
Abstract
In this study, we aimed to investigate the protective effect of three kinds of vegetable oils with different fatty acid compositions against cognitive impairment in an Alzheimer's disease (AD) mouse model. After intracerebroventricular injection of amyloid beta25–35 (Aβ 25–35) into the brain of institute of cancer research mice, olive oil (rich in oleic acid, C18:1), corn oil (rich in linoleic acid, C18:2), and perilla oil (rich in α-linolenic acid [ALA], C18:3) were administered at the oral dose of 500 mg/kg/day for 14 days. The results revealed that Aβ 25–35 induced learning and memory dysfunction according to the T-maze, novel object recognition, and Morris water maze tests. Among the three vegetable oils, however, the perilla oil group of mice showed marked attenuation of cognitive impairment, that is, a greater number of explorations on a new route/object than on an old route/object in the T-maze and novel object recognition tests. In the Morris water maze test, perilla oil decreased the time to reach the platform and increased the number of crossings over the target quadrant in which the platform was located previously. Furthermore, the beneficial effect of perilla oil supplementation on oxidative stress was reflected in the inhibition of malondialdehyde and nitric oxide (NO) production in Aβ 25–35-injected mice. We also found that perilla oil downregulated protein expression levels of inducible NO synthase and cyclooxygenase-2 and upregulated brain-derived neurotrophic factor. These findings showed that ALA-rich perilla oil has a potential for prevention or treatment of neurodegenerative diseases such as AD.
Introduction
A
Natural oils have unique properties depending on the fatty acid profile. Composition and double bonds of fatty acids depend on the kind of oil. The concentration of oleic acid (OA, C18:1) is high in olive oil compared with other seed oils. OA, which is a monounsaturated fatty acid, increases the plasma level of high-density lipoprotein, and this effect is associated with lower incidence of cardiovascular diseases. 3 Corn oil is commonly used as a source of linoleic acid (LA, 18:2n-6). Although there are various vegetable oils that are a good source of OA or LA, only a few vegetable oils such as walnut oil, flaxseed oil, and perilla oil contribute significant amounts of α-linolenic acid (ALA). 4 Among them, perilla oil derived from Perilla frutescens has the highest concentration of ALA: nearly 50–60%. 5 Many studies have shown beneficial effects of consumption of perilla oil: retinal protection, suppression of allergy, and prevention of coronary heart disease. 6,7 Moreover, ALA, a major fatty acid of perilla oil, reduces coronary disease risk and has an anti-inflammatory effect. 8,9 However, little attention has been given to cognitive improvement by vegetable oils with different fatty acid compositions. We hypothesized that fatty acid composition influences the protective effects against cognitive impairment induced by Aβ 25–35.
In this study, we tried to elucidate the effect of oils with different fatty acid compositions on learning and memory function. Olive oil (rich in OA), corn oil (rich in LA), or perilla oil (rich in ALA) was administered to AD mice and the learning and cognitive function were determined by behavioral tests. In addition, the protective mechanisms against AD were investigated.
Materials and Methods
Preparation of oils
Olive and corn oil were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Perilla oil that was used in this study was obtained by high-yield methods for purification of α-linolenic acid from P. frutescens var. japonica oil. 10
Instruments and reagents
Aβ 25–35 and malondialdehyde (MDA) were acquired from Sigma-Aldrich Co. Dimethyl sulfoxide and NaCl were purchased from Bio Basics, Inc. (Ontario, Canada). Thiobarbituric acid (TBA) was acquired from Lancaster Synthesis (Ward Hill, MA, USA). Phosphoric acid and 1-butanol were acquired from Samchun Pure Chemical Company (Pyeongtaek, Korea).
Animals and experimental protocols
Mouse strains showed different characteristics because of their unique genetic or physiological condition. The institute of cancer research (ICR) mouse, a strain of albino mice, was developed by Dr. Hauschka to create a fertile mouse line. The mice of this strain are commonly used for research in a wide range of fields, including neuroscience, cognitive function, immunology, and pharmacology because of fast growth rate, resistance to diseases, mimic natural pathologies, and relatively easy to breed. 11 Male ICR mice (5 weeks old; Orient, Inc., Seongnam, Korea) weighing 25–27 g were housed in plastic cages with free access to food and water and were maintained in a controlled environment (20°C ± 2°C, 50% ± 10% relative humidity, 12-h light/12-h dark cycle). The mice were subdivided into five groups comprising seven mice per group. The groups were defined as follows: Normal = 0.9% NaCl injection + oral administration of water; Control = Aβ 25–35 injection + oral administration of water; Olive oil = Aβ 25–35 injection + oral administration of olive oil (500 mg/kg/day); Corn oil = Aβ 25–35 injection + oral administration of corn oil (500 mg/kg/day); and Perilla oil = Aβ 25–35 injection + oral administration of perilla oil (500 mg/kg/day). We administered vegetable oils orally to mice by gastric gavage at a volume of 100 μL for 14 days. There were no significant differences in initial body weight among the groups. The timeline of behavioral procedures is presented in Figure 1.
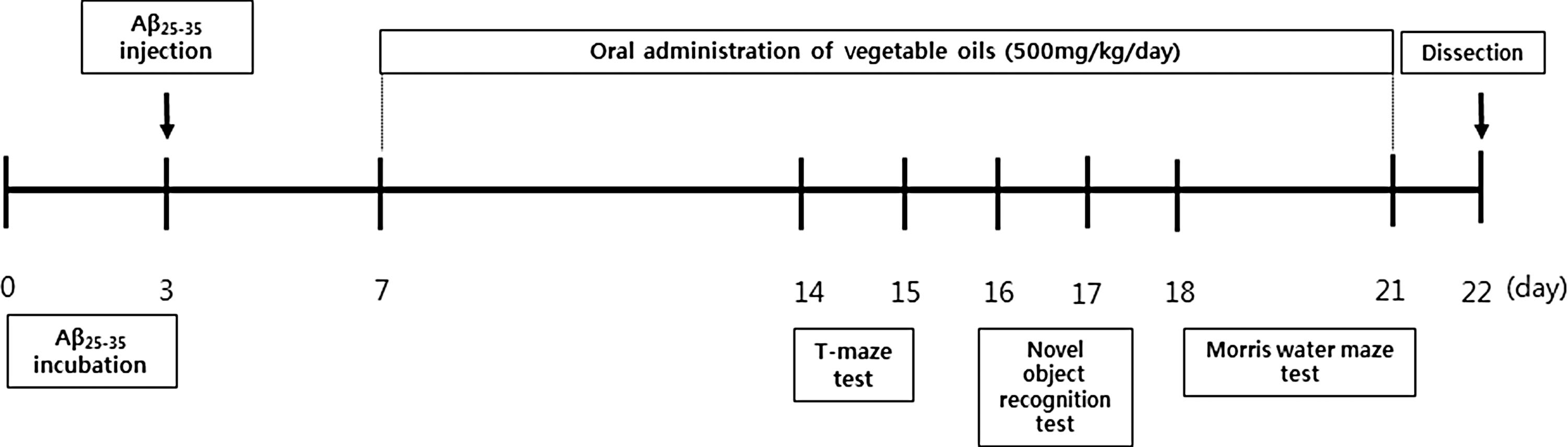
Behavioral experimental schedule for mice injected with Aβ 25–35.
The Aβ 25–35-infused mouse model
Aβ 25–35 was aggregated according to the procedure outlined by Maurice et al. 12 In brief, the peptide was dissolved and diluted in sterile distilled water to achieve a concentration of 1 mg/mL, aliquoted into tubes, and then dissolved. Aβ 25–35 was incubated at 37°C for 3 days before injection to induce aggregation. Distilled water containing aggregated Aβ 25–35 was injected into mice according to the procedure established by Laursen and Belknap. 13 The mice were lightly anesthetized with a Zoletil and Rompun mixture intraperitoneally, and the Aβ 25–35 solution was injected 0.8 mm posterior to the bregma and 1.5 mm lateral to the sagittal suture. All injections were made using a 10-μL Hamilton microsyringe fitted with a 26-gauge needle, which was inserted 2.2 mm beneath the surface of the brain. The animals were injected with 5 μL of sterile distilled water or 5 nmol of Aβ 25–35 aggregate into each cerebral lateral ventricle at a rate of 1 μL/min. The needle was left in the injection site for 1 min. The animal protocol used in this study has been reviewed by the Pusan National University–Institutional Animal Care and Use Committee (PUN-IACUC) on their ethical procedures and scientific acre, and it has been approved (Approval Number PNU-2015-0888).
The T-maze test
This test was conducted according to the procedure established by Montgomery. 14 The maze apparatus was T-shaped, and the walls were made of black boards (length of start and goal stems = 50 cm, width = 13 cm, height = 20 cm) that were glued to a square black board bottom. The maze consisted of a start box, left arm, and right arm with a door to separate the two sides. The mice were placed at the start box, and the number of touches and exploration of the right arm of the T-maze were recorded during a 10-min period (training session). The mice were then placed back into the same apparatus 24 h after the training session. They were allowed to explore the right and left sides of the maze freely for 10 min, and the numbers of touches and exploration were recorded (test session). The space perceptive ability (%) was calculated as the ratio of the number of left or right maze entries to the number of total maze entries multiplied by 100.
The novel object recognition test
This test was performed in a square black open-field apparatus (40 × 30 × 20 cm). 15 Two identical objects (plastic bottles) were placed at fixed distances within the square field. The mice were then placed at the center of the square field, and the number of touches of each object was recorded during a 10-min period (training session). The mice were placed back into the same field 24 h after the training session, but this time, one of the objects used during the training session was replaced with a new object (another plastic bottle). The mice were allowed to explore freely for 10 min, and the number of touches was recorded (test session). The object cognitive ability (%), a ratio of the amount of time spent exploring any one of the two original objects (training session) or the novel object (test session) to the total time spent exploring both objects, was used to measure cognitive function.
The Morris water maze test
This test was conducted according to the procedure established by Morris, with slight modifications. 16 The apparatus used in the Morris water maze test consisted of a dark plastic circular pool, 80 cm in diameter, surrounded by a 40-cm-high wall, and randomly divided into quadrants. White poster color was added to the pool water to make it opaque, and the water temperature was maintained at 22°C ± 1°C. Eight centimeter diameter platform was placed 1 cm below the water surface in the middle of one quadrant. The position of the platform was unchanged during the training session. Four posters on the walls of the apparatus provided visual cues for navigation. Three training trials per day were conducted for 3 days. During the training trials, the mice were placed randomly in the water facing the pool wall and allowed to swim for a maximum of 60 sec. The latency time required to find the platform was recorded. Mice that found the platform were allowed to rest there for 15 sec. If they failed to find the platform within 60 sec, they were allowed to stay on the platform for 15 sec to help them remember. A probe trial of the Morris water maze test was performed 1 day after the 3 days of training. In the primary test, the experiment was performed as before. In the secondary test, the trial was performed without the platform. The mice were placed in the pool and swam for 60 sec looking for the platform, and the latency time that the mice spent in the position previously occupied by the platform was recorded. In the tertiary test, the water was transparent and the number of times that it took the mouse to reach the platform, which was visible 1 cm above the surface of the water, was counted. Occupancy of the target quadrant (%) was calculated as the percentage of time spent in the target quadrant during a 60 sec trial.
Measurement of lipid peroxidation
MDA levels were measured by the method described by Ohkawa et al. 17 After completion of the behavioral testing, the mice were anesthetized by Zoletil and Rompun mixture intraperitoneally. Their brain, liver, and kidneys were excised immediately and placed on ice. The dissected tissue was homogenized with saline and mixed with a solution of 1% phosphoric acid and 0.67% TBA. After boiling for 45 min, the solution mixture was cooled in an ice bath, and 2 mL of 1-butanol was added followed by centrifugation at 3000 rpm (UNION 32R, Hanil Science Industrial Co., Ltd., Incheon, Korea) for 10 min. The absorbance values of the supernatant were measured at 535 and 520 nm. The level of lipid peroxidation was calculated using an MDA standard curve.
Nitric oxide scavenging activity
The nitric oxide (NO) concentration in tissues was determined by the method described by Schmidt et al.
18
One hundred fifty microliters of the supernatant from the lipid peroxidation procedure was mixed with 130 μL of distilled water, and 20 μL of the diluted solution was added to the same amount of phosphoric acid and 0.1%
Measurement of acetylcholinesterase enzymatic activity
This activity was measured using a modified method of Ellman et al. 19 Briefly, the brain sample was homogenized with 0.1 M phosphate buffer (PB, pH 7.4) and centrifuged at 13,000 rpm (Combi 514R, Hanil Science Industrial Co., Ltd., Incheon, Korea) at 4°C for 10 min. The resulting supernatant was used as an enzyme source for the acetylcholinesterase (AChE) assay; 400 μL of the supernatant, 4.1 mL of PB, 100 μL of 1 mM 5,5-dithiobis-(2-nitrobenzoic) acid, and 20 μL of 500 μM acetylthiocholine iodide were mixed. Then, the activity of the enzyme was measured using the spectrophotometer at a wavelength of 412 nm after 5 min of incubation at room temperature.
Western blotting
The murine brain was rapidly removed and homogenized with the lysis buffer containing a protease inhibitor cocktail. The homogenates were centrifuged at 13,000 rpm (Combi 514R, Hanil Science Industrial Co., Ltd., Incheon, Korea) for 30 min. After the supernatant was collected, the protein concentration was determined. Equal amounts of total protein were resolved by 10–13% SDS-PAGE gel. After electrophoresis, proteins were transferred to a PVDF membrane (Millipore, MA, USA). The membranes were incubated with 10% skim milk dissolved in phosphate-buffered saline-Tween® 20 (PBS-T) for 60 min and further incubated with a primary antibody in PBS-T overnight at 4°C. The primary antibodies were the following: anti-inducible nitric oxide synthase (iNOS, (1:200; Santa Cruz, CA, USA), anti-cyclooxygenase (COX)-2 (1:200; Santa Cruz), anti-brain-derived neurotrophic factor (BDNF, (1:200; Santa Cruz), and anti-β-actin (1:200; Santa Cruz). After that, the membranes were incubated with appropriate secondary antibodies (1:1000; Cell Signaling, Beverly, MA, USA) for 1 h. The immunocomplexes were visualized by pico-enhanced peroxidase detection (ELPIS-Biotech, Daejeon, Korea), and Western blot bands were assessed with a Davinci-Chemiluminescent imaging system (CoreBio, Seoul, Korea).
Statistical analysis
Results are expressed as mean ± SD. Statistical significance was determined using one-way analysis of variance, followed by Duncan's post hoc tests. Significance was set to P < .05.
Results
The T-maze test
Figure 2 shows the effects of vegetable oils on new route perception against Aβ 25–35-induced cognitive deficit. In contrast to the normal group, the Aβ 25–35-injected control group and olive oil- and corn oil-treated group did not show any significant changes between old and new route. However, perilla oil-treated mice showed a greater number of entries into a new route than the old route, indicating that perilla oil alleviated the cognitive deficit induced by Aβ 25–35.
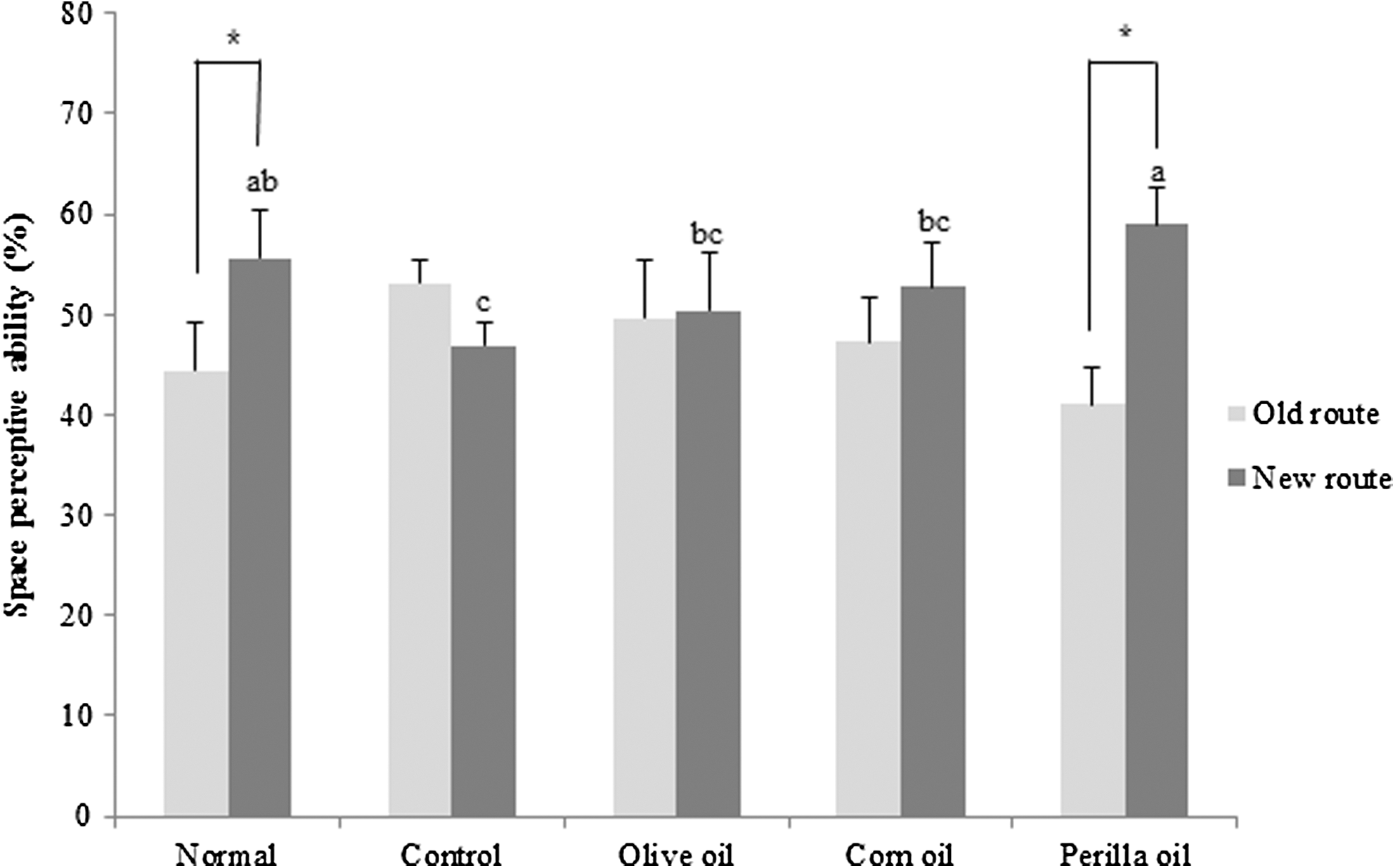
Effects of three kinds of vegetable oils on the spatial perceptive ability in the T-maze test. Data are presented as mean ± SD. *P < .05 compared with the old route. a–cMeans with different letters are significantly different (P < .05) from each other. Olive oil: Oral administration of olive oil (500 mg/kg/day), Corn oil: Oral administration of corn oil (500 mg/kg/day), Perilla oil: Oral administration of perilla oil (500 mg/kg/day).
The novel object recognition test
The effects of different kinds of vegetable oils on Aβ 25–35-induced memory impairment using the novel object recognition test are shown in Figure 3. After training, one of the familiar objects was replaced by a novel object. As a result, there was no significant variation in the number of touches between a familiar and novel object in the Aβ 25–35-injected control group. In contrast, the administered three kinds of vegetable oils (500 mg/kg/day) showed an increased number of touches on the novel object compared with the familiar object. Particularly, the perilla oil-treated group showed a relatively better recovery of recognition, indicating that perilla oil administration effectively improved the recognition ability of mice impaired by Aβ 25–35.
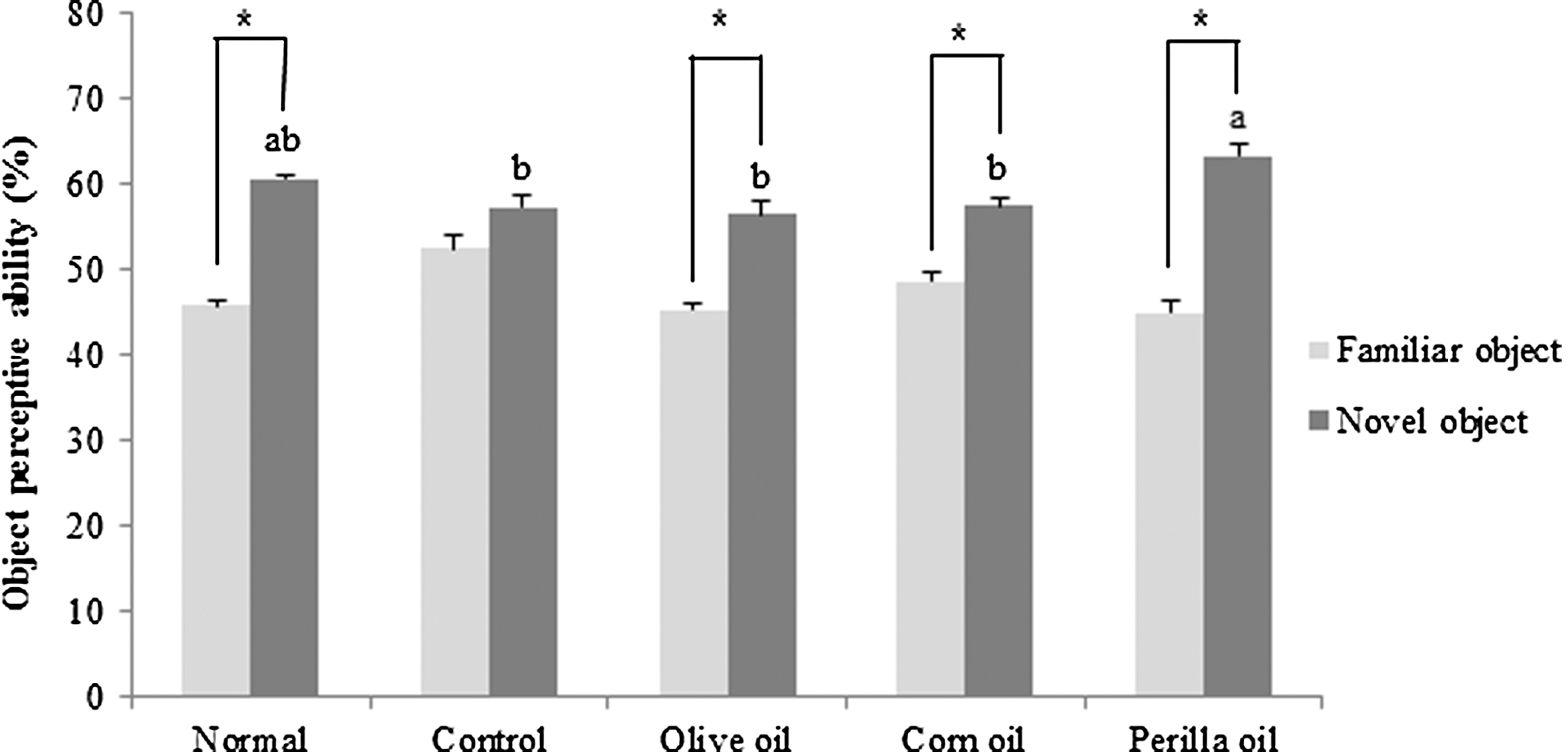
Effects of three kinds of vegetable oils on recognition memory in the novel object recognition test. Data represent mean ± SD. *P < .05 compared with the familiar object. a,bMeans with different letters are significantly different (P < .05) from each other. Olive oil: Oral administration of olive oil (500 mg/kg/day), Corn oil: Oral administration of corn oil (500 mg/kg/day), Perilla oil: Oral administration of perilla oil (500 mg/kg/day).
The Morris water maze test
In this test, we determined whether the Aβ 25–35-induced long-term memory deficit is improved by the administration of vegetable oils. Olive oil, corn oil, or perilla oil was orally administered to Aβ 25–35-injected mice for 14 days. The Aβ 25–35-injected mice were trained to reach the hidden platform for 3 days in a Morris water maze. On the final day, compared to the normal group, the Aβ 25–35-injected mouse group took a relatively longer time to reach the platform (Fig. 4). Although olive oil- and corn oil-treated groups did not show significant differences with the control group, the perilla oil group showed shorter time to reach the hidden platform. To further assess the spatial memory, the time to find the removed platform was measured. As shown in Figure 5, the percentage of total time spent in the target quadrant was 47.80% in the normal group compared to 25.00% in the control group. Olive and corn oil administrations did not have significant effects, whereas the perilla oil group showed a significantly increased time spent in the targeted quadrant: 33.88%. To exclude the influence of visual or exercise ability of mice, the escape latency to reach the visible platform was tested. The result indicated no statistically significant differences among the groups (Fig. 6). Overall, our results showed that perilla oil administration prevented long-term memory dysfunction in the AD mouse model.
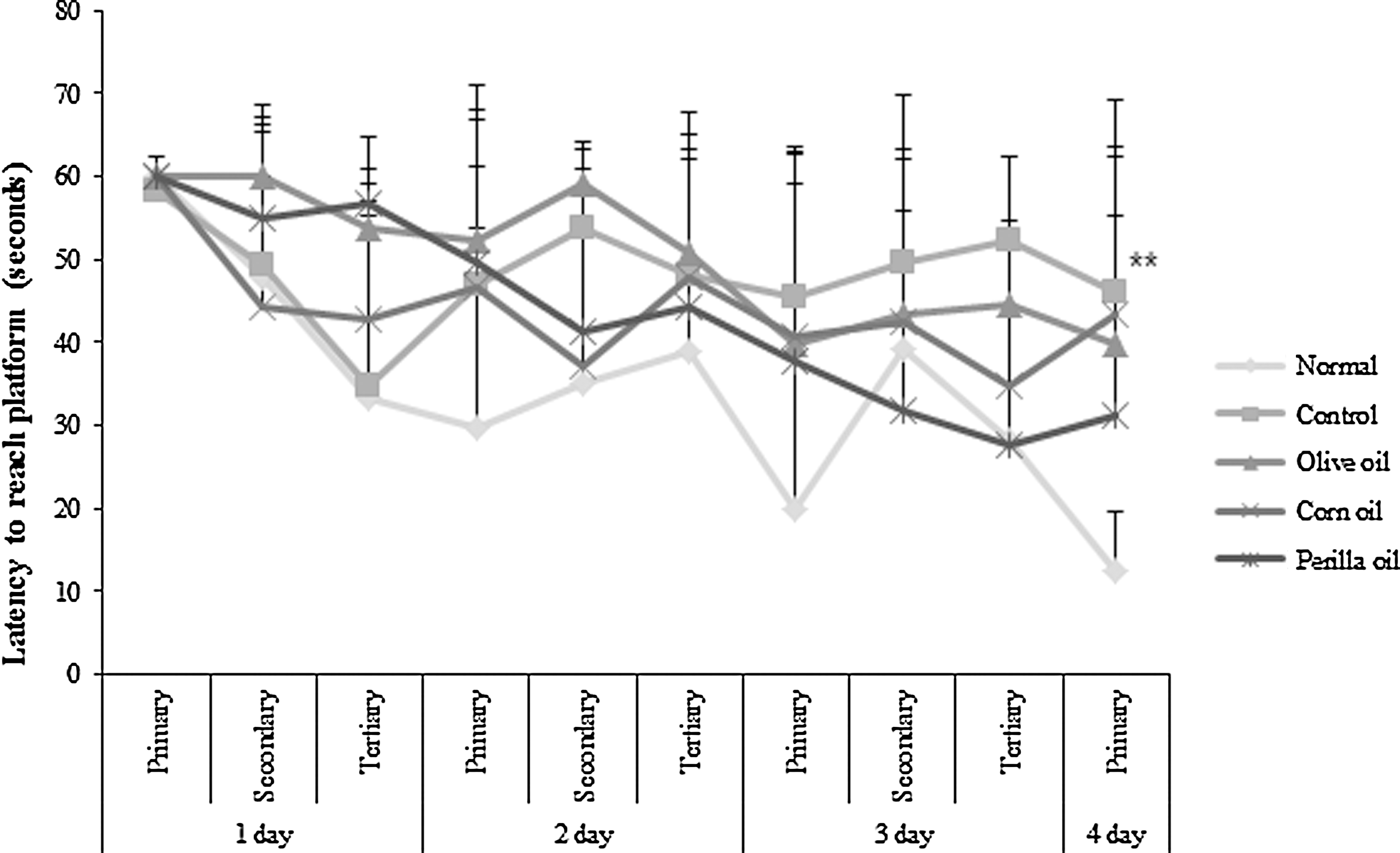
Effects of three kinds of vegetable oils on escape latency to the platform in the Morris water maze test. Data represent mean ± SD. **P < .01 compared with the normal group. Olive oil: Oral administration of olive oil (500 mg/kg/day), Corn oil: Oral administration of corn oil (500 mg/kg/day), Perilla oil: Oral administration of perilla oil (500 mg/kg/day).
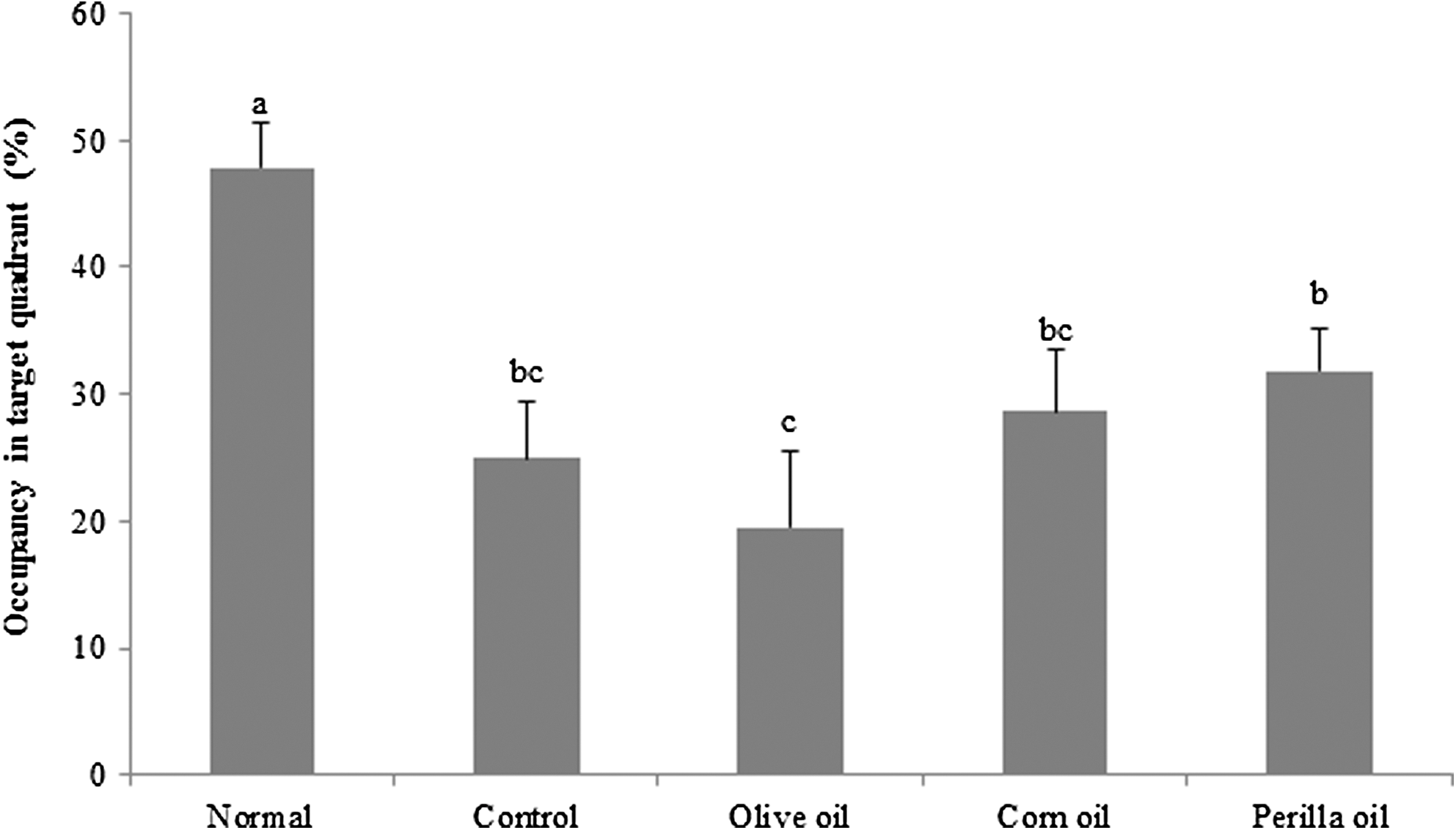
Effects of three kinds of vegetable oils on occupancy time of the target quadrant in the Morris water maze test. The percentage of time spent in the target quadrant was calculated in the water maze test on the final test day. Data represent mean ± SD. a–cMeans with different letters are significantly different (P < .05) from each other. Olive oil: Oral administration of olive oil (500 mg/kg/day), Corn oil: Oral administration of corn oil (500 mg/kg/day), Perilla oil: Oral administration of perilla oil (500 mg/kg/day).
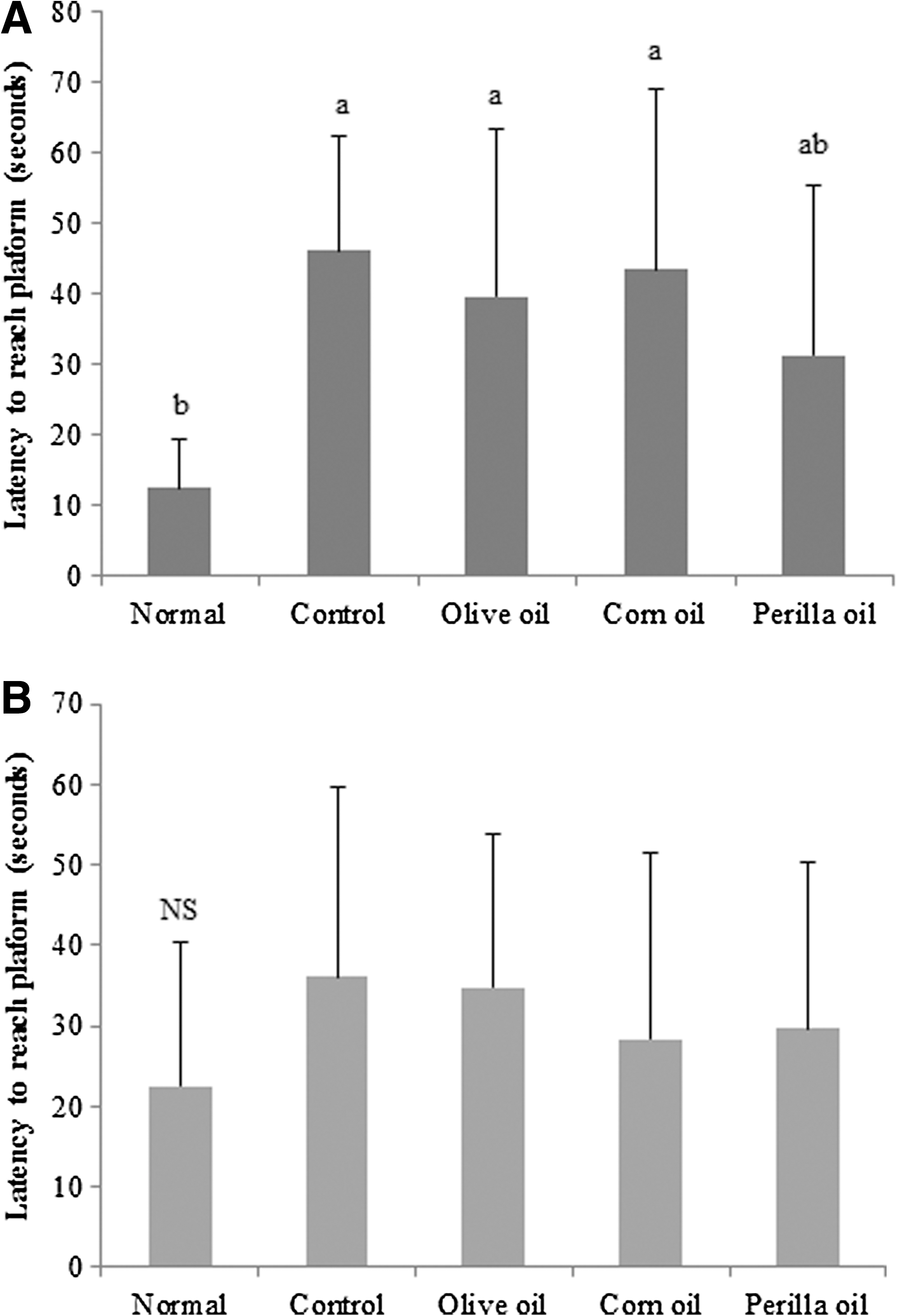
Effects of three kinds of vegetable oils on latency to reach a hidden platform
Measurement of lipid peroxidation
The inhibitory effects of vegetable oils on lipid peroxidation induced by Aβ 25–35 are shown in Table 1. Injection of Aβ 25–35 caused a significant increase in MDA levels in the brain (from 20.85 to 35.22 nmol/mg protein), while vegetable oil-administered groups showed attenuation of the overproduction of MDA. In particular, perilla oil markedly decreased the levels of MDA in the brain to 15.78 nmol/mg protein. MDA concentration in the kidney of Aβ 25–35-injected control group was 21.52 nmol/mg protein, which was twofold higher than in the normal group (10.48 nmol/mg protein). In contrast, the levels of MDA in the kidney significantly decreased in olive, corn, and perilla oil groups to values of 16.16, 17.47, and 14.18 nmol/mg protein, respectively. In addition, there was a significant decline in liver MDA concentrations after treatment with vegetable oils compared to the control group. The MDA values of the normal group and control group were 9.18 and 19.51 nmol/mg protein, respectively. In contrast, the olive and corn oil group showed a decreased MDA level: to 15.34 and 14.80 nmol/mg protein, and the perilla oil group showed the highest decline of MDA levels in the liver: 12.17 nmol/mg protein. These results revealed that supplementation with perilla oil effectively attenuated the lipid peroxidation induced by Aβ 25–35.
Values are presented as mean ± SD.
Different superscript letters indicate significant differences (P < .05) according to Duncan's multiple range test.
Olive oil: Oral administration of olive oil (500 mg/kg/day).
Corn oil: Oral administration of corn oil (500 mg/kg/day).
Perilla oil: Oral administration of perilla oil (500 mg/kg/day).
MDA, malondialdehyde.
Measurement of NO generation
The NO scavenging effects of vegetable oils in the brain, kidney, and liver are shown in Table 2. The levels of NO were significantly increased in the brain of the Aβ 25–35-injected mice compared to the noninjected mice group (8.92 nmol/mg protein). Nevertheless, the perilla oil group showed a decrease in NO concentration: from 13.25 nmol/mg protein in the control group to 11.08 nmol/mg protein. In the kidney, there were no significant changes after treatment with a vegetable oil compared to the control group. NO production in the liver of the Aβ 25–35-injected control group was 33.30 nmol/mg protein, while in olive oil-, corn oil-, and perilla oil-treated groups, it was lower at 26.01, 29.16, and 25.16 nmol/mg protein, respectively. These results showed that perilla oil decreased levels of NO in the brain and liver of mice and especially exerted the strongest protective activity against NO production in comparison with the other oils.
Values are presented as mean ± SD.
Different superscript letters indicate significant differences (P < .05) according to Duncan's multiple range test.
Olive oil: Oral administration of olive oil (500 mg/kg/day).
Corn oil: Oral administration of corn oil (500 mg/kg/day).
Perilla oil: Oral administration of perilla oil (500 mg/kg/day).
Measurement of AChE enzymatic activity
The effect of three kinds of vegetable oils on the AChE activity was assessed using brain tissues. As shown in Figure 7, the levels of AChE activity after injection of Aβ 25–35 were significantly increased compared to the non-Aβ 25–35-injected group. Nevertheless, administration of corn oil and perilla oil decreased the AChE activity, while the olive oil did not inhibit the enzymatic activity. In particular, the AChE activity was notably inhibited by administration of perilla oil, indicating that this oil contributes to the reduction of the ACh degradation rate.

Effects of three kinds of vegetable oils on acetylcholinesterase activity. Data represent mean ± SD. a–cMeans with different letters are significantly different (P < .05) from each other. Olive oil: Oral administration of olive oil (500 mg/kg/day), Corn oil: Oral administration of corn oil (500 mg/kg/day), Perilla oil: Oral administration of perilla oil (500 mg/kg/day).
Aβ 25–35-induced iNOS and COX-2 protein expression
We investigated the effect of three vegetable oils on protein expression of inflammatory mediators induced by Aβ 25–35. We found that the injection of Aβ 25–35 significantly upregulated iNOS and COX-2 compared to the normal group (Fig. 8), whereas administration of olive oil, corn oil, or perilla oil inhibited expressions of these inflammatory mediators. Although olive oil and corn oil inhibited COX-2 protein levels slightly, the perilla oil group attenuated Aβ 25–35-induced iNOS and COX-2 expression noticeably.
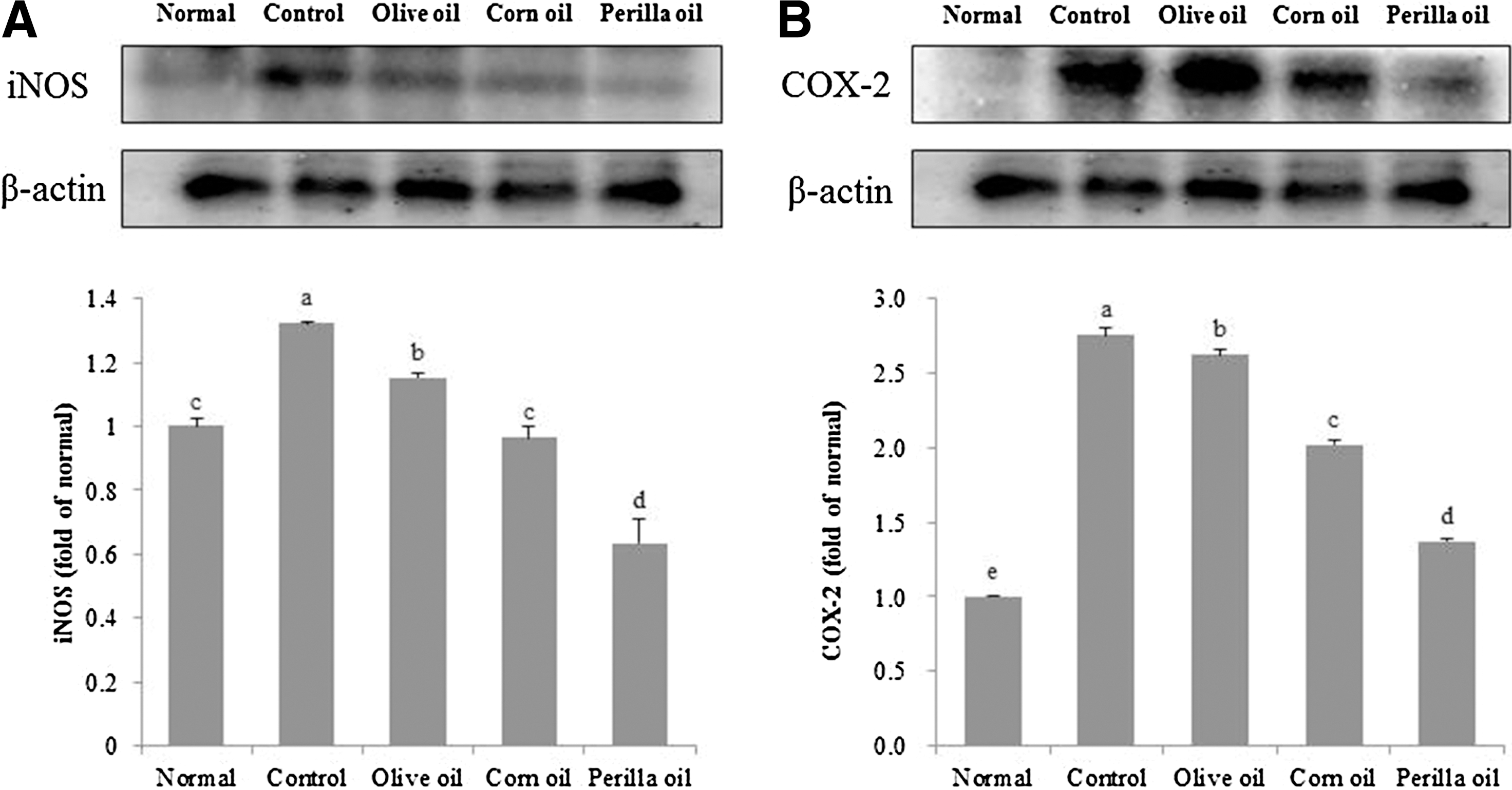
Effects of three kinds of vegetable oils on iNOS
Aβ 25–35-induced BDNF protein expression
We performed Western blotting to confirm the effect of three kinds of vegetable oils on BDNF protein expression. As shown in Figure 9, the Aβ 25–35-injected control group showed a decreased BDNF protein expression compared to the normal group. On the other hand, statistical analysis indicated that perilla oil increased the BDNF protein expression, whereas BDNF upregulation was not observed in the olive oil and corn oil groups.
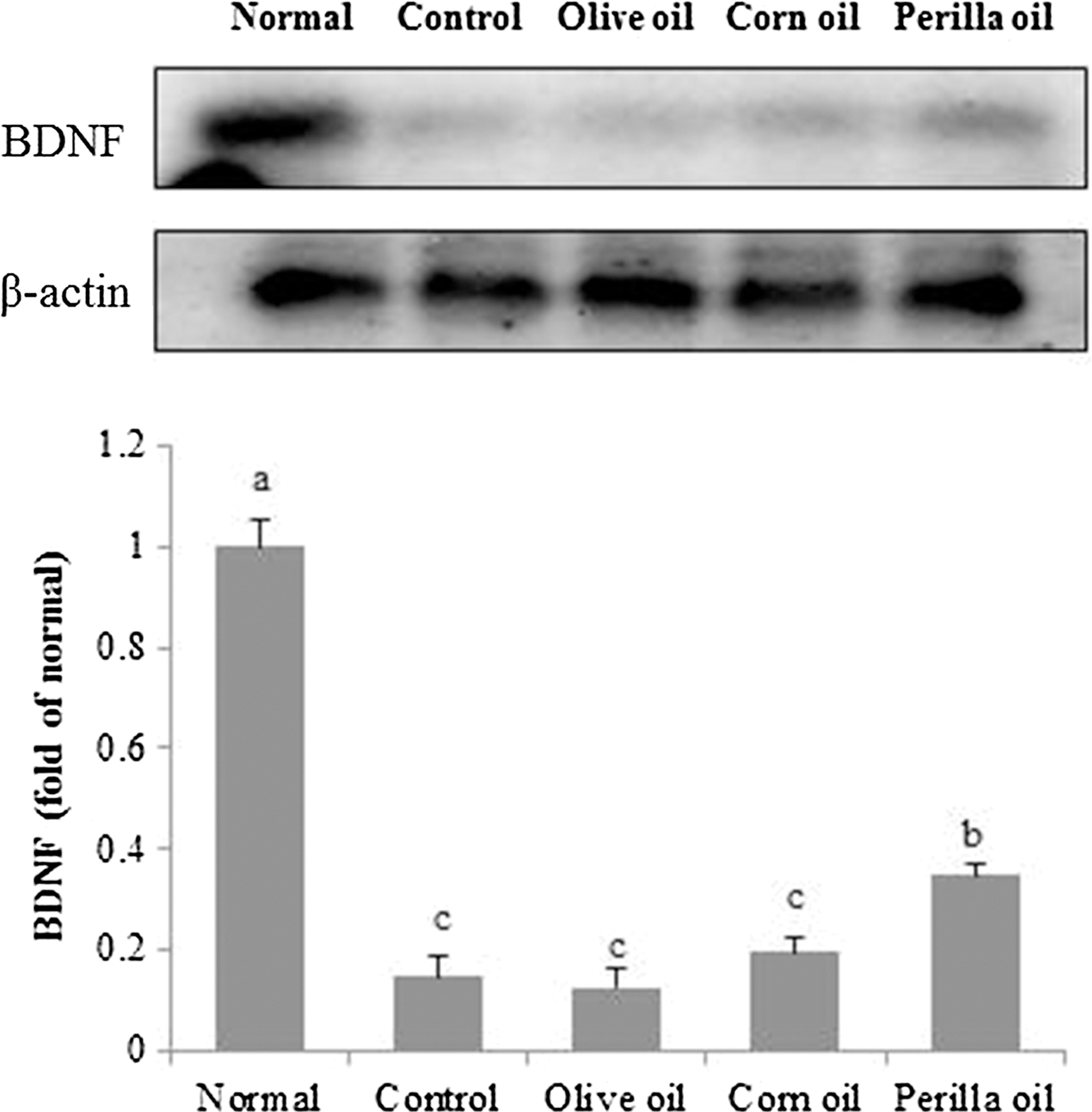
Effects of three kinds of vegetable oils on expression levels of brain-derived neurotrophic factor protein. Data represent mean ± SD. a–cMeans with different letters are significantly different (P < .05) from each other. Olive oil: Oral administration of olive oil (500 mg/kg/day), Corn oil: Oral administration of corn oil (500 mg/kg/day), Perilla oil: Oral administration of perilla oil (500 mg/kg/day).
Discussion
In this study, the effects of three kinds of vegetable oils (with different fatty acid compositions) on cognitive impairment were explored in Aβ 25–35-induced AD mouse model. Aβ deposition causes neuronal loss and cognitive decline, which is a pathological marker of AD. 20 Moreover, Aβ induces reactive oxygen species, reactive nitrogen species, and lipid and protein peroxidation, and performs a critical function in oxidative stress in the AD brain. 21 Although there are many recent studies elucidating the pathological processes of AD, the therapeutic strategy of targeting Aβ is still limited. We previously found that injection of Aβ 25–35 into the brain of mice results in learning and memory impairments, suggesting that accumulation of Aβ 25–35 contributes to neurotoxicity and brain aberrations. 22,23 Increasing attention has been focused on consumption of natural products in relation to AD research.
We tested whether the degree of unsaturation of 18-carbon fatty acids influences the neuroprotective effect of vegetable oil against the pathological changes caused by Aβ 25–35. We administered olive oil, corn oil, and perilla oil to Aβ 25–35-injected mice, and then the effects of the three kinds of vegetable oils (with different fatty acids composition) on Aβ 25–35-induced impairment of learning and memory were assessed by the T-maze, novel object recognition, and Morris water maze tests. Memory is divided into short-term (working memory) and long-term memory (reference memory). 24 These short-term and long-term memories are the results of differential neurobiological processes. When people or animals receive stimuli in certain environments, they memorize the experience through these two memory systems. The T-maze test, which is the most common version of the delayed alternation problem, was used to assess spatial working memory. According to Dudchenko, 25 the animal may remember their initial response to choose the alternative response, which is a short-term memory task. Test trial schedules vary in time after final training trials, varying from 30 min up to 24 h (the next day). 26 In T-maze or novel object recognition test, the mice showed an increased exploration of new route/object after training session, thus suggesting improvement of spatial working memory. 27,28 In this study, the effects of vegetable oils on space and object perceptive ability were examined in the T-maze test and novel object recognition test, respectively. Olive oil, corn oil, and perilla oil administrations resulted in higher rates of exploration of a novel object than a familiar object in the novel object recognition test, but olive oil- and corn oil-treated mouse groups did not show any change in perception between an old and new route in the T-maze test. In contrast, administration of perilla oil significantly increased the number of entries into a new route. These experiments revealed that perilla oil protected Aβ 25–35-induced memory dysfunction in mice.
Spatial memory is an essential adaptive behavior, which is required for survival. Previous evidence has demonstrated that spatial memory impairments were observed with hippocampus damage that the region of hippocampus plays a specific role in spatial information. 29,30 The injection of Aβ into mice brain causes hippocampal disruption, resulting in impairment of spatial memory. 31,32 These reports indicated that memory impairments in Aβ-injected mice were induced by hippocampus damage. Yoshida et al. 33 demonstrated that dietary perilla oils increased densities of synaptic vesicles in the hippocampus group, indicating that perilla oil led to improvement of learning performance. Spatial ability is involved in hidden platform of Morris water maze. The effect of vegetable oils on hippocampal-dependent long-term spatial memory in Aβ 25–35-injected mice was studied using the Morris water maze. After a training session for 3 days, we found that the escape latency of the perilla oil group was obviously shorter compared with the control group. Moreover, the injection of Aβ 25–35 caused a memory loss in the mice, with the consequence of reducing the number of crossings of the quadrant where the platform was located. Nevertheless, the perilla oil group showed an increase in the time spent in the target quadrant and the number of platform crossings during the test session. The time to reach the visible platform was not significantly different among all groups, indicating that this function is not associated with the exercise or visual ability. Taken together, our data suggest that the administration of perilla oil could protect from Aβ 25–35-induced deficits and may improve cognitive function in the AD mouse model.
There are extensive studies showing that the n-3 polyunsaturated fatty acids (PUFAs) have a wide range of effects on human health. The eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) are important for human brain health and, not surprisingly, are associated with prevention of neurodegenerative disorders, including depression and AD. 34,35 Nonetheless, EPA and DHA derived from certain fish oils are known to lead to food contamination such as methyl-mercury poisoning. 36 It was well established that ALA is a fatty acid for initiation of n-3 PUFA synthesis and is metabolized to EPA and DHA. 37 Therefore, perilla oil, a rich source of ALA, can be an alternative to fish oil supplements and has a neuroprotective effect similar to EPA and DHA.
The brain is a vulnerable target of oxidative damage because PUFAs are abundantly present in the brain. 38 Aβ peptide toxicity induces lipid peroxidation in the brain. 39 The key indicator of lipid peroxidation is the MDA level, which is generally higher in AD. We found significant elevation of MDA levels in the brains of mice injected with Aβ 25–35. On the other hand, MDA production was inhibited in the perilla oil group of mice compared to the Aβ 25–35-injected control group. These findings are in agreement with other studies showing that MDA content of the perilla oil group is remarkably decreased in comparison with the corn oil group in rats, 40 indicating that ALA from perilla oil, one of the n-3 PUFAs derived from plants, may be related to inhibitory action on lipid peroxidation.
Aβ can induce a proinflammatory response, including a release of cytokines and expression of iNOS. 41 Increasing expression of iNOS generates NO, which reacts with the superoxide radical and converts it to peroxynitrite. One of the possible pathological features of AD is the accumulation of NO resulting in oxidative damage. 42 According to Ii et al., 43 microglial cells are activated by Aβ to induce NO production that causes NO-mediated neuronal death. Furthermore, large quantities of NO can result in tissue damage or death. 44 Our results revealed that Aβ 25–35 significantly increased NO production in the brain and liver. On the other hand, oral administration of vegetable oils inhibited NO production in the liver. In particular, perilla oil significantly attenuated NO production in the brain.
Dietary supplementation with ALA-rich oil reduces production of inflammatory cytokines such as interleukin 1β or tumor necrosis factor α. 45 To determine whether these inhibitory effects on NO production were related to iNOS and COX-2 modulation, we examined protein expression by Western blotting. Our results showed that the expression levels of iNOS and COX-2 proteins are increased in the brains of Aβ 25–35-injected mice, as in prior studies. 46 On the other hand, considering the above result on NO production, the oral administration of perilla oil attenuated iNOS and COX-2 expression more significantly than the other vegetable oils did. These findings revealed that tissue damage induced by NO could be effectively prevented by treatment with perilla oil.
Lately, AChE inhibitors are considered promising therapeutic agents for AD. 47 The AChE activity is enhanced around Aβ in the AD brain. 48 Our data showed that the AChE activity in the brain was increased in the Aβ 25–35-injected mice group compared with the normal group. However, the AChE activity was significantly attenuated by administration of each of three kinds of vegetable oils. In particular, the perilla oil showed the strongest inhibitory effect on the AChE activity, resulting in increased ACh levels and contributing to reversal of the learning and memory deficits caused by Aβ 25–35.
BDNF plays an important role in neural development and maintenance of the central nervous system and mediates short- and long-term memory abilities. 49 Inhibition of BDNF protein expression induced by Aβ could be explained by a possible mechanism contributing to the development of a memory deficit in AD brain. 50 After Aβ 25–35 injection, we found that BDNF protein was significantly downregulated in the mouse brain. However, perilla oil administration was able to notably block the Aβ 25–35-induced decrease in BDNF levels. This result indicates that perilla oil administration may restore memory performance impaired by Aβ 25–35.
Several research groups have demonstrated bioavailability effects of dietary oils with different fatty acid compositions, where fatty acids have the same carbon length, but different double bonds. 51,52 The ALA-rich diet is positively associated with human health. Singer et al. demonstrated that a linseed oil-rich diet, which is rich in ALA, significantly depresses cholesterol levels in contrast to OA- or LA-rich diet. 53 In addition, compared with soybean oil, which is rich in OA and LA, perilla oil inhibits the development of mammary tumors in vivo. 54,55 Moreover, this PUFA has an anti-inflammatory effect induced by lipopolysaccharide, and ALA shows more beneficial effects than LA. 56 Therefore, consumption of ALA-rich perilla oil may be a promising functional food in relation to improvement of learning and memory abilities.
Our results showed that perilla oil improves learning and memory impairment as demonstrated by behavioral tests in the Aβ 25–35-injected mouse model. We also found that among the three kinds of vegetable oils, perilla oil significantly attenuated lipid peroxidation and NO production caused by Aβ 25–35 injection. This effect was mediated by suppression of iNOS and COX-2 protein expression levels. Furthermore, perilla oil has anti-AChE properties and enhances the memory ability by ameliorating Aβ 25–35-induced suppression of BDNF protein expression. In conclusion, perilla oil supplementation can alleviate cognitive impairment induced by Aβ 25–35, and this effect is related to the content of ALA.
Footnotes
Acknowledgments
This work was carried out with support of the “Cooperative Research Program for Agriculture Science & Technology Development (PJ010156032016),” Rural Development Administration, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.