
Abstract
Although various treatments have been used for weight loss to date, obese people rarely have safe and effective treatment options. Therefore, the antiobesity effects of several natural compounds are being actively investigated. This study was conducted to investigate the antiadipogenic effects of Monascus ruber-fermented Fagopyrum esculentum (red yeast buckwheat, RYB) in 3T3-L1 cells. We assessed the intracellular lipid content and adipocyte differentiation by oil red O staining and the expression of genes and proteins associated with adipocyte differentiation by reverse transcription–polymerase chain reaction and western blotting in 3T3-L1 cells. RYB dose dependently inhibited 3T3-L1 cell differentiation at concentrations of 50–800 μg/mL, without cytotoxic effects. It also suppressed the expression of adipogenic transcription factors, including peroxisome proliferator-activated receptor γ and CCAAT/enhancer-binding protein α, and adipocyte-specific genes, such as adipocyte fatty acid—binding protein (aP2), fatty acid synthase, and leptin, during preadipocyte differentiation into adipocytes. Furthermore, RYB reduced cyclin-dependent kinase 2 and cyclin expression and increased p21 and p27 expression, thus causing cell cycle arrest at the G1/S phase. Collectively, these results suggest that RYB may be an effective nutraceutical for weight loss as indicated by its ability to suppress adipogenesis-specific gene expression and cause cell cycle arrest at the G1/S interphase.
Introduction
A
Buckwheat (Fagopyrum esculentum Moench), belonging to the family Polygonaceae, is widely cultivated worldwide and has been used as a cereal crop in Asia. 7 It contains abundant dietary fiber, minerals, vitamins, phenolic acids, and flavonoids and has attracted interest because of its nutritional and medicinal value. 8 –10 It has also been used as a medicine according to Donguibogam, the Korean traditional medicinal encyclopedia. 11 While the antioxidant, anti-inflammatory, and anticancer effects of F. esculentum have been investigated recently, there have been few studies on its antiobesity effects.
The Monascus species, such as Monascus ruber, Monascus purpureus, Monascus pilosus, and Monascus anka, 12 have long been used for the production of fermented food and as preserving agents and traditional medicines in Asia. 13 M. ruber is known to inhibit cholesterol biosynthesis. 14 Furthermore, the secondary metabolites of M. ruber, including pigments (yellow, orange, and red), γ-aminobutyric acid (GABA), and monacolin K, have been shown to possess antibacterial, cholesterol-lowering, and anti-inflammatory activities. 15 –17
While the antiobesity effects of Monascus-fermented products have been actively researched, there have been few reports on the antiadipogenic effects of M. ruber-fermented F. esculentum. The 3T3-L1 cells have been widely used in antiobesity experiments and also for investigating the molecular mechanisms of adipocyte differentiation. 18,19
Therefore, this study was performed to investigate the effects of M. ruber-fermented F. esculentum on adipocyte differentiation and fat accumulation and their regulatory mechanisms in 3T3-L1 cells, to determine whether red yeast buckwheat (RYB) has the potential to become an antiobesity agent.
Materials and Methods
Reagents and chemicals
All cell culture materials were purchased from Gibco BRL (Gaithersburg, MD, USA). Antibodies against peroxisome proliferator-activated receptor γ (PPARγ), CCAAT/enhancer-binding protein α (C/EBPα), adipocyte fatty acid-binding protein (aP2), leptin, fatty acid synthase (FAS), cyclin-dependent kinase 2 (CDK2), cyclin D, cyclin E, p21, p27, and β-actin were purchased from Abcam (Cambridge, UK). The enhanced chemiluminescence (ECL) detection kit, Hyperfilm ECL, and nylon and polyvinylidene difluoride (PVDF) membranes were purchased from Amersham Biosciences (Buckinghamshire, UK). TRIzol® reagent for RNA isolation was purchased from Invitrogen (Carlsbad, CA, USA). All other reagents were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Preparation of RYB extract
RYB was obtained from the Cell Activation Research Institute (CARI, Korea). It was prepared by traditional methods involving the fermentation of moistened buckwheat with a proprietary strain of the food fungus, M. ruber. In brief, 30 g of RYB powder was boiled with 500 mL of deionized distilled water (DDW) at 100°C for 4 h. The aqueous extracts were then filtered through a Büchner funnel and evaporated at 60°C under vacuum. The extract was dissolved in 50 mL of DDW and freeze-dried at −60°C. The powder was stored at −20°C until use.
Cell culture and differentiation
Murine 3T3-L1 preadipocytes were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). They were cultured in Dulbecco's Modified Eagle's Medium (DMEM; Gibco, Rockville, MD, USA) supplemented with 10% fetal bovine serum (FBS; Gibco) and 1% penicillin–streptomycin (Gibco) in a humidified 5% CO2 atmosphere at 37°C. When the cells were confluent (designated as day 0), cellular differentiation was induced using DMEM supplemented with 10% FBS, 1 μM dexamethasone (DEX; Sigma-Aldrich, St. Louis, MO, USA), 0.5 mM 1-methyl-3-isobutylmethylxanthine (IBMX; Sigma-Aldrich), and 10 μg/mL insulin (INS; Sigma-Aldrich). The cells were then treated with different concentrations of RYB (0, 50, 100, 200, 400, and 800 μg/mL). After 48 h (day 2), the culture medium was replaced with DMEM supplemented with 10% FBS, and this was repeated every 2 days until day 8.
Determination of cell viability
We determined the effect of different concentrations of RYB on 3T3-L1 preadipocyte viability using a Cell Counting Kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Tokyo, Japan). In brief, the cells were seeded in a 96-well plate at a density of 1 × 104 cells/well and treated with RYB (0–800 μg/mL) for 24 h. To measure the cell viability, CCK-8 reagent was added to each well and the optical density was measured at 450 nm using an Infinite® F50 microplate reader (Tecan, Männedorf, Switzerland). The viability of RYB-treated cells was expressed as a percentage of the control cell viability.
Oil red O staining
Lipid accumulation in 3T3-L1 cells, which indicates the extent of cell differentiation, was measured by oil red O staining. In brief, differentiated 3T3-L1 cells were fixed in 10% formaldehyde in phosphate-buffered saline for 1 h, washed with distilled water, and dried completely. The cells were then stained with 0.5% oil red O solution in 60:40 (v/v) isopropanol:triple distilled water for 15 min at room temperature, washed four times with triple distilled water, and dried. The treated cells were observed under an Olympus microscope (BX51, Tokyo, Japan) and representative images were captured using an Olympus DP70 camera. Cell differentiation was quantified by elution of the stain with isopropanol and measurement of the optical density at 520 nm.
Reverse transcription–polymerase chain reaction
Reverse transcription–polymerase chain reaction (RT-PCR) was performed to measure the mRNA expression of PPARγ, C/EBPα, aP2, leptin, FAS, CDK2, cyclin D, cyclin E, p21, and p27. In brief, on day 8 after differentiation, total RNA was extracted from 3T3-L1 cells using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). cDNA was synthesized from 1 μg of total RNA using a First-Strand cDNA Synthesis Kit (Invitrogen), following the manufacturer's recommended protocol. The polymerase chain reaction (PCR) was conducted using an ABI 2720 Thermal Cycler (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA), with the following reaction conditions: initial denaturation at 95°C for 5 min followed by 30 amplification cycles consisting of denaturation for 40 s at 95°C, annealing for 40 s (temperature 56–62°C), and extension for 1 min at 72°C. The oligonucleotide primer sequences used are as shown in Table 1. The PCR products were separated on an agarose gel (1.5%) by electrophoresis for 30 min at 100 V. The bands were visualized, and their relative intensities were analyzed using the ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Western blot analysis
Cell lysates from each sample (30 μg) were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE, 10% gel) and transferred to PVDF membranes. The membranes were blocked overnight with 5% skim milk in Tris-buffered saline-Tween 20 (TBST, 20 mM Tris-HCl, pH 7.6, 140 mM NaCl, and 0.1% Tween 20) and incubated with the following primary antibodies (1:1000 dilution): PPARγ, C/EBPα, aP2, FAS, leptin, CDK2, cyclin D, cyclin E, p21, p27, and β-actin (Abcam, Cambridge, UK). The membranes were then washed four times with TBST buffer and incubated with the corresponding horseradish peroxidase-conjugated secondary antibody (1:2000 dilution). The immunoreactive protein bands were visualized using an enhanced chemiluminescence kit (Amersham Pharmacia Biotech, Buckinghamshire, UK). The relative intensities of the bands were analyzed using the Image J 1.41 Software.
Statistical analysis
The results are expressed as the mean ± standard deviation (SD) of three independent experiments. Statistical significance was determined by one-way analysis of variance with Duncan's multiple range tests, using the GraphPad Prism 4.0 software (GraphPad, Inc., San Diego, CA, USA). Values of P < .05 were considered statistically significant. The statistical software package SPSS 10.0 (SPSS institute, Chicago, IL, USA) was used for these analyses.
Results
Effects of RYB on 3T3-L1 cell proliferation
When the cytotoxicity was evaluated in 3T3-L1 cells treated with different concentrations (50–800 μg/mL) of RYB for 24 h by CCK-8 assay, RYB was not cytotoxic in the concentration range tested (Fig. 1).
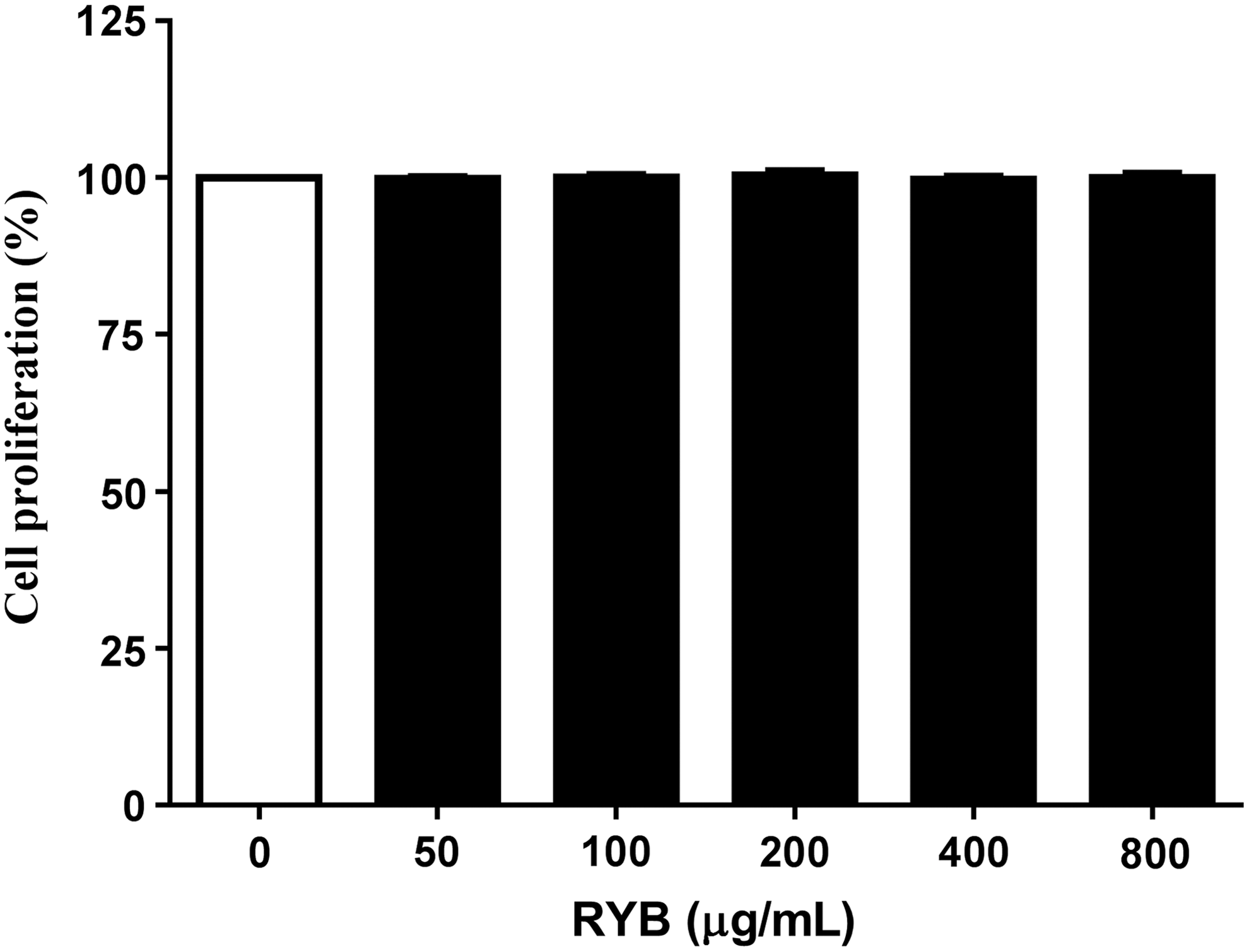
Effect of RYB on 3T3-L1 cell viability. 3T3-L1 preadipocytes were incubated with RYB extracts (0–800 μg/mL) for 24 h. Posttreatment cell viability was determined by Cell Counting Kit (CCK)-8 assay. Values are presented as mean ± SD (n = 3). Viability of untreated controls was set to 100%. RYB, red yeast buckwheat extract; SD, standard deviation.
Effect of RYB on intracellular lipid accumulation in 3T3-L1 cells
The inhibitory effect of RYB on 3T3-L1 preadipocyte differentiation was assessed by oil red O staining, which measured the change in lipid accumulation during the differentiation of preadipocytes into adipocytes. The oil red O-stained cell population decreased gradually with increasing concentrations of RYB (50–800 μg/mL) (Fig. 2A). The intracellular lipid content of differentiated adipocytes was significantly decreased to 6%, 12%, 22%, 28%, and 51% in response to 50, 100, 200, 400, and 800 μg/mL RYB, respectively (Fig. 2B), over a period of 6 d, indicating that RYB dose dependently suppressed intracellular lipid accumulation.
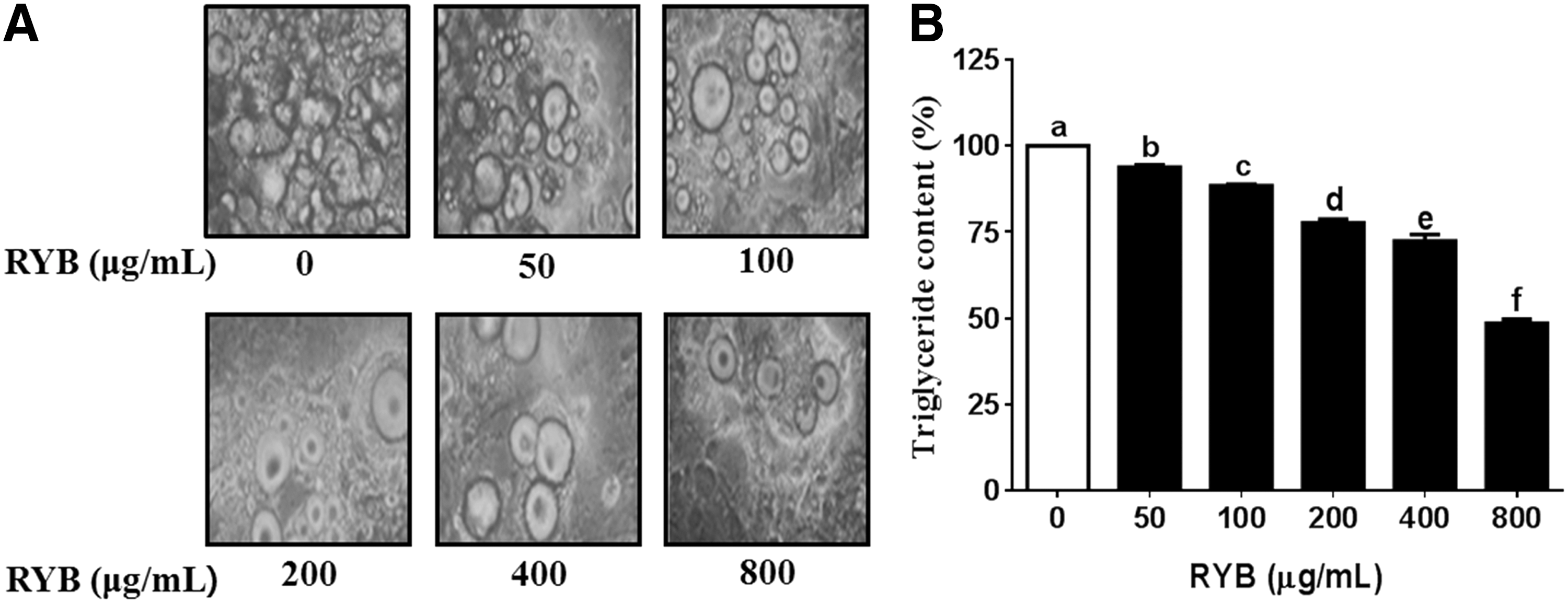
Effect of RYB on adipocyte differentiation in 3T3-L1 cells.
Inhibitory effect of RYB on adipogenesis-specific gene and protein expression in 3T3-L1 cells
We measured the expression of PPARγ, C/EBPα, aP2, FAS, and leptin using RT-PCR to investigate whether RYB can regulate adipogenesis in 3T3-L1 cells. The expression of PPARγ mRNA was dose dependently decreased to 3%, 27%, 46%, 60%, and 77% of the expression in untreated cells, in response to 50, 100, 200, 400, and 800 μg/mL RYB, respectively (Fig. 3B). The expression of C/EBPα mRNA was decreased to 4%, 29%, 46%, 61%, and 78% of the expression in untreated cells, in response to 50, 100, 200, 400, and 800 μg/mL RYB, respectively (Fig. 3C). While the aP2 mRNA expression was not decreased after treatment with 50 μg/mL RYB, it was significantly decreased (13–51%) after treatment with RYB concentrations greater than 100 μg/mL (Fig. 3D). FAS mRNA levels were decreased to 30–80% of the expression in untreated cells (Fig. 3E). The expression of leptin mRNA was dose dependently decreased after treatment with 50, 100, 200, 400, and 800 μg/mL RYB to 17%, 54%, 82%, 88%, and 92%, respectively (Fig. 3F).
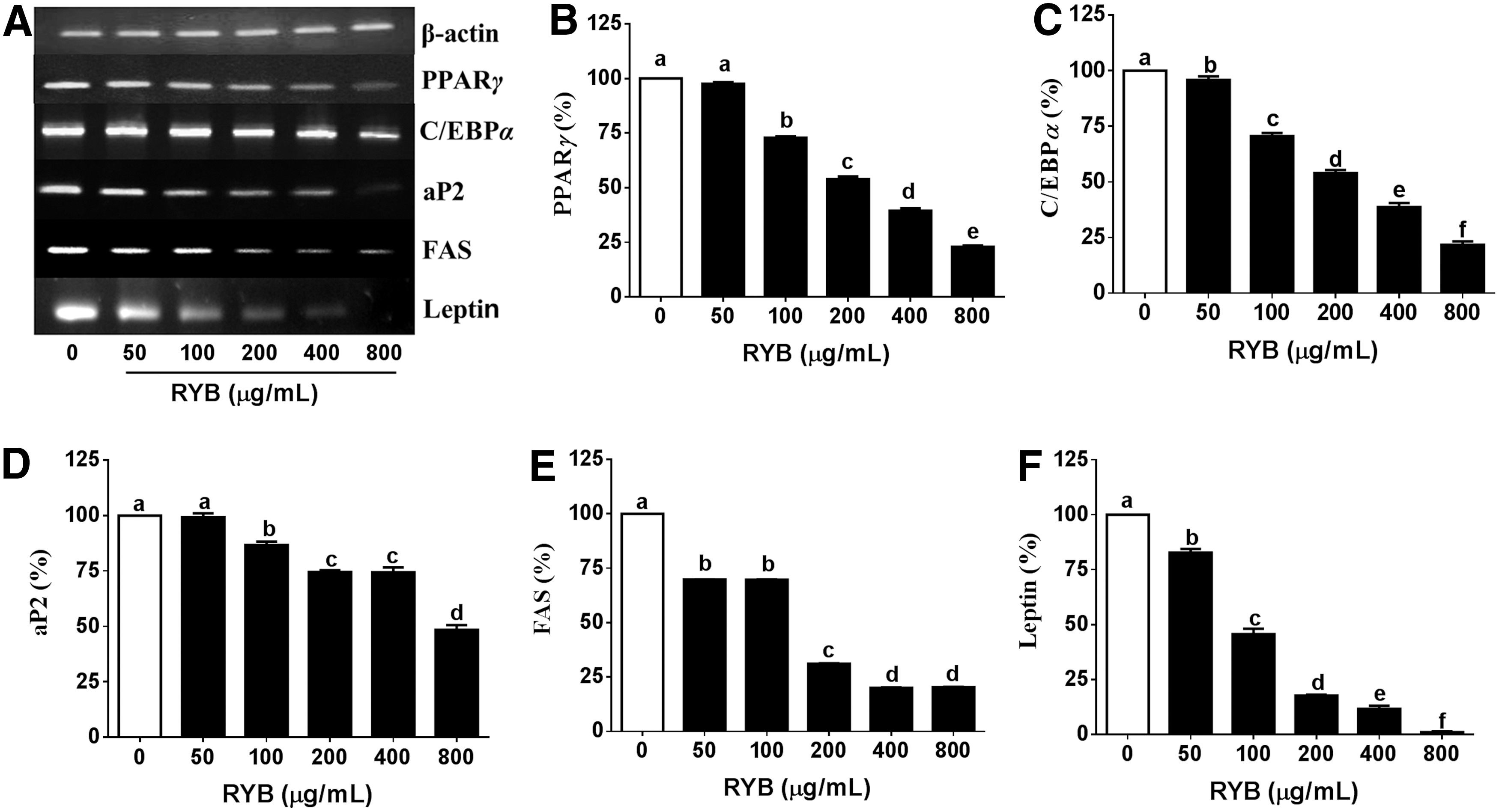
Effect of RYB on the mRNA expression of major adipogenic genes in 3T3-L1 cells.
Next, we measured the expression of proteins associated with preadipocyte differentiation and fat accumulation via western blotting, to investigate the mechanisms underlying the antiadipogenic effect of RYB in 3T3-L1 cells. The protein expression of PPARγ and C/EBPα was decreased in response to RYB treatment with maximum reduction observed at a concentration of 800 μg/mL RYB (Fig. 4B, C). However, PPARγ expression did not change in response to 50 μg/mL RYB. RYB treatment also significantly decreased the expression of aP2, FAS, and leptin in a dose-dependent manner, in comparison to the expression in untreated cells (Fig. 4D–F). However, the levels of aP2, FAS, and leptin showed no difference in response to RYB doses between 0 and 50 μg/mL.
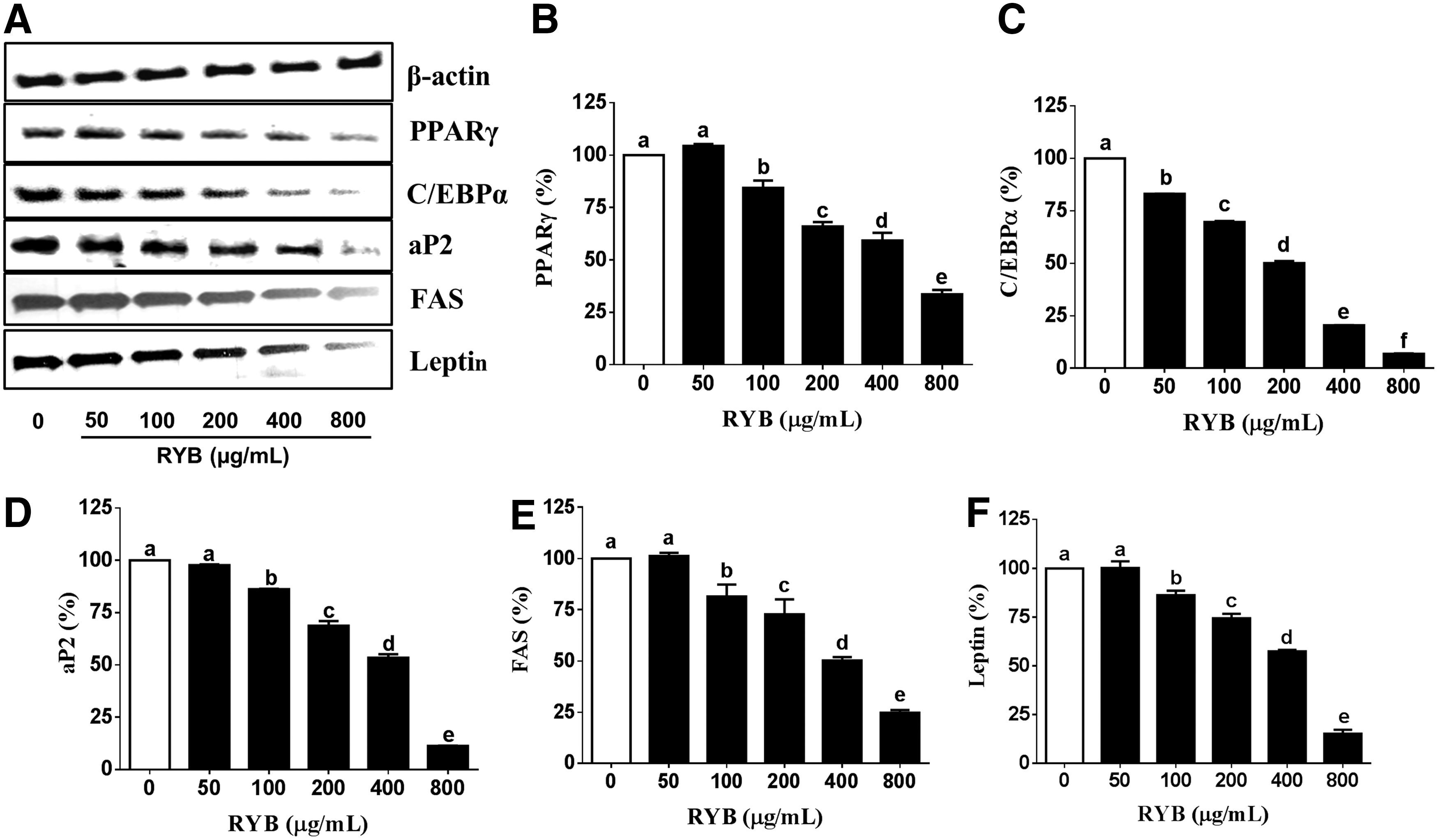
Effect of RYB on the protein expression of major adipogenic transcription factors in 3T3-L1 cells.
Effect of RYB on cell cycle progression in 3T3-L1 cells
To study the effect of RYB treatment on cell cycle progression, we examined the expression of cell cycle regulators, such as CDK, cyclins, p21, and p27, in 3T3-L1 cells. CDK2 expression was dose dependently decreased (19–62%) by RYB treatment, in comparison with the expression in untreated cells (Fig. 5B). The cyclin D mRNA expression was dose dependently decreased to 23%, 29%, 31%, 46%, and 56% of the expression in untreated cells, in response to 50, 100, 200, 400, and 800 μg/mL RYB, respectively (Fig. 5C). The cyclin E mRNA expression was also decreased to 16%, 19%, 20%, 41%, and 65% of the expression in untreated cells, in response to 50, 100, 200, 400, and 800 μg/mL RYB, respectively (Fig. 5D). On the contrary, the expression of p21 significantly increased after RYB treatment when compared to untreated cells but showed no dose-dependent response between 50 and 400 μg/mL (Fig. 5E). As shown in Figure 5F, p27 mRNA expression was significantly increased by RYB treatment in a dose-dependent manner. Furthermore, the expression of cyclin D, p21, and p27 proteins in RYB-treated 3T3-L1 cells followed the same trend as their respective mRNA expression (Fig. 6C, E, F). The expression CDK2 and cyclin E in response to 800 μg/mL RYB was significantly decreased to 73% and 38%, respectively, of the expression observed in response to 0, 50, 100, 200, and 400 μg/mL RYB (Fig. 6B, D). Conversely, RYB treatment significantly increased the expression of p21 and p27 proteins in a dose-dependent manner (Fig. 6E, F).
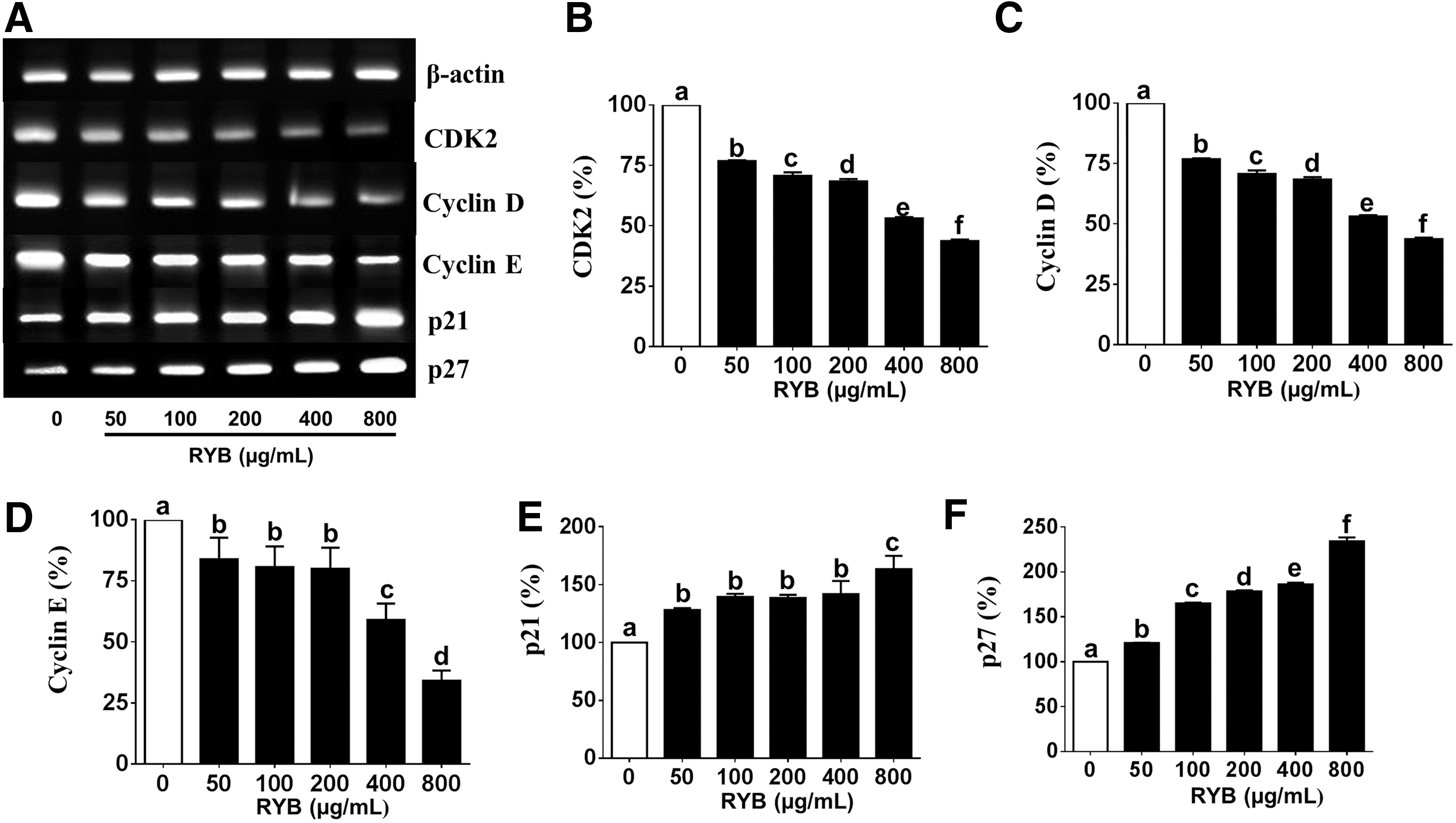
Effect of RYB on the mRNA expression of major cell cycle regulators in 3T3-L1 cells.
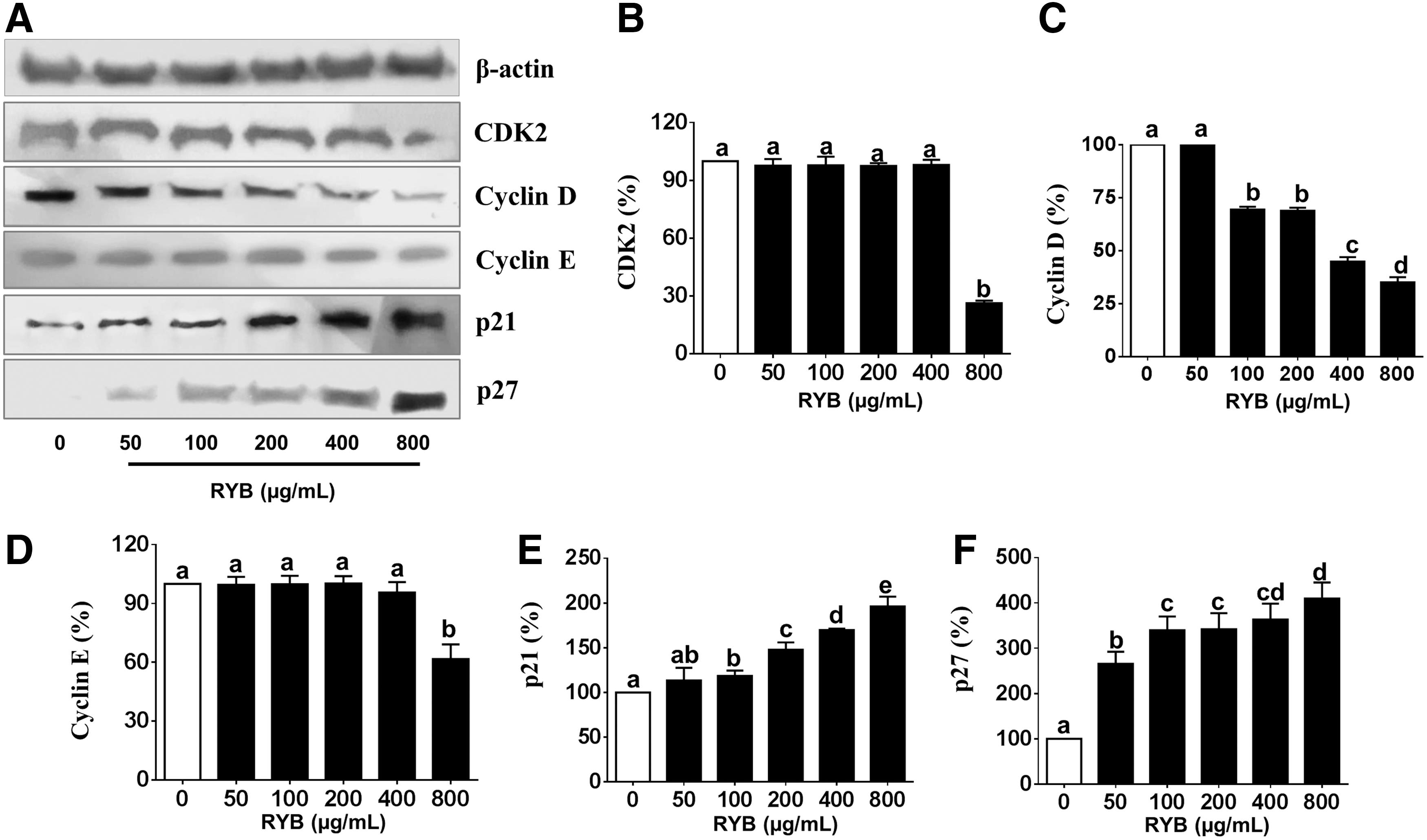
Effect of RYB on the protein expression of major cell cycle regulators in 3T3-L1 cells.
Discussion
In this study, we investigated the effects of RYB on adipocyte differentiation and its regulatory mechanisms in 3T3-L1 cells. Our results demonstrated that RYB exhibits an antiadipogenic effect by downregulating the expression of adipogenic transcription factors and adipocyte-specific genes, leading to decreased adipocyte differentiation and intracellular lipid accumulation in 3T3-L1 cells.
Adipogenesis is regulated by a complex network of transcription factors. We demonstrated that RYB treatment reduced the intracellular lipid accumulation in 3T3-L1 cells, which is consistent with the findings of previous reports. Anka, a fermented rice product of Monascus species, has been reported to reduce serum triglycerides and adipose tissue weight in rats. 20 The fermented mixture of buckwheat and soybean with rice-koji salt has been reported to reduce body weight gain and fat pad mass in high-fat diet-induced obese mice. 21 In addition, Monascus-fermented red ginseng has been reported to dose dependently reduce fat pad mass and abdominal fat thickness in high-fat diet-induced obese mice. 22 M. pilosus-fermented black soybean has been reported to inhibit lipid accumulation in 3T3-L1 cells and in high-fat diet-induced obese mice. 23
Monascus-fermented rice is known to contain compounds of the monacolin group such as lovastatin, a 3-hydroxy-3-methyl-glutaryl coenzyme A (HMG-CoA) reductase inhibitor, 14,20 which has been reported to inhibit adipogenesis in 3T3-L1 cells. 24
Our results also showed that RYB treatment significantly suppressed the mRNA and protein expression of PPARγ and C/EBPα in 3T3-L1 cells. The transcription factors PPARγ and C/EBPα play a key role in regulating adipogenesis and are expressed in the late stages of differentiation. 25 C/EBPα appears to promote adipocyte differentiation in conjunction with PPARγ by cross-regulation. 26 Moreover, they are needed to activate the expression of adipocyte-specific genes, including aP2, FAS, lipoprotein lipase, adiponectin, and leptin, which induce triglyceride synthesis. 27 We examined whether RYB could regulate the expression of aP2, FAS, and leptin proteins that are activated by PPARγ and C/EBPα during adipocyte differentiation. 25 aP2, also called the fatty acid binding protein 4 (FABP4), plays a critical role in adipocyte differentiation. 28 In this study, RYB treatment significantly inhibited the expression of aP2, FAS, and leptin proteins that are responsible for the morphological changes and fat accumulation in 3T3-L1 cells during adipogenesis. These results showed that the downregulation of PPARγ and C/EBPα expression caused the suppression of aP2, FAS, and leptin expression.
The expression of cell cycle regulators, including CDK2, cyclin D, and cyclin E was assessed to investigate whether RYB treatment affected the cell cycle progression. RYB treatment was observed to dose dependently inhibit the expression of these genes. CDK2, in association with cyclins D and E, controls the G1 to S phase transition during mitotic clonal expansion (MCE). 29 Our results showed that RYB inhibited the MCE process as evidenced by the impaired G1 to S phase transition and reduced expression of cell cycle regulators, including CDK2 and cyclins. These results agreed with several studies that showed that the inhibition of CDK2 and cyclin A expression by treatment with EGCG, glucosamine-6-phosphate, and curcumin blocked the G1 to S phase transition in the cell cycle. 27
In the present study, RYB treatment increased the expression of p21 and p27, which are major CDK inhibitors, in 3T3-L1 cells. The activation of p21 and p27 has been reported to cause G1 cell cycle arrest and inhibition of cell proliferation. 30 –32 Moreover, the fat pad weight and adipocyte number of p27 knockout, p21 knockout, and p27/p21 double knockout mice were reported to be increased in comparison with those observed in control mice. 33
Our results are consistent with these findings and demonstrate that RYB treatment inhibits cell proliferation and differentiation by causing cell cycle arrest at the G1/S interphase via the downregulation of cell cycle regulator expression and upregulation of CDK inhibitor expression.
Collectively, our findings suggest that RYB may be an effective nutraceutical for weight loss, as indicated by its ability to suppress adipogenesis-specific gene expression and cause cell cycle arrest at G1/S interphase.
Further animal studies are required to conclusively establish a causal association between RYB consumption and antiadipogenic effects.
Footnotes
Acknowledgment
This study was supported by Konkuk University.
Author Disclosure Statement
No competing financial interests exist.