
Abstract
Skeletal muscle atrophy is a decrease in muscle mass that occurs when protein degradation exceeds protein synthesis. Leucine (Leu), an essential branched-chain amino acid in animal nutrition, regulates skeletal muscle protein metabolism. Two experiments were conducted to evaluate whether Leu could alleviate lipopolysaccharide (LPS)-induced skeletal muscle wasting by modulating skeletal muscle protein synthesis and degradation. A total of 24 rats were randomly allocated into three groups (n = 8): (1) non-challenged control; (2) LPS-challenged control; and (3) LPS +3.0% Leu. Rats were fed with control or Leu-supplemented (part of the casein was replaced with 3.0% Leu) diets throughout the trial and were injected intraperitoneally with sterile saline or LPS at days 6, 11, 16, and 21. On the morning of day 22, serum samples were collected and rats were then sacrificed for liver and muscle analysis. In vitro protein degradation, nuclear factor-κB (NF-κB) activity, and proteolytic enzyme activities of the muscles from immune-challenged rats were also measured. Our results showed that the LPS challenge resulted in not only enhanced serum interleukin-1 and liver C-reactive protein (CRP) concentrations but also decreased the average daily body weight gain and muscle fiber diameter. However, dietary Leu inclusion attenuated the increase in CRP level and the decrease in muscle fiber diameter. Importantly, the LPS challenge caused a significant elevation in the muscle proteolysis rate, but dietary Leu supplementation significantly blocked the muscle proteolysis. The mRNA expression of NF-κB, muscle atrophy F-box (MAFbx), and muscle ring finger 1 (MuRF1) was upregulated by the LPS challenge in gastrocnemius muscles, but was downregulated by Leu supplementation. Interestingly, when muscles from the LPS-challenged rats were incubated with Leu in vitro, proteasome-, calpain-, and cathepsin-L-dependent muscle proteolysis and NF-κB activity were decreased. Collectively, the data suggest that Leu supplementation could inhibit excessive skeletal muscle degradation, as well as enhance protein synthesis and, thus, attenuate the negative effects caused by the LPS-induced immune challenge.
Introduction
S
Leucine (C6H13NO2, Leu) is an essential branched-chain amino acid that cannot be de novo synthesized by animals. 9 The biochemical actions of Leu include regulating gene expression, 10,11 increasing the secretion of growth hormones, 12 and repairing muscles. 13,14 Recent nutritional studies have shown that dietary supplementation with Leu not only stimulates protein synthesis 15 –17 but also inhibits protein degradation 18 –20 in skeletal muscle. In addition, Leu may function to alleviate a variety of infections and immune challenges. 21 –24 Considering its many health benefits, Leu has received considerable research interest as an important nutritional supplement to protect muscles from degradation. However, few experiments have been conducted to study the effects of extended Leu supplementation on the skeletal muscle wasting in immune-challenged animals. Therefore, further investigation is necessary.
In the present study, we used LPS-induced immune-challenged rats as a model of in vivo rat muscle wasting conditions to investigate whether dietary Leu could alleviate muscle atrophy. Furthermore, possible mechanisms by which Leu exerted its action were studied in the gastrocnemius muscle (mostly fast-twitch glycolytic fibers) harvested from LPS-challenged control rats. Accordingly, we hypothesized that dietary supplementation with Leu potentially regulates skeletal muscle protein synthesis and degradation signaling in LPS-challenged rats and, therefore, protects against muscle atrophy.
Materials and Methods
Materials
Sprague-Dawley (SD) rats were supplied by Dossy Experimental Animals Co., Ltd. (Chengdu, China). Leu (855448-2.5G), LPS (L26637-5 MG), and calpeptin (C8999-5 MG) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Cycloheximide (94271-100 MG) was purchased from Amresco LLC (Solon, OH, USA). Cathepsin-L inhibitor IV (219433-1MGCN) and clasto-lactacystin β-lactone (426102-100UGCN) were purchased from Merck Millipore (Darmstadt, Germany). All of the other reagents were as indicated in the specified methods.
Animal care and experimental design
All experimental procedures used in the present study were approved by the Animal Care and Use Committee of Sichuan Agricultural University (Chengdu, China). Twenty-four male SD rats, weighing 190 − 200 g, were randomly divided into three groups (n = 8). The rats were individually caged in stainless-steel cages and were provided ad libitum access to water in an environmentally controlled room with a 12-h light/12-h dark cycle. All rats were fed once daily (18:00 h), and feed consumption was determined daily throughout the trial. The basal diet (Table 1) was formulated on the basis of AIN-93G requirements for growth phase of laboratory rodents. 25
The premix provided the following per kg of diets: 100 mg Zn, 50 mg Mn, 100 mg Fe, 8 mg Cu, 0.30 mg I, and 0.35 mg Se.
The premix provided the following per kg of diets: 2200 IU vitamin A, 220 IU vitamin D3, 16 IU vitamin E, 0.50 mg vitamin K3, 1 mg vitamin B1, 3.50 mg vitamin B2, 1.5 mg vitamin B6, 17.50 μg vitamin B12, 0.30 mg folic acid, 0.05 mg biotin, and 15 mg niacin.
Based on the molecular weight of the free base.
The experimental groups were as follows: (1) nonchallenged control (CON; rats fed a control diet and injected with sterile saline); (2) LPS-challenged control (LPS; rats fed the control diet and injected with LPS); and (3) LPS +3.0% Leu treatment (LPSL; rats fed a diet in which part of the casein was replaced by 3.0% Leu, and the animals were injected with LPS). After feeding the experimental diets for 5 days, the immune challenge was induced by LPS. Briefly, the challenged groups were injected intraperitoneally with 50 μg/kg body weight LPS at four times (days 6, 11, 16, and 21), while the control group was simultaneously injected intraperitoneally with the same amount of sterile saline. The Leu 26 and LPS 27,28 dose were chosen according to previous studies.
Blood and tissue sampling
At 08:00 h of day 22, the rats were weighed, before collecting blood samples in tubes. Briefly, the tubes were centrifuged at 3000 g for 15 min to isolate the serum. The serum samples were stored at −20°C until further analysis. After blood sampling, all the rats were sacrificed by cervical dislocation after ether anesthesia. Liver samples were removed immediately and frozen in liquid nitrogen and then stored at −80°C until biochemical analysis. Muscle samples were harvested and used to determine fiber size (diameter and density), protein degradation rates, and mRNA levels of nuclear factor-κB (NF-κB), MAFbx, and MuRF1. In the second experimental series (in vitro), muscles harvested from immune-challenged rats (after the fourth LPS injection) were incubated in the absence or presence of 2 mM Leu, to determine the direct effects of Leu on protein degradation rates, proteolytic enzymes, and NF-κB activity.
Biochemical analysis of serum and liver
Analyses of cytokines and immunoglobulins
All immune indices were determined using ELISA Kits (R&D Systems, Minneapolis, MN, USA). The concentrations of inflammatory factors such as IL-1 and immunoglobulins (IgA, IgG, and IgM) in the serum samples were detected following the procedures described previously. 29,30
Assay of C-reactive protein
About 0.1 g liver sample was weighed, thawed, and homogenized (5 min) in nine times the volume (w/v) of ice-cold physiologic saline. The mixtures were then centrifuged at 3000 g, 4°C, for 15 min, and the supernatants were collected and stored at −20°C until C-reactive protein (CRP) analysis. The liver CRP was assayed using a commercially available rat CRP ELISA Kit (R&D Systems, Minneapolis, MN, USA). All of the procedures were performed according to the manufacturer's instructions.
Histochemical analysis of muscle
Muscle samples for histochemical analysis were taken from an equivalent region within the gastrocnemius muscle. Transverse serial sections (10 μm) were cut from entire blocks (1.0 × 1.0 × 1.5 cm) on a cryostat microtome (HM525; Microm GmbH, Munich, Germany) at −20°C and mounted on glass slides. The slides were stained with hematoxylin and eosin (H&E) solution after fixation, and quantitative analysis of all muscle samples for fiber size (diameter and density) was performed using an image analysis program (TEMA1.04; Scanbeam, Hadsund, Denmark). All portions of the sections analyzed were free from sample disruption and freeze damage. The fiber density was calculated using the mean number of fibers per mm2.
RNA extraction, reverse transcription, and quantitative real-time PCR
Muscle (gastrocnemius) total RNA isolation, cDNA synthesis, and quantitative real-time PCR analysis were conducted as described previously. 31 Primers used for quantitative real-time PCR analysis were synthesized commercially by Invitrogen (Shanghai, China) and are listed in Table 2. GAPDH was used as the reference gene to normalize mRNA expression levels of the target genes. The relative quantification of gene expression among the treatment groups was analyzed by the 2−ΔΔCt method. 32
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; MAFbx, muscle atrophy F-box; mTOR, mammalian target of rapamycin; MuRF1, muscle ring finger 1; NF-κB, nuclear factor-κB.
Muscle incubations
Gastrocnemius muscles were gently dissected with intact tendons, mounted on stainless steel supports at resting length, and incubated under physiological conditions in a shaking water bath at 37°C for 2 h, as described in detail previously. 33 Proteolysis rates were determined by measuring the net release of free tyrosine into the incubation medium, because tyrosine is neither synthesized nor degraded in muscle samples. Cycloheximide (0.5 mM) was added to the incubation medium to prevent reincorporation of tyrosine into muscle protein. Paired muscles from the LPS-challenged rats were incubated in vitro, in the absence or presence of Leu dissolved in clasto-lactacystin β-lactone, cathepsin-L inhibitor IV, or calpeptin at 100 μM.
Proteolytic enzymes and NF-κB activities
The effects of Leu on proteolytic enzyme activities were determined by incubating paired gastrocnemius muscles harvested from the LPS-challenged rats for 2 h, as described above, in the absence or presence of Leu (2 mM). The muscles were then rinsed in physiologic saline, weighed, and homogenized (1:10, w/v) in nine volumes of ice-cold physiologic saline. The mixture of muscles and physiological saline was centrifuged at 2000 g, 4°C, for 20 min. Total protein concentration in the supernatant was measured according to Bradford 34 using bovine serum albumin as the protein standard. The 20S proteasome, calpain, cathepsin-L, and NF-κB activities were determined using commercially available assay kits (BioVision, Milpitas, CA, USA) according to the manufacturer's instructions.
Statistical analysis
The results were reported as the mean values with their standard errors. Each rat was considered as one statistical unit. Student's t-test or analysis of variance (ANOVA) followed by Holm-Sidak's or Duncan's method was used as appropriate. P < .05 was considered as statistically significant.
Results
Growth performance
The growth performance results are given in Figure 1. The LPS challenge resulted in a remarkable decrease (P < .05) in average daily body weight gain (ADG), whereas dietary supplementation with Leu had no obvious effect (P > .05) on alleviating the ADG loss caused by the LPS challenge. No significant difference (P > .05) in average daily feed intake (ADFI) among the three treatment groups was observed.
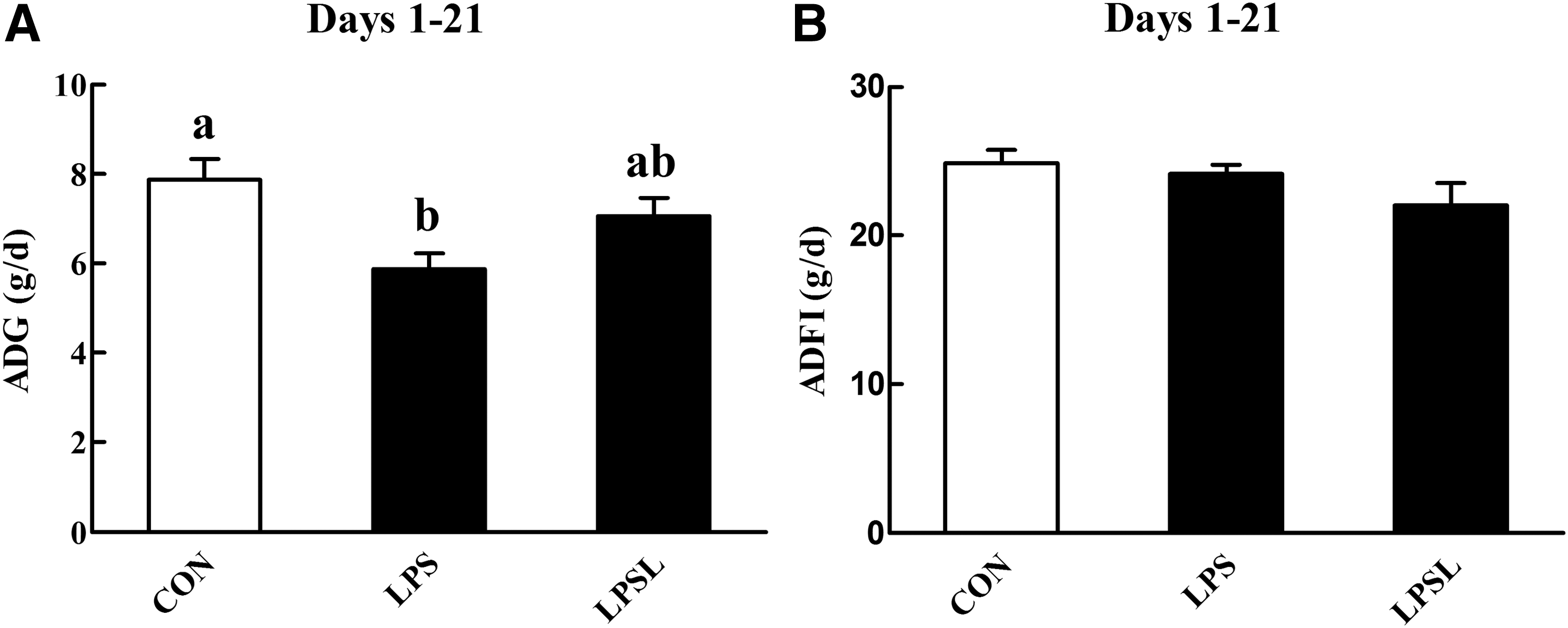
Influences of Leu on the growth performance of rats over the experimental periods.
Serum parameters
Following LPS injection, the serum IL-1 (Fig. 2A) and IgM (Fig. 2C) concentrations were increased (P < .05) by 18.76% and 16.79%, respectively. Moreover, the concentrations of IgG (Fig. 2B) and IgA (Fig. 2D) in serum did not differ (P > .05) between rats in the different treatments.
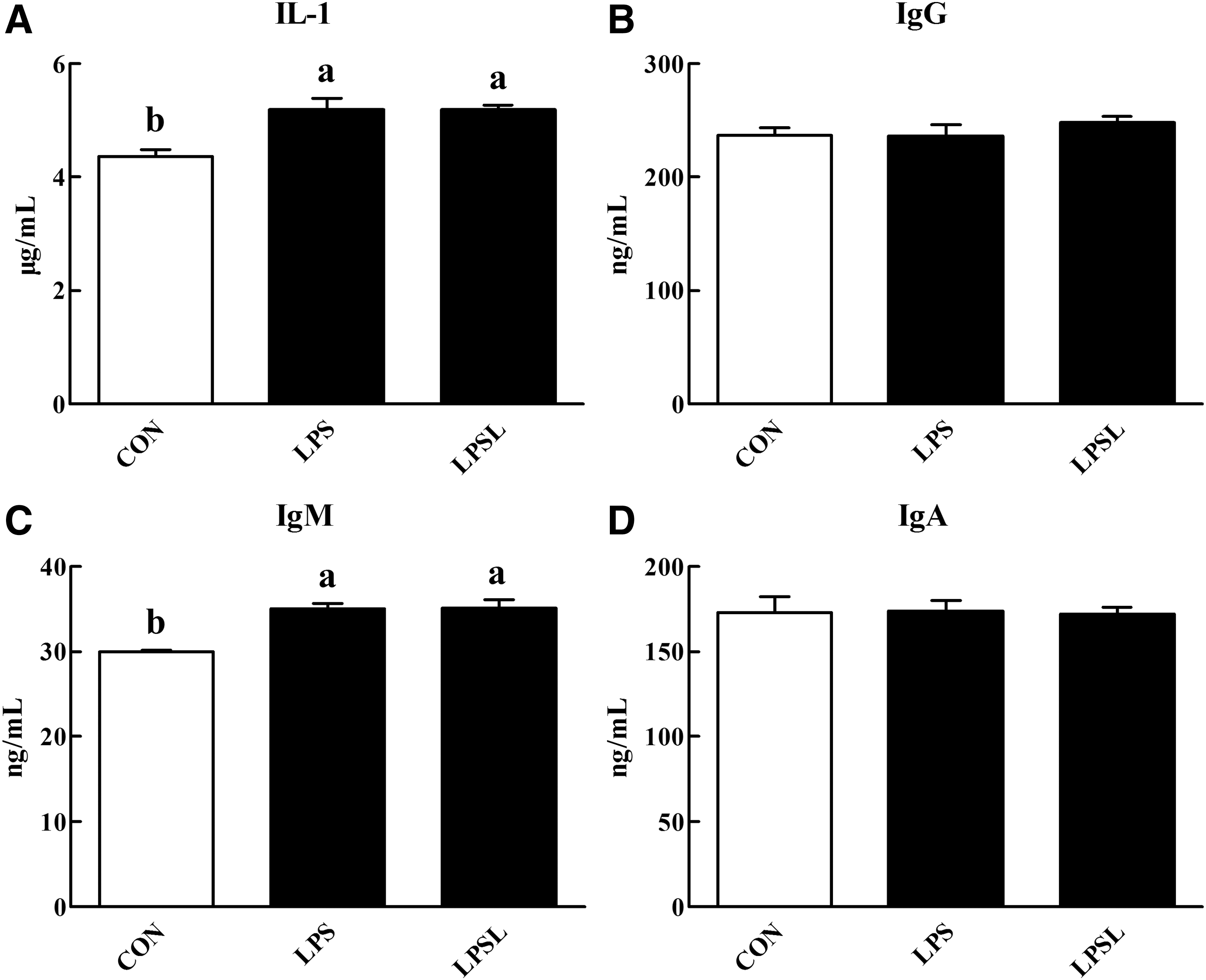
Influences of Leu on the serum parameters of rats.
Fiber size, CRP, and proteolysis rate
The H&E staining results are shown in Figure 3C. We found that LPS injection succeeded in causing musculature morphology injury. We further calculated the fiber diameter (Fig. 3A) and fiber density (Fig. 3B) of the rats in all three treatments. The LPS challenge did not affect (P > .05) the fiber density of gastrocnemius muscles, but it caused a significant decrease (P < .05) in the fiber diameter. Dietary supplementation with Leu significantly alleviated (P < .05) the decrease in fiber diameter of gastrocnemius muscles after the LPS challenge.
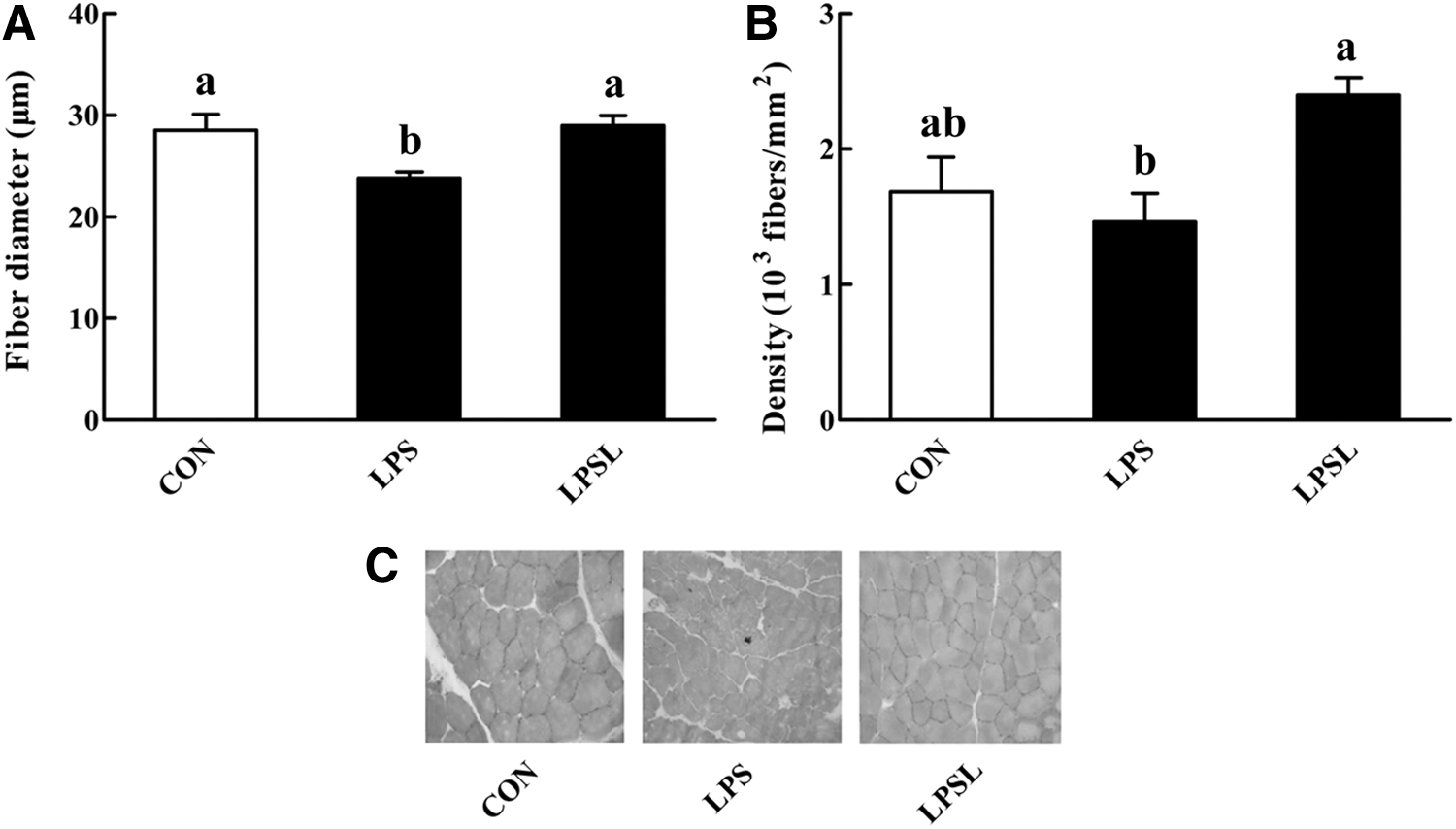
Influences of Leu on muscle fiber diameter
Relative to the CON group, LPS administration increased (P < .05) the liver CRP concentration (Fig. 4). Among the LPS-challenged rats, Leu supplementation decreased (P < .05) the liver CRP concentration.
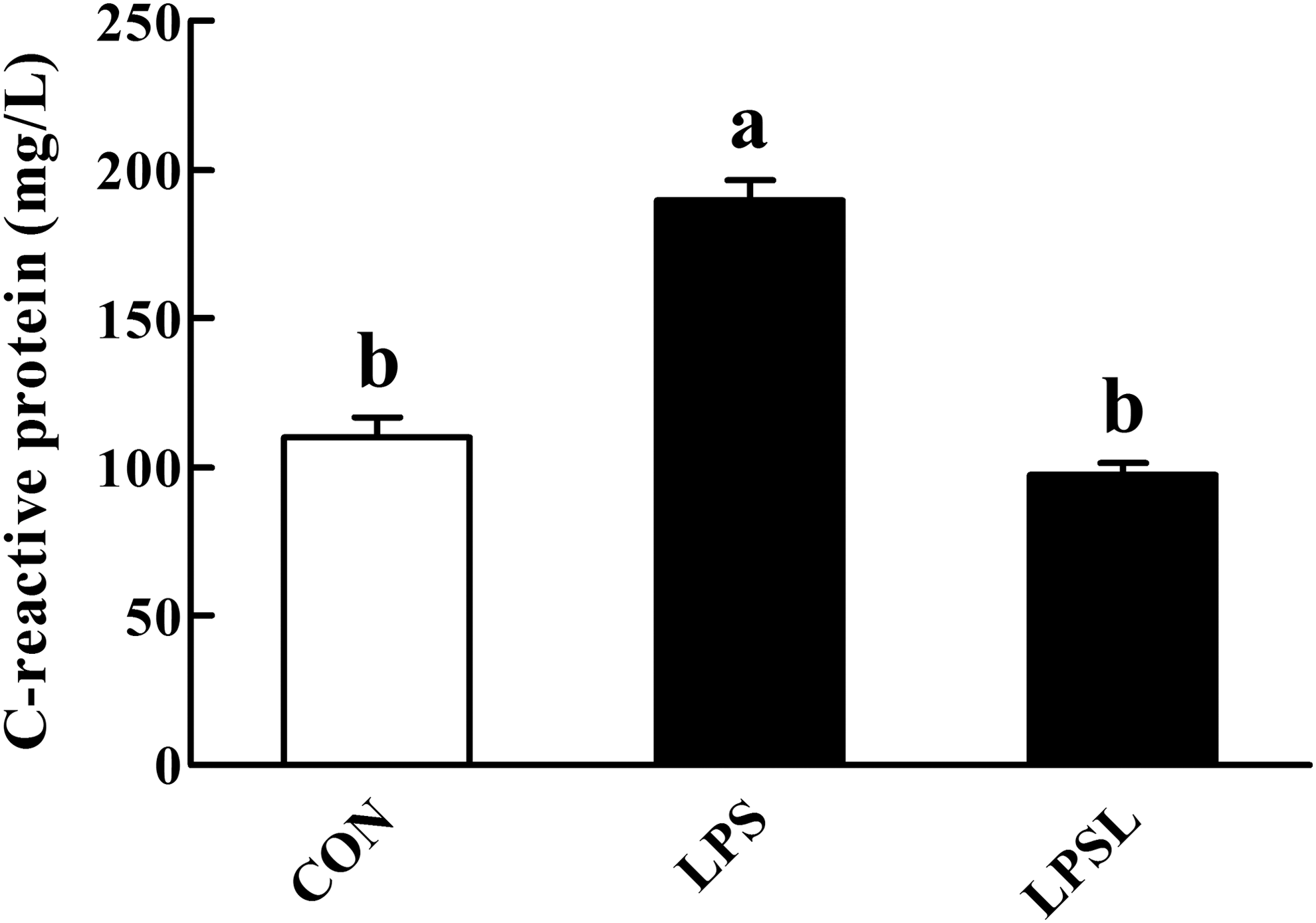
Influences of Leu on the liver C-reactive protein in rats. Values are means (eight rats/treatment), with standard errors represented by vertical bars. abMean values with unlike letters were significantly different (P < .05). CON (non-challenged control), rats receiving the control diet and injected with sterile saline; LPS (LPS-challenged control), rats receiving the control diet and injected with LPS; and LPSL, rats receiving a 3.0% Leu diet and injected with LPS.
As shown in Figure 5, the muscle proteolysis rate (24.07 ± 1.37 vs. 15.63 ± 0.95 nmol tyrosine/mg protein/h) was significantly increased (P < .05) by the LPS challenge. However, dietary supplementation with Leu significantly blocked (P < .05) the gastrocnemius muscle proteolysis caused by LPS injection.
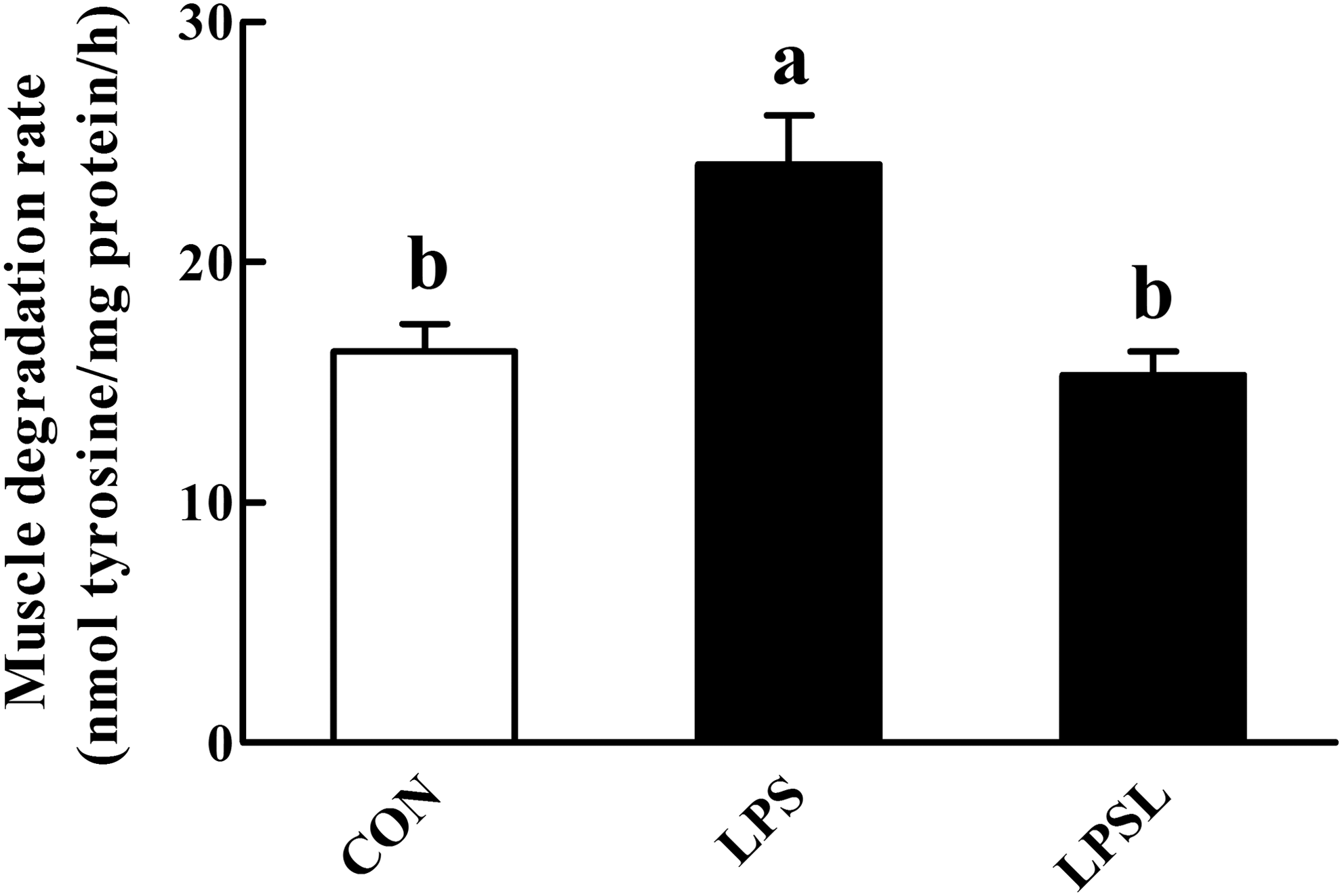
Influences of Leu on the gastrocnemius muscle degradation rate in rats. Values are means (eight rats/treatment), with standard errors represented by vertical bars. abMean values with unlike letters were significantly different (P < .05). CON (non-challenged control), rats receiving the control diet and injected with sterile saline; LPS (LPS-challenged control), rats receiving the control diet and injected with LPS; and LPSL, rats receiving a 3.0% Leu diet and injected with LPS.
Gene expression of MAFbx, MuRF1, NF-κB, and mTOR
Expression of the muscle proteolytic genes, such as the ubiquitin ligases MAFbx (Fig. 6A) and MuRF1 (Fig. 6B), increased (P < .05) at least twofold after LPS administration. Importantly, Leu ingestion resulted in significant downregulation (P < .05) of these genes in gastrocnemius muscles after LPS administration.

Influences of Leu on the relative mRNA expression of MAFbx
Compared to the CON rats, the LPS challenge significantly elevated (P < .05) the transcription of NF-κB in gastrocnemius muscles (Fig. 6C). Meanwhile, among the LPS-challenged rats, Leu supplementation caused decreased (P < .05) NF-κB transcription in the gastrocnemius muscles.
In Figure 6D, mammalian target of rapamycin (mTOR) mRNA abundance in the gastrocnemius muscles was not affected (P > .05) by LPS challenge. However, dietary Leu supplementation noticeably increased (P < .05) the gastrocnemius muscles mTOR mRNA abundance among the LPS-challenged rats.
Proteolysis by different proteolytic pathways
The influences of Leu on individual proteolytic pathways (Table 3) were assessed by treating incubated gastrocnemius muscles from LPS-challenged rats with 100 μM of the specific proteasome inhibitor clasto-lactacystin β-lactone, in the absence or presence of 2 mM Leu. Calculated as the percentage of protein degradation that was blocked by clasto-lactacystin β-lactone, Leu decreased the proteasome-dependent protein degradation by ∼59.00%. Similar experimental approaches were used to determine the influences of Leu on calpain- and cathepsin-L-dependent proteolysis. When incubated gastrocnemius muscles were treated with 100 μM of calpeptin (calpain inhibitor) or cathepsin-L inhibitor IV in the presence of Leu, the calculated calpain- and cathepsin-L-dependent protein degradation had decreased by 35.02% and 23.66%, respectively.
The inhibition of protein degradation caused by an inhibitor was calculated as the portion of protein degradation regulated by that specific proteolytic pathway. Values are means of eight replicates per treatment.
Gastrocnemius muscles were harvested from LPS-challenged control rats and incubated for 2 h in the absence of Leu.
Gastrocnemius muscles were harvested from LPS-challenged control rats and incubated for 2 h in the presence of Leu (2 mM).
Leu, leucine; LPS, lipopolysaccharide.
NF-κB and proteolytic enzyme activities
Figure 7A shows that when the LPS-challenged gastrocnemius muscles from rats were incubated in the presence of 2 mM Leu, the activity of NF-κB was remarkably (P < .05) decreased. Treatment of incubated muscles from LPS-challenged rats with Leu inhibited (P < .05) calpain (Fig. 7B), cathepsin-L (Fig. 7C), and 20S proteasome (Fig. 7D) activities. Specifically, Leu decreased calpain, 20S proteasome, and cathepsin-L activity by 43.88%, 43.89%, and 50.47%.
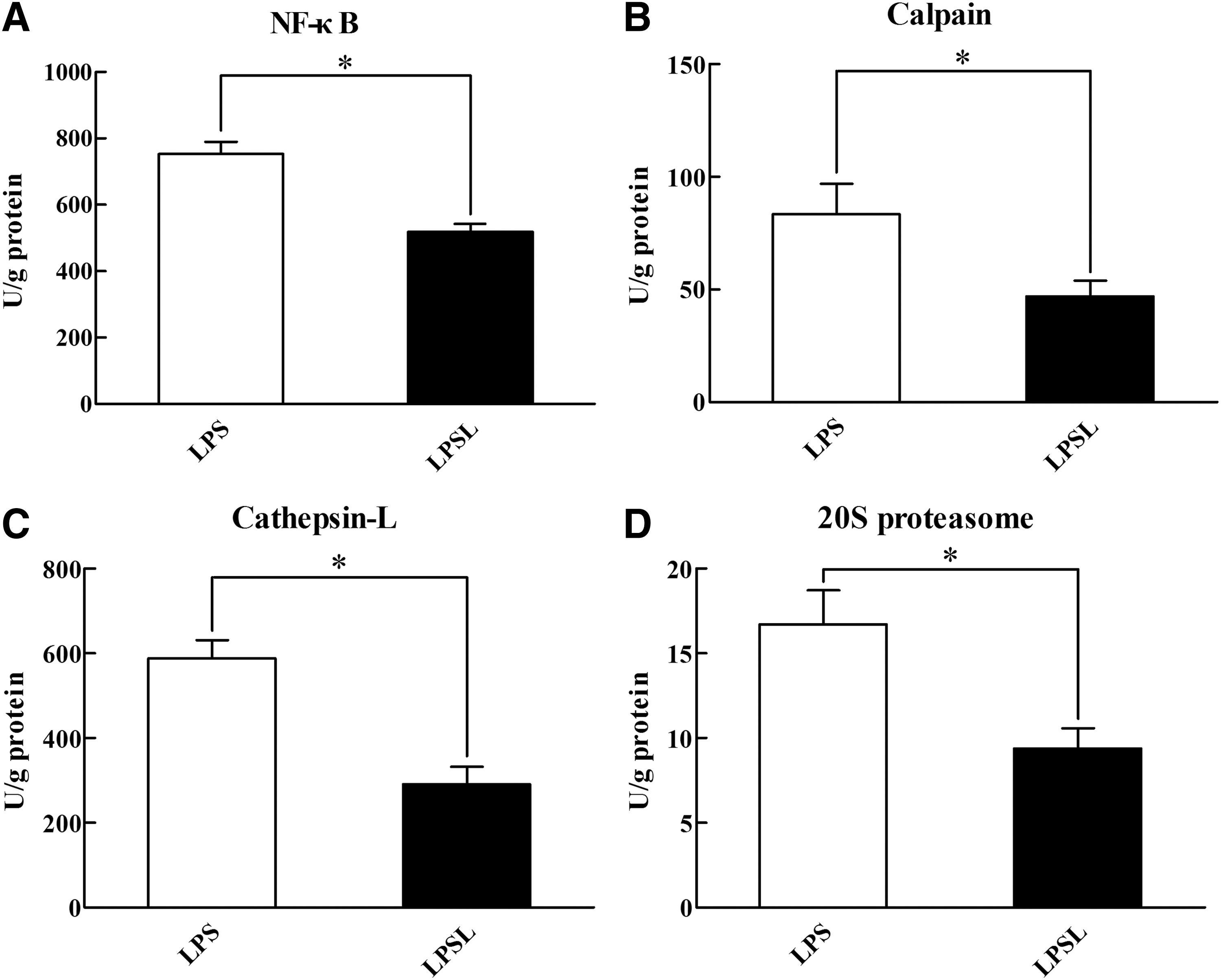
Influences of Leu on the activity of NF-κB
Discussion
An immune challenge is well established as the key limiting factor for animal production. 35 The mechanism of immune challenge on performance and protein synthesis is likely due to higher cytokine release and associated endocrine changes resulting from the immune system activation. LPS is the main virulence factors of gram-negative bacteria, 36 which can cause an increase in the release of cytokines, such as IL-1. 37 In addition, CRP is secreted by the liver in response to a variety of inflammatory cytokines. 38 In this study, an LPS-challenged rat model was used to evaluate whether Leu supplementation could alleviate the immune challenge, by inhibiting skeletal muscle proteolysis in the animals. It was shown that serum IL-1 and liver CRP levels were significantly increased after the LPS challenge. These results indicated that the LPS immune-challenged model was successful.
The LPS challenge also caused a remarkable decrease in ADG. This is similar to previous findings, which report that an LPS-induced immune challenge causes decreased growth performance. 35,39 Also, the decreased muscle fiber diameter and the elevated muscle degradation rate observed in the muscle-atrophying rats were consistent with the loss of weight and muscle protein caused by LPS administration, found in a previous study. 40 It is known that Leu supplementation can increase skeletal muscle protein synthesis by stimulating mTOR-dependent translation initiation. 41 In the present study, we also found that Leu supplementation upregulated the mTOR mRNA expression in LPS-challenged rats, suggesting that 3.0% dietary Leu supplementation facilitates protein synthesis in rats suffering from the immune challenge. These findings may provide insights into mechanisms associated with prevention of skeletal muscle wasting in LPS-challenged rats. However, further studies are necessary to clearly define the optimum levels of Leu requirements in muscle wasting animals.
The molecular mechanism by which Leu attenuated skeletal muscle wasting was evaluated by examining the role of MAFbx and MuRF1, two critical muscle-specific E3 ligases of the ubiquitin-proteasome system, which are recognized as the primary regulators of protein degradation. We observed that Leu supplementation attenuated the increased MAFbx and MuRF1 mRNA levels following the LPS challenge. It is well known that muscle protein degradation requires the proteasome and the enhanced expression of MAFbx and MuRF1, 7,42 while expression of the proteasome subunits and MuRF1 is regulated through activation of the NF-κB transcription factor. 43 Thus, we further explored the impacts of Leu on NF-κB expression. The consequent findings showed that the LPS challenge significantly elevated the NF-κB mRNA level, whereas Leu ingestion decreased the NF-κB mRNA level. Collectively, it is possible that the Leu-induced muscle protein degradation protective effects were at least, in part, associated with the decreased MAFbx and MuRF1 expression through the inhibition of NF-κB expression.
Although the ubiquitin-proteasome pathway plays a particularly important role in muscle wasting, 44,45 there is evidence that additional proteolytic mechanisms are involved. 46 For instance, previous studies suggest that the calpain- and cathepsin-L-dependent proteolysis are activated in atrophying muscle. 47,48 Therefore, we next tested whether a similar mechanism may be involved in the effects of Leu observed in the present study. When muscles from LPS-challenged rats were incubated in the presence of 2 mM Leu, significant inhibitory effects on proteasome-, calpain- and cathepsin-L-dependent protein degradation occurred, indicating that Leu can reduce the catabolic response to an LPS challenge by inhibiting multiple proteolytic pathways in skeletal muscle. A previous study showed that multiple types of skeletal muscle atrophy may involve a common program of changes in gene expression. 49 Accordingly, the activity of NF-κB in the incubated muscles was determined because this protein complex controls the expression of a large number of genes, including proteolytic enzymes. 50 As expected, Leu decreased the activity of NF-κB among the LPS-challenged rats. The inhibitory effects of Leu on the activities of three major proteolytic pathways not only validated multiple proteolytic mechanisms during muscle atrophy but also indicated that several proteolysis-associated proteins might be transcriptionally regulated by NF-κB. However, the direct link between these mechanisms and decreased muscle proteolysis is still unknown.
In conclusion, our findings suggest that Leu played beneficial roles in skeletal muscle protein synthesis signaling and in liver acute-phase protein production suppression in LPS-challenged rats. In addition, Leu supplementation reversed the deleterious changes in certain signaling molecules involved in skeletal muscle protein degradation of LPS-challenged rats, which was associated with the decreased MAFbx and MuRF1 expression through downregulation of the NF-κB-dependent signaling pathway and inhibition of the proteasome-, calpain-, and cathepsin-L-dependent protein degradation pathways.
Footnotes
Acknowledgments
We thank Shan Wu, Quyuan Wang, and Huifen Wang for their ongoing assistance during the experiments. This work was supported by the Special Fund for Agro-scientific Research in the Public Interest (201403047) and the Fok Ying-Tung Education Foundation (141027).
Authors' Contributions
Jun He participated in the experimental design and conceived the experiment. Jin Wan carried out the animal trial, performed the biochemical experiments, and participated in data interpretation. Daiwen Chen and Bing Yu contributed to the study design. Yuheng Luo, Xiangbing Mao, Ping Zheng, Jie Yu, and Junqiu Luo assisted with all data interpretation. Jin Wan was also responsible for writing the article.
Author Disclosure Statement
No competing financial interests exist.