
Abstract
Escalating evidence indicates that ginseng treatment protects against psychotoxic behaviors and memory impairment. Although the underlying mechanism of schizophrenia remains elusive, recent investigations proposed that downregulation of glutathione (GSH) can be involved in the pathogenesis of this disorder. Since little is known about the effects of ginseng in a schizophrenia-like animal model, we selected mountain-cultivated ginseng (MG) from a variety of ginseng extracts to investigate the effect of ginseng on the psychosis induced by phencyclidine (PCP) in mice. PCP (10 mg/kg/day, s.c.) was administered for 14 consecutive days. Novel object recognition, forced swimming, and social interaction tests were performed during the withdrawal period of 7 days. In addition, behavioral sensitization to an acute challenge of PCP was evaluated. The parameters of the GSH-dependent system in the prefrontal cortex (PFC) were examined. MG (200 mg/kg, i.p./day) or antipsychotic clozapine (10 mg/kg, p.o./day) was administered for seven consecutive days after the final PCP treatment. PCP significantly produced abnormal behaviors, followed by increases in Nrf2 nuclear translocation, its DNA binding activity, and glutamate–cysteine ligase (GCL) mRNA expression in the PFC. PCP treatment significantly decreased GSH/glutathione disulfide (GSSG) ratio and glutathione peroxidase (GPx) activity. MG significantly attenuated abnormal behaviors and the decreases in GSH/GSSG ratio and GPx activity induced by PCP. MG attenuated the increases in Nrf2 activity and GCL expression caused by PCP. The protective potentials of MG were comparable to those of clozapine. MG ameliorates PCP-induced schizophrenia-like psychosis in mice through the positive modulation of the glutathione system.
Introduction
S
A recent report indicated that glutathione concentrations are decreased in the medial prefrontal cortex (PFC) and cerebrospinal fluid in schizophrenics. 4 Moreover, glutamate–cysteine ligase modifier subunit (GCLm)-deficient mice display schizophrenia-like behavioral activities in association with NMDA receptor hypofunction, altered dopamine neurotransmission, abnormal neuronal synchronization, and deficient myelination. 5 Furthermore, the glutathione precursor N-acetylcysteine (NAC) has been shown to have positive effects against neurochemical changes induced by GCLm gene knockout. 6
Panax ginseng Meyer (ginseng) occupies an important position in ethnic medicine in far-eastern Asian countries. 7 Although the neuropharmacological mechanisms of ginseng action need to be further elucidated, the therapeutic effects are attributable to ginseng ginsenosides, the active compounds of ginseng. We have demonstrated that ginsenosides attenuate methamphetamine (MA)-induced psychotoxicity associated with enhanced AP-1 DNA binding activity and proenkephalin expression by modulating the adenosine A2A receptor. 8 Similarly, ginsenosides were also shown to attenuate kainic acid-induced synaptosomal oxidative stress by stimulation of the adenosine A2A receptor. 9 In addition, recent reports demonstrate that ginsenoside protects against MA-induced dopaminergic toxicity by inhibition of protein kinase C δ-dependent mitochondrial oxidative stress in vivo 10 and in vitro. 11
Ginseng distributed in Korean markets is mainly supplied from mass cultivation in fields with artificial shade. 12 However, as an alternative to field cultivation, wild-stimulated methods, such as mountain cultivation, currently hold considerable interest because consumers prefer wild-stimulated ginseng. 12 –14 Furthermore, it is important that mountain cultivation techniques for ginseng should not include fungicide sprays or soil amendment applications. 15
Little is known about the effect of ginseng against the schizophrenia-like psychosis induced by PCP in mice. In this study, we selected mountain-cultivated ginseng (MG) extracts to clarify the underlying mechanism of ginseng in response to schizophrenia-like psychosis induced by PCP. We observed for the first time that a nuclear factor erythroid-2-related factor 2 (Nrf2)-dependent mechanism is important for MG-mediated pharmacological modulation in response to schizophrenia-like psychosis induced by PCP in mice. We have focused on the PFC in our study because accumulating evidence suggests that PCP may induce psychosis by mainly disturbing the function of the PFC. 16 –18
Materials and Methods
Preparation of extracts
Seven-year-old mountain-cultivated ginseng (Panax ginseng) was purchased from Pyeongchang, Korea in September 2013. The product specimens were stored at the Oriental Medical Food and Nutrition Research Laboratory, Semyung University, Jecheon, Korea. Distilled water (250 mL) was added to 1 g of powdered sample. After the fluids were refluxed twice for 2 h, the residual fluids were combined, concentrated under reduced pressure, and then MG extracts were obtained. The content of each ginsenoside in the MG extract was determined by high-performance liquid chromatography (HPLC) analyses. Details on the HPLC analysis are described in the Supplementary Data (Supplementary Data are available online at
Animals
All animals were treated in accordance with the National Institutes of Health (NIH) Guide for the Humane Care and Use of Laboratory Animals (NIH Publication No. 85-23, 1985;
Drug treatment
PCP hydrochloride (Tocris Bioscience, Ellisville, MO) was dissolved in 0.9% sterile saline, as was the MG extract. Clozapine (Sigma-Aldrich, St. Louis, MO) was dissolved in 0.1 N HCl at a concentration of 10 mg/mL and then diluted with saline. All solutions were prepared immediately before use. Mice received PCP (10 mg/kg, s.c./day) for 14 consecutive days. MG (200 mg/kg, i.p./day) or clozapine (10 mg/kg, p.o./day), as a drug control, was injected once a day for seven consecutive days after the final PCP treatment. The experimental design is illustrated in Figure 1.

Experimental design for evaluating the effects of MG or CLZ on PCP-induced schizophrenia-like psychosis in mice. CLZ, clozapine; MG, mountain-cultivated ginseng; PCP, phencyclidine.
Behavior tests
Novel object recognition, forced swimming, social interaction, and behavior sensitization tests were performed as described previously. 19 –22 MG or clozapine was administered 1 h before each behavior test. An automated video-tracking system (Noldus Information Technology, Wageningen, The Netherlands) was used to record and analyze the movements of mice in all four tests. More details are included in the Supplementary Data.
Determination of glutathione and glutathione disulfide by HPLC
Glutathione (GSH) and glutathione disulfide (GSSG) were examined using an HPLC-UV/Vis detection system (Model LC-20AT and SPD-20A; Shimadzu, Kyoto, Japan) immediately after dissecting PFC tissues, as described previously. 23 Glutathione derivatives (GSH and GSSG) were quantified in relationship to the internal standard (cysteic acid). More details are provided in the Supplementary Data.
Analysis of nuclear translocation of Nrf2
Nuclear and cytosolic fractions of PFC tissue were extracted using the NE-PER nuclear and cytoplasmic extraction kit (Thermo Scientific, Rockford, IL), according to the manufacturer's instructions. 24 More details are provided in the Supplementary Data.
Transcription factor activation assay
The DNA binding activity was assessed using the TransAM™ Nrf2 kit (#50296; Active Motif, Carlsbad, CA) according to the manufacturer's instructions. 25 More details are provided in the Supplementary Data.
Reverse transcription polymerase chain reaction
Total RNA from the PFC was isolated using the RNeasy Mini kit (Qiagen, Valencia, CA). Reverse transcription was performed as a 1-h incubation at 37°C using reactant, containing AMV reverse transcriptase and random oligonucleotide primers. 26 Details of the primer sequences, predicted product sizes, and GenBank accession numbers (where available) of the genes amplified by PCR are included in the Supplementary Data.
Determination of SOD, GPx, GR, and GST activity
Activities of superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione reductase (GR), or glutathione-S-transferase (GST) were measured as previously described, 27 –30 using homogenates of PFC tissue. For more details, please refer to the Supplementary Data.
Statistics
Data were analyzed using IBM SPSS ver. 21.0 (IBM, Chicago, IL). Statistical analyses were performed using one-way analysis of variance (ANOVA). A post hoc Fisher's PLSD test was then applied. A P value <.05 was deemed to indicate statistical significance.
Results
Effects of MG on PCP-induced adverse behavioral effects in comparison with those of clozapine
The effects of MG were evaluated according to the experimental design shown in Figure 1. Repeated treatment with PCP resulted in significant decreases (P < .05) in interaction times in the social interaction test (Fig. 2A) and recognition indexes (P < .01) in the novel object recognition test (Fig. 2B). Significant increases in immobility time (P < .05) in the forced swimming test (Fig. 2C) and a significant behavior sensitization (P < .01) (Fig. 2D) were also seen. MG treatment significantly attenuated the PCP-induced sociability deficit (P < .05) (Fig. 2A), impaired visual recognition memory (P < .01) (Fig. 2B), immobility time (P < .05) (Fig. 2C), and behavior sensitization (P < .01) (Fig. 2D). The effects of MG were comparable to those of the antipsychotic clozapine (Fig. 2).
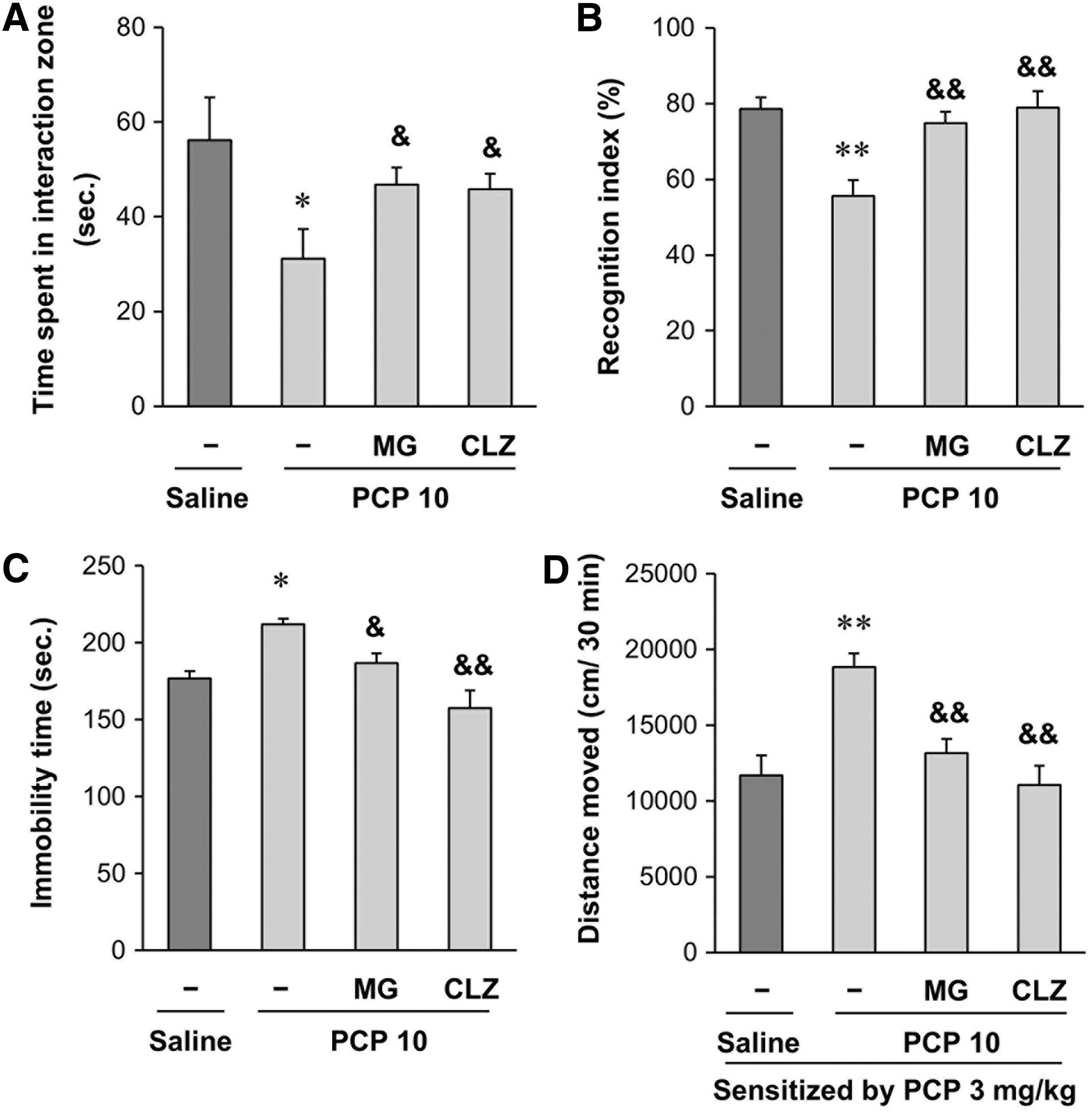
Effects of MG or CLZ on the PCP-induced abnormal alterations in the social interaction test
Effects of MG on PCP-induced changes in nuclear translocation and DNA binding activity of Nrf2 in the PFC in comparison with those of clozapine
Repeated treatment with PCP resulted in a significant decrease in cytosolic Nrf2 expression (Fig. 3A). Neither MG nor clozapine significantly altered the PCP-induced decrease in cytosolic Nrf2 expression. In contrast, PCP treatment significantly increased the nuclear translocation (P < .05) (Fig. 3B) and DNA binding activity of Nrf2 (P < .05) (Fig. 3C). Treatment with MG significantly facilitated these increases in nuclear translocation (P < .05) (Fig. 3B) and DNA binding activity of Nrf2 (P < .05) (Fig. 3C). The facilitating effects of MG were comparable to those of the antipsychotic clozapine, suggesting positive modulation due to MG and clozapine in response to PCP-induced Nrf2 activity.

Effects of MG or CLZ on PCP-induced abnormal Nrf2 nuclear translocation
Effects of MG on PCP-induced mRNA expression of GCLm and GCLc in the PFC in comparison with those of clozapine
Similar to changes in Nrf2 nuclear translocation and its DNA binding activity, repeated treatment with PCP resulted in significant increases in GCLm (P < .05) (Fig. 4A) and GCLc mRNA levels (P < .05) (Fig. 4B). Treatment with MG resulted in further upregulation in response to PCP-induced increases in GCLm (P < .05) (Fig. 4A) and GCLc mRNA expression (P < .05) (Fig. 4B). This effect of MG was comparable to that of the antipsychotic clozapine (Fig. 4A, B).

Effects of MG or CLZ on the PCP-induced abnormal alterations in the mRNA levels of GCLm
Effects of MG on PCP-induced alterations of GSH, GSSG, and the GSH/GSSG ratio in the PFC in comparison with those of clozapine
Repeated treatment with PCP resulted in significant decreases (P < .05) in concentrations of GSH (Fig. 5A) and the GSH/GSSG ratio (P < .01) (Fig. 5C), while a significant increase (P < .05) in GSSG (Fig. 5B) was observed. MG treatment significantly attenuated the PCP-induced alterations in GSH (P < .05) (Fig. 5A), GSSG (P < .05) (Fig. 5B), and the GSG/GSSG ratio (P < .05) (Fig. 5C). The positive modulation of GSH concentrations by MG was comparable to that of clozapine.
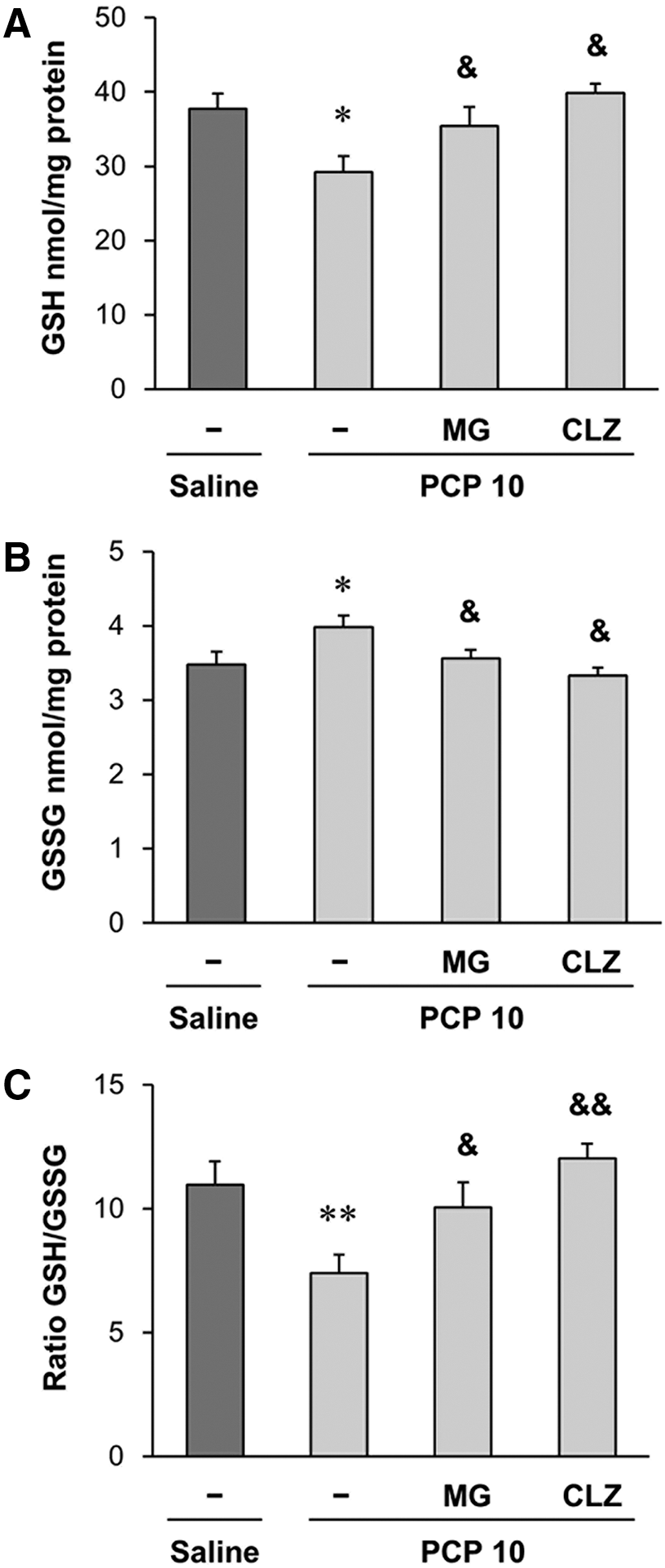
Effects of MG or CLZ on the PCP-induced abnormal alterations in GSH
Effects of MG on PCP-induced alterations in SOD-1, SOD-2 GPx, GR, and GST activity in the PFC in comparison with those of clozapine
Repeated treatment with PCP resulted in significant decreases (P < .05) in SOD-1 (Fig. 6A) and GPx activity (Fig. 6C). However, PCP treatment did not significantly affect SOD-2, GR, or GST activity (Fig. 6B, D, and E). MG treatment significantly and selectively attenuated the PCP-induced decrease in GPx activity (P < .05) (Fig. 6A). This attenuation by MG was comparable to that by clozapine.

Effects of MG or CLZ on the PCP-induced abnormal alterations in SOD-1
Discussion
This study is the first to demonstrate that MG protects against PCP-induced schizophrenia-like psychosis by Nrf2-related GSH induction in the PFC. Importantly, these protective effects of MG were comparable to those of the antipsychotic clozapine, although it remains to be determined if there are overlapping molecular mechanisms shared between MG and clozapine.
Numerous clinical and neuropsychological studies have focused on the role of PFC dysfunction in schizophrenia. 31 Anatomical and metabolic abnormalities have been observed in the frontal lobe of schizophrenic patients. 32 –34 Furthermore, the majority of cognitive impairments associated with schizophrenics seem to mainly depend on the dysfunction of the frontal cortex. 35 Previously, it was suggested that repeated PCP treatment induces glutamatergic hypofunction in the PFC of mice, as shown by decreases in extracellular glutamate release in the PFC. 3,36 In addition, we have suggested that prenatal PCP-induced NMDA receptor blockade might result in the interruption of glutamatergic neurodevelopment and consequent glutamatergic hypofunction in the PFC. 37
Accumulating evidence indicates that oxidative stress related to impairment of GSH metabolism is critical in the pathophysiology of schizophrenia. 4,38 Similar to our results, it was reported that GPx and GCL activities were decreased in schizophrenic patients, 39,40 and the GSH level was lower in the cerebrospinal fluid of schizophrenia patients without medication than in the cerebrospinal fluid of healthy subjects. 4 Furthermore, a postmortem study of schizophrenia showed that GSH levels and GPx expression were decreased in the PFC. 41 In addition, several clinical trials have indicated that NAC treatment improves symptoms or enhances the efficacy of other psychotics in patients with schizophrenia, 42,43 suggesting that NAC might be useful as an adjunctive medication. In addition, NAC was reported to attenuate psychotoxicity induced by repeated MA treatment in rats and nonhuman primates. 44,45 Thus, it is proposed here that impairment in the GSH system might be a susceptibility factor for schizophrenia.
Nrf2 is a redox-sensitive transcription factor that regulates the expression of detoxification enzymes. The Nrf2-regulated transcriptional pathways include a broad spectrum of genes, including genes encoding the antioxidants GCLm, GCLc, and GSH. 26 Therefore, Nrf2-mediated regulation of the antioxidant system is critical in protecting cells against oxidative stress. 46 It has been reported that ginsenoside Rg1 decreased the formation of reactive oxygen species induced by iron in cultured neurons through the enhancement of Nrf2 nuclear translocation and the activation of the PI3K/Akt pathway. 47 Similarly, protopanaxatriol extracted from ginseng provides neuroprotection against 3-nitropropinic acid-induced neurotoxicity by promotion of Nrf2-mediated expression of phase II antioxidant enzymes, such as heme oxygenase-1 and NADPH:quinone oxidase 1. 48 Thus, it is possible that components of ginseng positively modulates the Nrf2 transcription factor, resulting in neuroprotective conditions in vitro and in vivo. The specific neuropharmacological activity of each component of MG remains to be determined.
Since Nrf2 modulates the cytoplasmic response to oxidative stress through the transcriptional activation of genes involved in GSH synthesis, 49,50 there is a strong possibility that PCP-induced GSH depletion and the associated oxidative stress accelerate the nuclear translocation of Nrf2, GCLm, and GCLc expression, and subsequent upregulation of the target proteins related to antioxidant defense. Although this phenomenon remains to be fully elucidated, our results indicate that repeated PCP treatment triggers a compensatory mechanism, and it is more upregulated by MG posttreatment, consistent with previous data showing that ginseng induces neuroprotective effects in vitro by the positive regulation of the Keap-1/Nrf2 pathway. 51,52
Similar to our results, total ginseng extracts and total ginseng saponins were effective in attenuating depression-like behaviors induced by stress or hypercortisolism in previous studies. 53 –55 In addition, ginseng has been shown to alleviate memory impairment through the improvement of cholinergic function, the enhancement of neurogenesis, synaptic plasticity, and neurotrophic effects, and the inhibition of neuroinflammation and apoptotic processes. 56 –58 Moreover, the visual working memory of schizophrenic patients was improved by ginseng extract in a double-blind, placebo-controlled study. 59 These reports imply that ginseng might have efficacy as a therapeutic intervention against symptoms of psychiatric disorders such as schizophrenia.
In conclusion, our results suggest that MG treatment attenuates PCP-induced behavioral symptoms by the positive regulation of the Nrf2-related GSH system in the prefrontal cortices of mice. Further study is required to clarify the respective pharmacological activity of each component of MG (Supplementary Data) in response to the schizophrenia-like psychosis induced by PCP.
Footnotes
Acknowledgments
This study is supported by a grant (Project No. S111415L020100) of the “Forestry Technology Projects” provided by the Korea Forest Service and, in part, by a grant (14182MFDS979) from the Korea Food and Drug Administration, Republic of Korea. The-Vinh Tran and Yunsung Nam were supported by the BK21 PLUS program.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.