
Abstract
Brown rice has nutritional benefits due to its abundant nutrients and high dietary fiber content. However, it is often consumed in a fermented form for improved chewy texture and digestibility. In this study, a polysaccharide, FBR-4a, was isolated from fermented brown rice and its chemical composition was analyzed. In addition, its immune-stimulating activity was determined using RAW264.7 cells. FBR-4a is a carbohydrate and consists of nine types of monosaccharides, including galactose, arabinose, rhamnose, mannose, fucose, xylose, glucose, galacturonic acid, and glucuronic acid. FBR-4a increased the gene expression of iNOS and IL-6 and enhanced the production of immune-stimulatory molecules (nitric oxide [NO], IL-6, and TNF-α) in RAW264.7 cells. Western blot analysis showed that FBR-4a phosphorylated the mitogen-activated protein kinases, NF-κB and c-Jun (major component of AP-1 transcription factor) in RAW264.7 cells, in a concentration-dependent manner. FBR-4a-induced IL-6 and NO production was completely suppressed by IκBα and JNK inhibition, but not by ERK and p38 inhibition. Furthermore, the phosphorylation of c-Jun was also suppressed by IκBα and JNK inhibition. These findings suggest that FBR-4a isolated from fermented brown rice has immune potentiation activity on macrophages through IκBα and JNK signaling pathways, which play a central role in IL-6 and NO production.
Introduction
R
Recently, plant-based polysaccharides have attracted increasing attention due to their potential biological activity, especially for their antioxidant 4 and immunomodulatory activities such as enhancement of cytotoxicity of natural killer cells, 5 proliferation of lymphocytes, 6 activation of macrophages, 7 and inhibition of cancer metastasis and angiogenesis. 8,9 A review by Schepetkin et al. 10 reported that botanical polysaccharides have relatively low toxicity, but are broad-spectrum immune modulators for host defense response. They suggested that the immune-stimulatory effect of polysaccharide occurs through macrophage stimulation and modulation of the complement system.
Macrophages play an important role in the innate immune response. Activated macrophages directly remove pathogens and foreign materials by phagocytosis or produce nitric oxide (NO) and reactive oxygen species. 11,12 The pathogens internalized by phagocytosis are degraded to peptide fragments, which are then displayed as antigen complexes with major histocompatibility complexes (MHCs) on the macrophage surface; the process is termed as antigen presentation. 13 These antigen and MHCs are recognized by T cells through T cell receptors and they promote T cell proliferation and differentiation. 14
Moreover, activated macrophages secrete cytokines, including IL-1, IL-6, and TNF-α, which activate adaptive immunity by promoting the differentiation of B cells to plasma cells. 15,16 The production of NO, IL-6, and the activation of surface molecules is initiated by cell surface receptors called pattern recognition receptors (PRRs). 17 Macrophages express PRRs such as Toll-like receptors, dectin-1, mannose receptor, complement receptor 3, which sense their ligands such as lipopolysaccharides, β-glucan from mushroom, mannan polysaccharide from yeast cell wall, and complement (C3b)-opsonized particles, respectively. 9 These receptors trigger intracellular signaling cascades such as mitogen-activated protein kinase (MAPK) and NF-κB pathways, which lead to the expression of immune-modulating substances in macrophages. 18
Brown rice has nutritional benefits due to its high content of dietary fiber; therefore, the intake of whole grains is recommended. However, brown rice is often consumed in fermented form for improved texture and digestibility. In this study, polysaccharides from fermented brown rice were isolated and purified. FBR-4a polysaccharide was obtained and evaluated to obtain polysaccharides with higher immune-stimulating activity. Furthermore, the immunomodulatory mechanism of FBR-4a was also identified.
Materials and Methods
Antibodies and reagents
Phospho-specific antibodies against JNK (Thr-183/Tyr-185), ERK (Thr-202/Tyr-204), p38 (Thr-180/Tyr-182), IκBα (Ser-32), and p65 (Ser-536), in addition to c-Jun (Ser-73) and antibodies against c-Jun (60A8), were purchased from Cell Signaling Technology (Danvers, MA, USA). Antibodies against JNK (FL), ERK1 (C-16), p38 (C-20), IκBα (C-21), p65 (C-20), and β-actin (I-19) were purchased from Santa Cruz Biotechnologies (Santa Cruz, CA, USA). Chemical inhibitors, SB 203580, JNK inhibitor II, PD 98059, and BAY 11-7082, were obtained from Calbiochem (Darmstadt, Germany). All chemical inhibitors were dissolved in dimethyl sulfoxide before use, and the final concentration was kept <0.1% for each experiment. Lipopolysaccharide (LPS) was obtained from Sigma-Aldrich (St. Louis, MO, USA). Dulbecco's modified Eagle's medium (DMEM) was purchased from Gibco (Life Technologies, New York, USA), and fetal bovine serum (FBS) was purchased from WELGENE (Daegu, Korea).
Extraction and purification of polysaccharide (FBR-4a) from fermented brown rice
Fermented brown rice powder was dissolved in water, followed by addition of three volumes of absolute ethanol to form precipitates. After centrifugation at 3490 g for 15 min, the precipitate was dissolved in distilled water and reprecipitated with four volumes of absolute ethanol. The precipitate was collected again by centrifugation (3490 g, 15 min), dialyzed for 2 days by using dialysis tubing (molecular weight >14,000), and lyophilized. Finally, the crude polysaccharide fraction (FBR-0) from fermented brown rice was obtained. FBR-0 was dissolved in distilled water and hydrolyzed by α-glucoamylase (10 U) and β-amylase (100 U) for 2 days (50°C, pH 5.5). Subsequently, the hydrolysate was boiled to inactivate the enzymes for 15 min, dialyzed, and lyophilized to obtain enzyme-treated crude polysaccharide (FBR-E). FBR-E was purified by ion-exchange chromatography on a DEAE-Sepharose FF (Cl− form) column (GE Healthcare, Uppsala, Sweden) with a stepwise gradient of NaCl (0, 0.05, 0.1, 0.2, and 0.3 M). Each fraction was collected, dialyzed against tap water, and freeze-dried. The active fraction, FBR-4, was eluted with 0.2 M NaCl and further purified by size-exclusion chromatography on a Sephadex G-100 column (GE Healthcare) equilibrated with 50 mM ammonium formate buffer (pH 5.5) at a flow rate of 1 mL/min at 4°C. After dialysis and lyophilization, subfraction, FBR-4a, was obtained.
Analysis of chemical properties of FBR-4a
The total neutral and acidic sugar contents of FBR-4a were determined by colorimetric method using the phenol–sulfuric acid reaction with galactose (Gal) as the standard 19 and the m-hydroxybiphenyl procedure with galacturonic acid (GalA) as the standard. 20 The protein content was determined using Bradford assay 21 with bovine serum albumin as a standard. Sugar composition of the polysaccharide sample was determined by gas chromatography analysis of their alditol acetates. 22 The sample was hydrolyzed with 2 M trifluoroacetic acid for 1.5 h at 121°C and converted into corresponding alditol acetates using the procedure of Zhao, Kiyohara, Yamada, Takemoto, and Kawamura. 23 The resulting alditol acetates were analyzed through GC system (ACME-6100; Young-Lin Co., Anyang, Korea) equipped with an SP-2380 capillary column (0.2 μm film thickness, 0.25 mm i.d. × 30 m; Supelco, Bellefonte, PA, USA). The carrier gas was N2, with a flow rate of 1.0 mL/min. The oven temperature was initially set at 60°C for 1 min, increased at 60–220°C (30°C/min), 220°C for 12 min, 220–250°C (8°C/min), and 250°C for 15 min. The molar percentage was calculated from the peak areas and response factors using the flame ionization detector.
Cell culture
RAW264.7 cells were obtained from the Korean Cell Line Bank (Seoul, Korea), and cultured at 37°C in DMEM supplemented with 10% heat-inactivated FBS and 1% antibiotic–antimycotic obtained from Life Technologies in a humidified atmosphere with 5% CO2 and 95% air.
Cytotoxicity assay
RAW264.7 cells were treated with FBR-4a polysaccharide at a series of concentrations (0.8–100 μg/mL) for 24 h, and then 10 μL of CCK-8 solution was added, and the cells were further incubated while cytotoxicity effects were evaluated. The percent cell viability was compared with that of the negative control, which corresponded to cells treated with only medium.
Determination of IL-6, TNF-α, and NO production
RAW264.7 cells were treated with FBR-4a polysaccharide at a series of concentrations (0.8–100 μg/mL) or with LPS (1 μg/mL) for 24 h. IL-6 ELISA kit from BD Biosciences (San Jose, CA, USA) and TNF-α ELISA kit from eBioscience (San Diego, CA, USA) were used according to the manufacturer's instructions. Nitrite oxide (NO) levels in culture media were measured using the Promega Griess Reagent System Kit, according to the manufacturer's instructions (Promega, Madison, WI, USA).
Preparation of cell lysates and immunoblotting
RAW264.7 cells were treated with indicated concentrations of FBR-4a for indicated time periods. Following treatment, the cells were washed with PBS twice, and then lysed in cold RIPA buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 0.25% deoxycholate, 1% NP-40, and 1 mM EDTA) containing 1 mM DTT, 1 mM PMSF, 1 mM sodium orthovanadate, 10 mM β-glycerophosphate, and a protease inhibitor cocktail (Roche Diagnostics Corp., Indianapolis, IN, USA). Cell lysates were resolved on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and subsequently transferred to an Immobilon-P PVDF membrane (Millipore, Billerica, MA, USA). The membrane was treated with 5% skim milk in Tris-buffered saline (TBS, pH 7.4) containing 0.1% Tween 20 (TBS-T) for 120 min and probed with specific primary antibodies overnight at 4°C. The membrane was then incubated with horseradish peroxidase-conjugated secondary antibody for 1 h and washed with TBS-T three times before visualization using an ECL system (Dogen, Korea).
Real-time reverse transcription–polymerase chain reaction
Total cellular RNA was isolated using the RNeasy mini kit (Qiagen, CA, USA). RNA was converted into cDNA by the RevertAid First-Strand cDNA Synthesis kit (Fermentas, MA, USA). Polymerase chain reaction (PCR) amplification was performed using the Power SYBR Green PCR Master Mix (Applied Biosystems, CA, USA) with sense and antisense primers. The primers for iNOS were 5′-GTTCTCAGCCCAACAATACAAGA-3′ and 5′-GTGGAC GGGTCGATGTCAC-3′; for IL-6, primers were 5′-GAGGATACCACTCCCAACAGACC-3′ and 5′-AAGTGCATCATCGTTGTTCATACA-3′. β-Actin and HPRT-1 were used as housekeeping genes, and for β-actin, the respective primers were 5′-GGCATTGTTACCAACTGGGACGAC-3′ and 5′-CCAGAGGCATACAGGGACAGCACAG-3′. For HPRT-1, the respective primers were 5′-GATTAGCGATGATGAACCAGGTT-3′ and 5′-CCTCCCATCTCCTTCATGACA-3′. Relative expression levels were determined by real-time reverse transcription-PCR (qRT-PCR) using the Step One Plus System (Applied Biosystems).
Statistical analysis
Statistical analysis was performed using Student's t-test, with **P < .01 or *P < .05 accepted as significant. Results are expressed as the mean ± standard deviation of triplicate experiments.
Results and Discussion
Chemical characterization of FBR-4a polysaccharide
FBR-4, a polysaccharide, was purified from fermented brown rice using DEAE Sepharose FF and Sephadex G-100 size exclusion column chromatography. To understand the chemical properties of FBR-4a, its chemical composition was analyzed. As shown in Table 1, FBR-4a comprised only carbohydrates, including 88.3% neutral sugar and 11.7% uronic acid; no proteins were detected. Next, the monosaccharide composition of FBR-4a was analyzed. As shown in Table 1 and Figure 1, FBR-4a consists of nine types of monosaccharides, including rhamnose, fucose, arabinose, xylose, mannose, galactose, glucose, GalA, and glucuronic acid. Relatively high content of galactose (43.2%) and arabinose (21.7%) suggested that FBR-4a has an arabinogalactan structure.
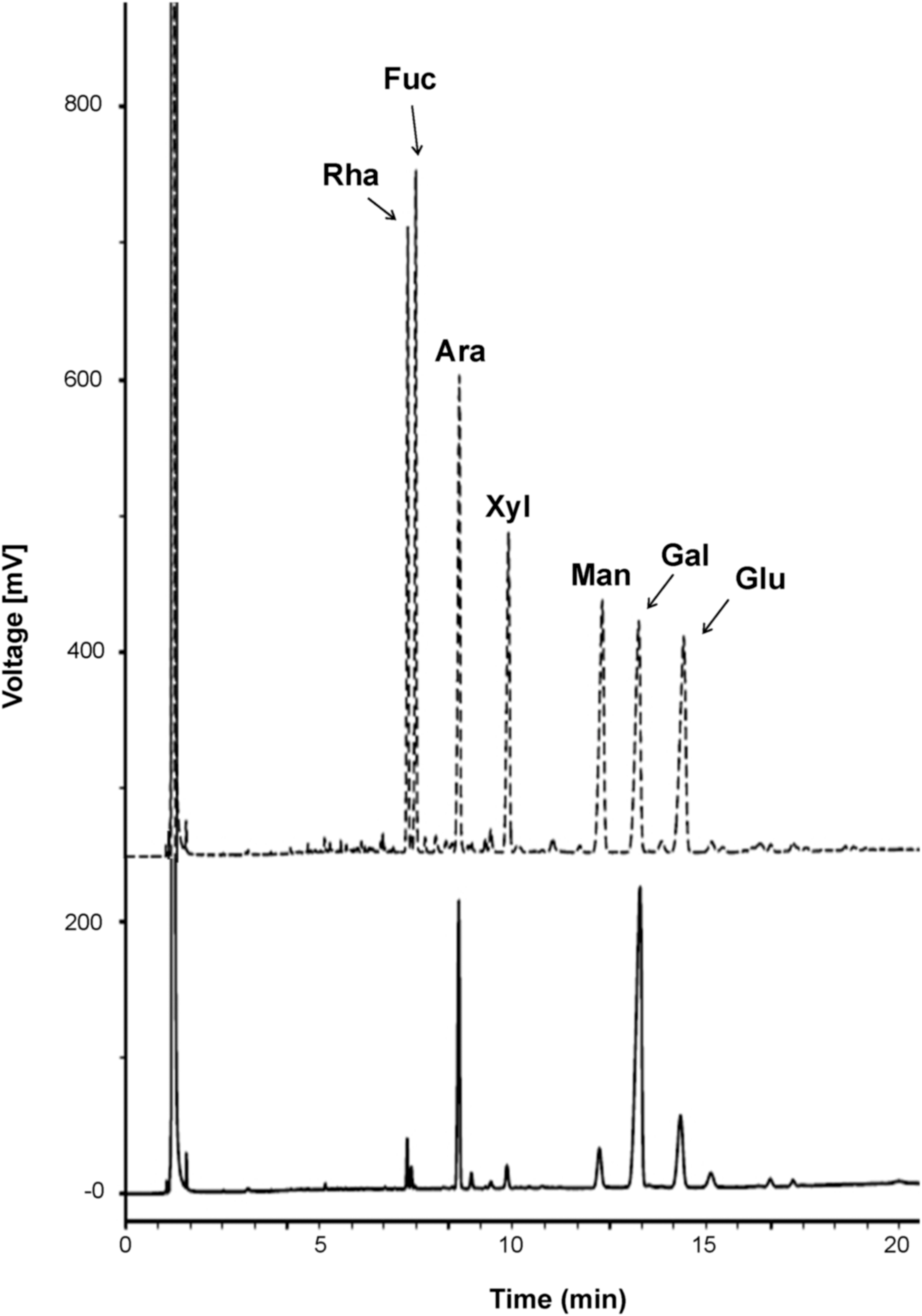
GC chromatography of mixed standards (dotted line) including rhamnose, fucose, arabinose, mannose, galactose, glucose and FBR-4a (full line) purified from fermented brown rice.
KDO means 2-keto-3-deoxy-D-manno-octulosonic acid.
Monosaccharides were analyzed using alditol acetates.
Mole % was calculated from the detected total carbohydrate.
Effects of FBR-4a on the viability of RAW264.7 cells
To investigate the role of the FBR-4a polysaccharide fraction in macrophage activation, first it was evaluated whether FBR-4a exhibits any cytotoxicity. RAW264.7 cells were treated with various concentrations of FBR-4a for 24 h and the cell viability was measured using the CCK-8-based MTT assay. FBR-4a showed cell proliferation activity at 0.8–100 μg/mL treatment for 24 h (Fig. 2), suggesting that the FBR-4a polysaccharide has no cytotoxicity in RAW264 cells.
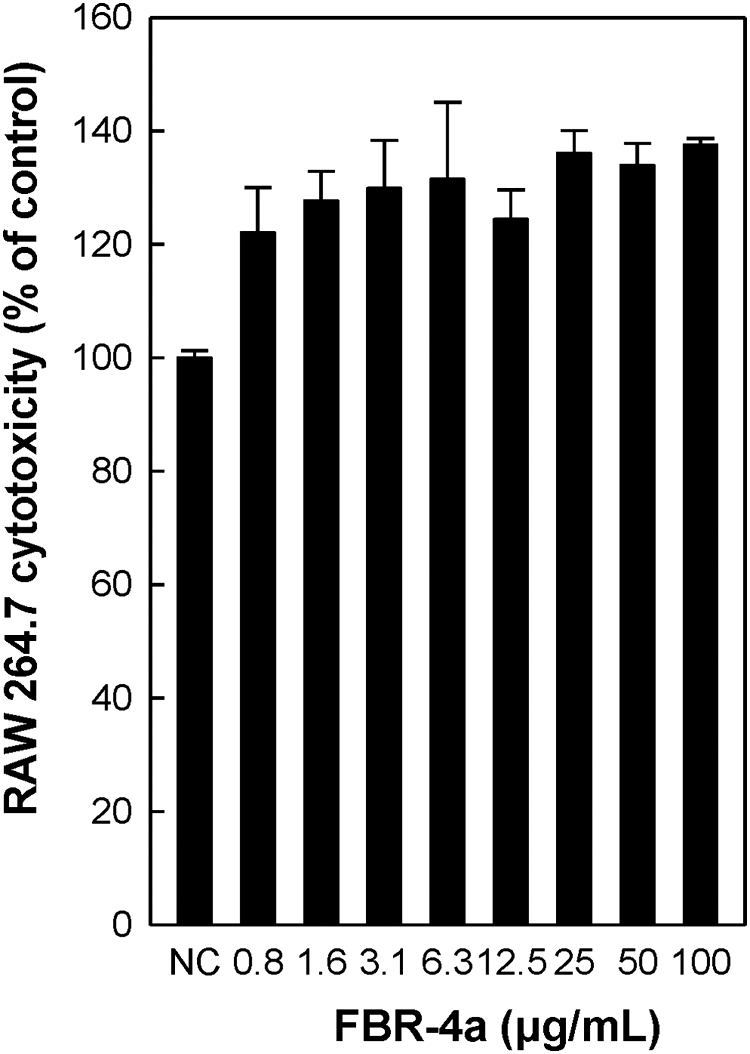
Cytotoxic effect of FBR-4a purified from fermented brown rice on RAW264.7 cells in vitro. RAW264.7 cells (1.0 × 105 cells/well, 96-well plates) were treated with FBR-4a polysaccharide at a series of concentrations (0.8–100 μg/mL) for 24 h, and cytotoxicity was determined using the CCK-8-based colorimetric assay. Not treated FBR-4a was used as the NC. NC, negative control.
Effects of FBR-4a on iNOS and IL-6 gene expression in RAW264.7 cells
Macrophage activation is the first line of host defense against pathogens or foreign materials. Activated macrophages produce NO to kill pathogens and release various cytokines such as IL-6, IL-12, and TNF-α to activate adaptive immunity. 24 Therefore, the gene expression of iNOS and IL-6 was investigated. The expression levels of both iNOS and IL-6 messenger RNA (mRNA) increased after FBR-4a treatment for 6 h in RAW264.7 cells (Fig. 3). The iNOS mRNA expression was markedly enhanced by 1 μg/mL FBR-4a treatment; 25 μg/mL treatment showed a similar level compared with LPS treatment. Similarly, FBR-4a also enhanced IL-6 mRNA expression in a concentration-dependent manner in RAW264.7 cells.
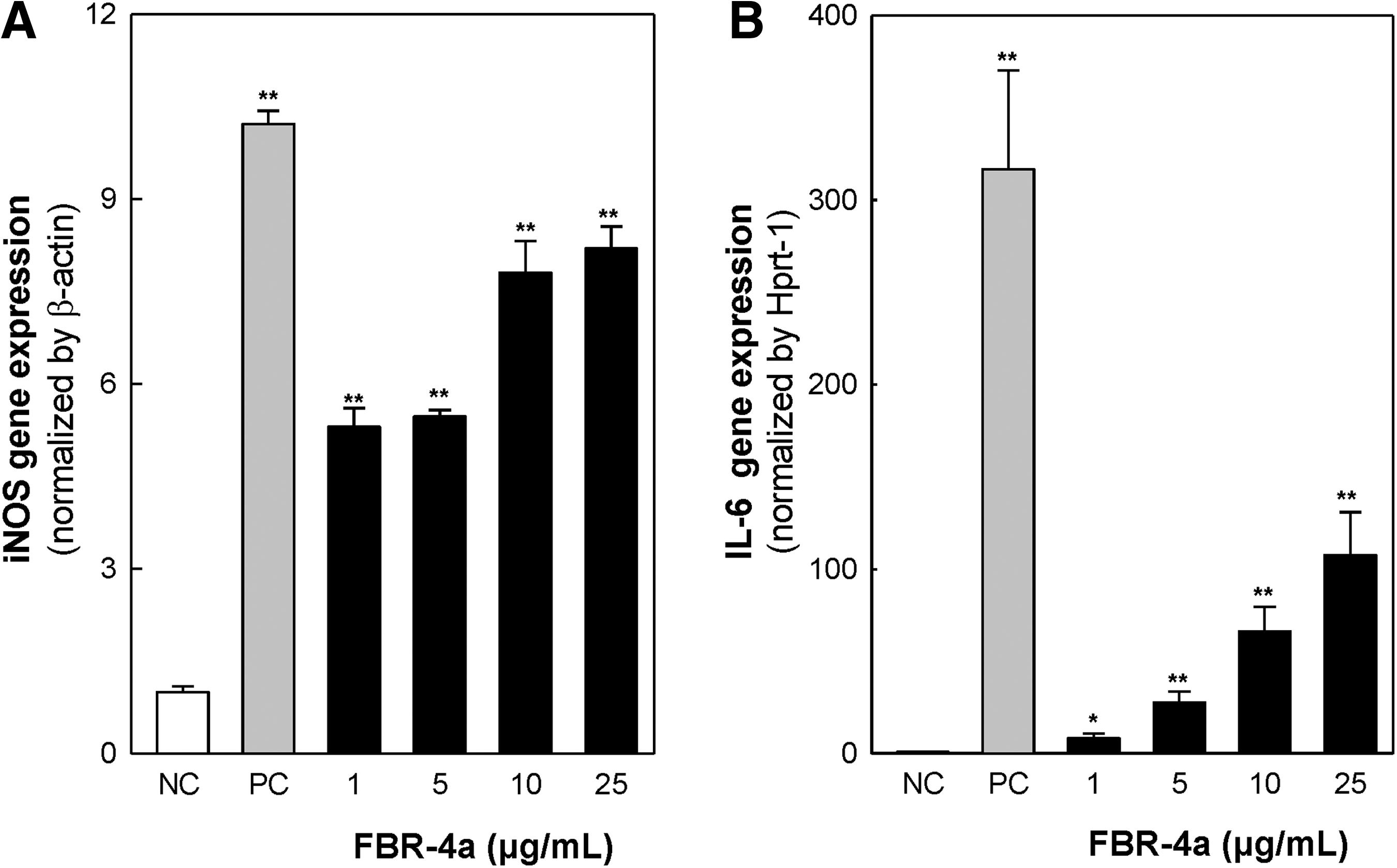
Effect of FBR-4a on mRNA expression of iNOS
Effects of FBR-4a on NO, IL-6, and TNF-α production
NO produced by macrophages of the immune system acts as a chemical barrier against bacteria and cancer cells. After activation, macrophages secrete cytokines such as IL-6 and TNF-α, which induce adaptive immune responses. To investigate, whether FBR-4a produces NO and cytokines, including IL-6 and TNF-α, RAW264.7 cells were treated with various concentrations of FBR-4a. As shown in Figure 4A, NO production was significantly increased with increase in concentration of FBR-4a from 0.8 to 100 μg/mL; the maximal NO production was observed at 50 μg/mL FBR-4a treatment. Similarly, IL-6 production increased with increasing FBR-4a concentration, dramatically increasing at 100 μg/mL treatment. Furthermore, the TNF-α production also increased FBR-4a concentration dependently. These data suggest that FBR-4a possesses macrophage-stimulating activity.
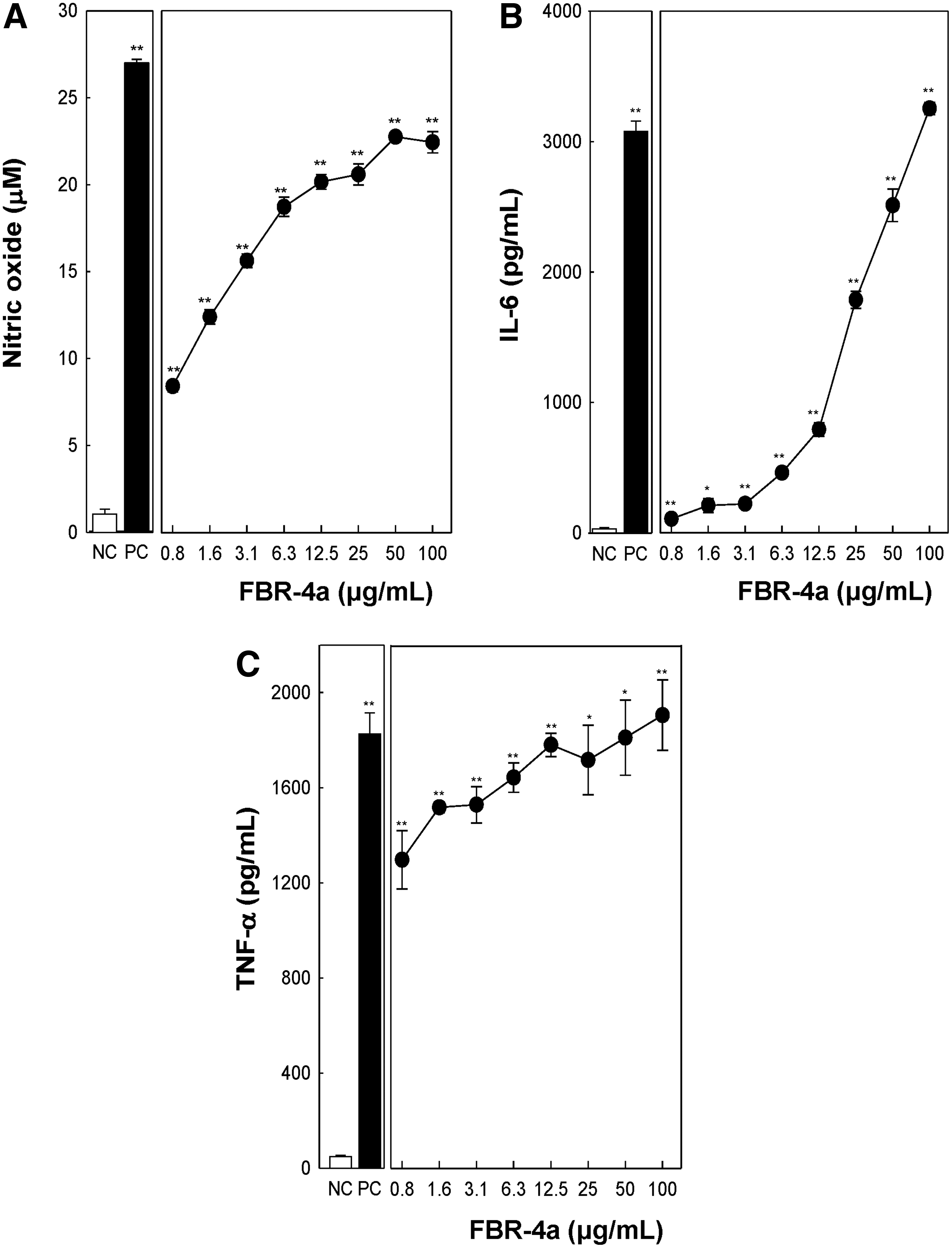
Effect of FBR-4a on nitric oxide
Effects of FBR-4a on MAPK phosphorylation and NF-κB activation in RAW264.7 cells
Recently, many studies have reported that botanical polysaccharides activate macrophages through their intracellular signaling pathways. MAPK and NF-κB are important proteins that regulate the gene expression of iNOS, IL-6, and TNF-α in RAW264.7 cells. 25 RAW264.7 cells were treated with various concentrations of FBR-4a or LPS (as a positive control) for 30 min to investigate whether FBR-4a activates MAPK and NF-κB pathways. FBR-4a significantly phosphorylated MAPKs (JNK, ERK, and p38) in a concentration-dependent manner (Fig. 5); however, total MAPKs were not affected. The phosphorylation of c-Jun, one of the AP-1 transcription factor family members, was also enhanced by FBR-4a treatment. Transcription factor c-Jun activation is important for gene expression and is mainly regulated by MAPKs such as JNK, ERK, and p38. 26,27 Next, NF-κB activation by FBR-4a was investigated in RAW264.7 cells. When the cells are in an unstimulated condition, NF-κB exists in cytosol with its inhibitory protein, IκB. After stimulation, IκB is phosphorylated by IKKs and is subsequently degraded in the proteasome, leading to phosphorylation and translocation of the p65 subunit of NF-κB into the nucleus. Therefore, this study evaluated the effect of FBR-4a on degradation of IκBα and phosphorylation of p65. 25,28 Treatment with FBR-4a induced IκBα degradation and increased phosphorylation of p65 in a concentration-dependent manner (Fig. 5). It suggests that FBR-4a activates MAPK and NF-κB signaling pathways are implicated in transcriptional activation in RAW264.7 cells.

Phosphorylation of MAPK, c-Jun, and NF-κB pathways by treatment of FBR-4a from RAW264.7 cells. RAW264.7 cells (3.0 × 106 cells/6-cm dish) were treated with FBR-4a polysaccharide at a series of concentrations (0–25 μg/mL) or LPS (1 μg/mL) for 30 min. The level of β-actin was measured as an internal loading control. MAPK, mitogen-activated protein kinase.
Effects of FBR-4a on IL-6 and NO production in RAW264.7 cells
To identify the underlying signaling pathways by which FBR-4a produces IL-6 and NO, cells were treated with chemical inhibitors specific to MAPKs and IκBα. As shown in Figure 6A, treatment with BAY11-7082 (BAY), a specific inhibitor for IκBα phosphorylation, completely inhibited FBR-4a-induced IL-6 production. Similarly, JNK inhibitor II, a specific inhibitor for JNK phosphorylation, also significantly inhibited FBR-4a-induced IL-6 production. In contrast, FBR-4a-induced IL-6 production was not inhibited when the cells were treated with SB203580 (SB) and PD98059 (PD), specific inhibitors of p38 and ERK, respectively (Fig. 6A). Similar results were obtained for FBR-4a-induced NO production. NO production was significantly inhibited by treatment with BAY and slightly inhibited by treatment with JNK inhibitor II (Fig. 6B). However, treatment with SB and PD did not affect NO production. These data suggested that the NF-κB pathway and JNK have important roles in FBR-4a-induced IL-6 and NO production in RAW264.7 cells.
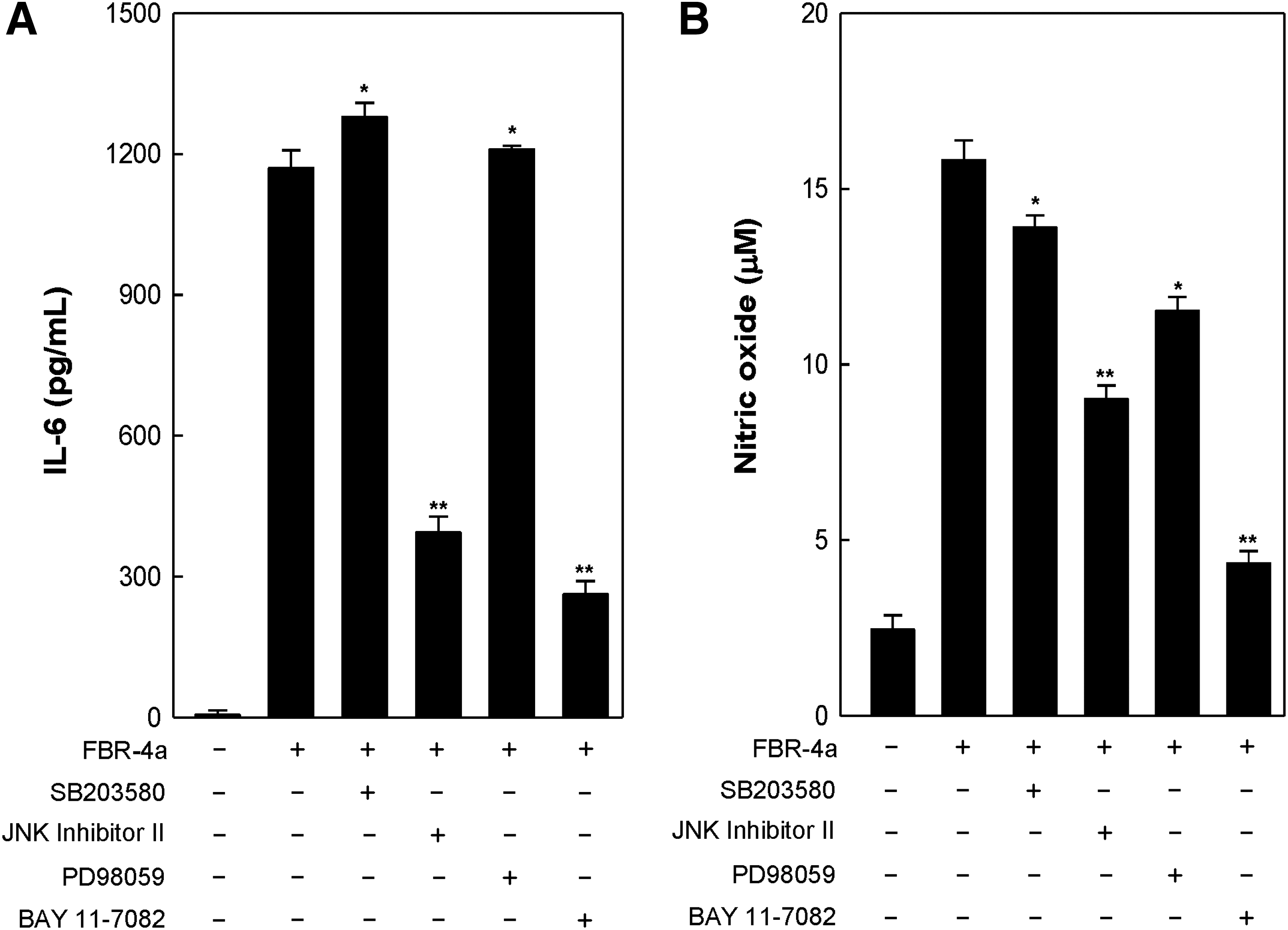
Effect of FBR-4a on IL-6
Effects of FBR-4a-induced JNK and NF-κB activation on c-Jun phosphorylation in RAW264.7 cells
RAW264.7 cells were treated with chemical inhibitors to identify the role of c-Jun activation in FBR-4a treatment. Chemical inhibitors against signal molecules suppressed phosphorylation of each molecule (Fig. 7A, B). Interestingly, FBR-4a-induced c-Jun phosphorylation was blocked by BAY and JNK inhibitor II treatment (Fig. 7A). However, when the cells are treated with PD and SB, FBR-4a-induced c-Jun phosphorylation was not inhibited. Collectively, these data suggest that c-Jun transcription factor regulated by NF-κB and JNK has a role in FBR-4a-induced IL-6 and NO production in RAW264.7 cells.
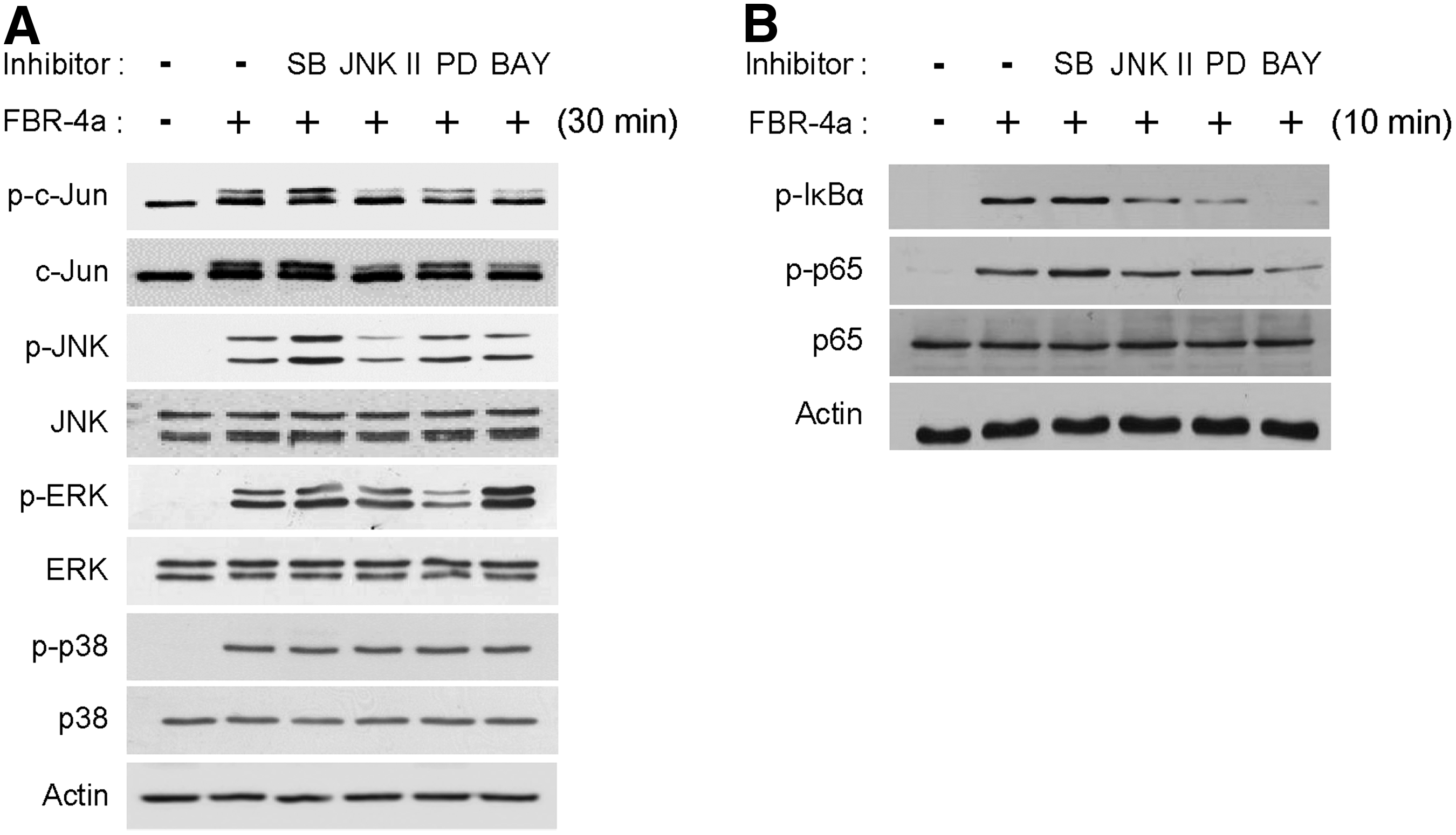
Effect of FBR-4a on MAPK, c-Jun phosphorylation
Summary
Recently, polysaccharides isolated from botanical sources have been shown to possess beneficial effects through modulation of macrophage immune functions. 29 Two things need to be investigated for these bioactive polysaccharides. First is analysis of chemical properties of polysaccharides, such as sugar composition, molecular weight, and types of glycosidic bonds. Second is elucidation of underlying mechanisms by which the polysaccharide induces macrophage activation.
This study investigated the chemical properties of FBR-4a and identified the macrophage activation pathways using chemical inhibitors. FBR-4a consisted of nine types of monosaccharides, mainly galactose (43.2%) and arabinose (21.7%), which suggested that FBR-4a possessed arabinogalactan structure. FBR-4a strongly induced iNOS and IL-6 mRNA expression and induced the production of NO, IL-6, and TNF-α in RAW264.7 cells. It was found that activation of signaling proteins, NF-κB and JNK, is required for FBR-4a-induced production of immunostimulatory molecules (IL-6 and NO). In conclusion, FBR-4a arabinogalactan purified from fermented brown rice has strong macrophage activation ability through NF-κB and JNK signaling pathways.
Footnotes
Acknowledgment
This work was supported by Kyonggi University's Graduate Research Assistantship 2016.
Author Disclosure Statement
No competing financial interests exist.