skeletal muscle. 36 AMPK is phosphorylated and activated by its upstream kinase, LKB1 (liver kinase B1), in an AMP-dependent manner 37 and is also phosphorylated and activated by Ca2+/calmodulin-dependent protein kinase kinase (CaMKK) in a Ca2+-dependent manner. 38 Therefore, krill oil may activate AMPK and its downstream targets through an increase in intracellular Ca2+. Indeed, our result (Fig. 4H) suggested that AMPK is activated by CaMKK under krill oil treatment. In conclusion, our study suggests that krill oil supplementation is a cost-effective and natural approach for improving dyslipidemia and, thereby, preventing metabolic disorders, including obesity. In particular, krill oil showed strong potential for preventing liver steatosis. We also demonstrated that AMPK mediates the effects of krill oil, opening up new avenues for mechanistic studies of krill oil effects.
Footnotes
Acknowledgments
This work was supported by the Food Functionality Evaluation Program funded by the Ministry of Agriculture, Food and Rural Affairs, with additional support from the Korea Food Research Institute and a grant (HI14C2700) from the Korean Health Technology R&D Project (AhR project).
Author Disclosure Statement
No competing financial interests exist.