
Abstract
This study was designed to investigate the antiobesity effects of Salvia plebeia R. Br. ethanolic extracts (SPE) in mice fed high-fat diets (HFD). Male C57BL/6J mice were randomly assigned to four groups: normal diet (Chow), high-fat diet (HFD, 45% fat), HFD+SPE 200 (200 mg/kg b.w.), and HFD+SPE 400 (400 mg/kg b.w.). Extracts were administered orally every day for 8 weeks. Increases in body/fat weight and feed efficiency ratio were monitored in all mice. In addition, obesity resulting from feeding HFD to the mice was confirmed by the increase of glucose level, aspartate transaminase, alanine transaminase, triglyceride (TG), high-density lipoprotein cholesterol, very low-density lipoprotein-c, leptin, and adiponectin in blood. The SPE-treated mice gained less body and mesenteric/subcutaneous adipose tissues weights and had lower TG, very low-density lipoprotein cholesterol, leptin, and glucose level in serum, compared to the HFD group. Moreover, histopathological examinations revealed that the size of adipocytes in liver and adipose tissue was significantly decreased by SPE, compared to the HFD group. The expression of adipogenesis transcription factors (e.g., peroxisome proliferator activated receptor γ and CCAAT/enhancer binding protein α) and lipogenesis-related target genes (adipocyte fatty acid-binding protein 2, lipoprotein lipase, fatty acid synthase, and sterol regulatory element-binding transcription factor 1c) in HFD-induced obese mice was decreased by SPE treatment. These results suggest that SPE attenuates the fat accumulation in HFD-induced obese mice by suppressing the expressions of genes related to adipogenesis and lipogenesis activity. Therefore, SPE could be developed as a potential therapy for reduction of body weight and antiobesity intervention.
Introduction
O
Salvia plebeia R. Br., also known as common sage, is an annual/biennial plant belonging to the Labiatae (mint) family and grows naturally throughout the entire area of Korea. Young leaves of Salvia plebeia are edible and have been used as a herbal medicine in Korean traditional medicine for hepatitis, cough, diarrhea, gonorrhea, menorrhagia, tumors, and hemorrhoids. 6,7 Salvia plebeia extract (SPE) was reported to have antioxidant, antibacterial, antiallergic, anti-inflammatory, and anticholesterol activity. 8,9 SPE contains caffeic acid, luteolin, luteolin-7-O-glucoside, rosmarinic acid, hispidulin, hispidulin-7-O-glucoside, and nepetin, as well as flavonoids, polyphenols, saponins, cardiac glycosides, unsaturated sterols, and essential oils. 10,11 Hispidulin, a flavone in SPE, plays a role as a receptor ligand for benzodiazepines, antithrombotic/antifungal drugs, and activates AMPK (adenosine monophosphate-activated kinase). 12 In addition, hispidulin inhibits the growth of cancer cells by suppression of mTOR and induction of p21 in glioblastoma multiform brain tumors. 13 Several researchers have reported antiobesity activities of homoplantaginin, luteolin, and rosmarinic acid isolated from Salvia plebeia. 14 –16
The aim of this study was to investigate the availability of SPE for functional food materials and food additives with antiobesity activity. Antiobesity activity of SPE was evaluated by measuring weight changes of fat tissues (body, liver, kidney, and epididymis), contents of blood lipid and leptin, the suppressive activity of adipocyte differentiation, and changes in adipocyte tissue.
Materials and Methods
Materials
Dried Salvia plebeia R. Br. was collected in April 2014 from a farm in Yeoju, Korea. Voucher specimens of Salvia plebeia R. Br. were deposited at the research institute of Gyeonggi-Do Agricultural Research and Extension Services (Yeoju, Korea). In addition, some aerial part of Salvia plebeia was deposited in the Research Institute of YD Life Science Co. Ltd. (Voucher specimens No. GC-Y-201404). Salvia plebeia (32 kg) was extracted twice with 50% (v/v) ethanol for 12 h at 50°C and then was filtered through filter paper (Whatman No. 3). 17 Filtrates were evaporated under vacuum at 50°C and lyophilized to remove the residual solvents. The yield of Salvia plebeia extract (SPE, Code name: YD-204B) was ∼24% (w/w), and 50 mg of extract was obtained from 210 mg of dried Salvia plebeia R. Br. Hispidulin, which was a reference compound in SPE, and was detected to be ∼0.39% ± 0.08% (w/w).
Animals and diets
Fifty 8-week-old male C57BL/6N mice (120–140 g) were obtained from Orient Bio Co., Ltd.. Mice were kept in an air-conditioned room at 23°C ± 1.0°C and 55% ± 5% relative humidity with a light/dark period of 12 h for 1 week. All mice were treated in accordance with the Guide of the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, Commission on Life Sciences, National Research Council, 1996), as approved by the Institutional Animal Care and Use Committee, Research Institute, YD Life Science.
For induction of obesity, mice were fed with a high-fat diet (D12451; Research Diets, Inc.) containing 45% of energy from fat as total calories for 8 weeks, except for the normal diet group (CD, D12450B; Research Diets, Inc.). The mice were randomly divided into five groups (n = 10/group): each treated with (1) Normal diet group (Chow), (2) Control group (high-fat diet [HFD], 45 kcal fat), (3) Orlistat-treated group (50 mg/kg, b.w.) with high-fat diet (positive control), (4) SPE 200 mg/kg body weight (b.w.)-treated group with high-fat diet, and (5) SPE 400 mg/kg b.w.-treated group with high-fat diet. All samples were dissolved in freshly distilled water and were administered orally once daily for 8 weeks. Distilled water was freely available during experiments.
Body weight and feed efficiency ratio
During the feeding period, the mice and food intake were monitored every day and were weighed every 3 days. All diets were supplied to mice at the same time every day, and the amounts of diets that mice ate were calculated from the amount of diets remaining in a cage. Feed efficiency ratio (FER) was calculated as follows;
Weight of liver and total adipocytes
After the termination of the experiment and fasting for 12 h, the mice were anesthetized with ether and major body organs were examined visually through the postmortem. Blood was removed totally by injection of PBS into blood vessels, and the isolated liver was washed with PBS and weighed. In addition, the epididymal, subcutaneous, kidney, mesenteric, and retroperitoneal adipose tissues (ATs) were dissected out, weighed, and were examined visually. All tissue samples were frozen in liquid nitrogen before storage at −80°C until assayed.
Biochemical analysis
Blood taken from the heart was held at room temperature for >30 min, and the serum was separated by centrifugation at 2,465 g for 20 min at 4°C. Alanine aminotransferase (ALT), aspartate aminotransferase (AST), triglyceride (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL-c), low-density lipoprotein cholesterol (LDL-c), and glucose levels in serum were determined with an automatic analyzer (DRI-CHEM 4000i; Fujifilm Co., Tokyo, Japan). The plasma concentrations of leptin were determined enzymatically using a commercial ELISA Kit (R&D System).
Histological examination
Specimens of liver tissue, epididymal AT, and subcutaneous AT were fixed for >24 h in 10% buffered formalin, dehydrated, and embedded in paraffin. Paraffin blocks of liver and ATs were cut at the thicknesses of 5 and 8 μm, respectively, and then paraffin was removed with xylene. Tissue specimens were stained with hematoxylin and eosin (H&E) and microscopically examined (Olympus). Steatosis and inflammation were scored numerically following semiquantitative pathological standards. Histopathological changes in liver and ATs were quantitatively expressed simply by the summation of individual grades (score 0 to 5) for steatosis and inflammation, giving a maximum score of 25 per section as follows: 0 = normal; 1 = minimal; 2 = slight; 3 = moderate; 4 = marked; and 5 = severe. 18
Three different parts of each slide of ATs were randomly selected, and the diameter of 30 adipocytes was measured using a camera (TDI Digicam HQ) equipped with an image analyzer (Tomoro Scope Eye 3.6; Techsan). All these histological evaluating procedures were performed by a board-certified toxicological pathologist in a blind manner.
Semiquantitative reverse transcription-polymerase chain reaction
The total RNA from epididymal and subcutaneous adipocyte tissue was extracted using the TRIzol reagent (Invitrogen). For synthesis of cDNA, the isolated RNA (10 μg) was reverse transcribed with a transcriptase (Moloney murine leukemia virus) and Oligo(dT) 15 primers. Genes for differentiation of adipocytes in cDNA were amplified using PCR Primers, as shown in Table 1. A quantitative reverse transcription-polymerase chain reaction (RT-PCR) was performed using the MJ Mini Gradient Thermal Cycler (Bio-Rad). PCR mixture was denatured at 95°C for 30 sec, annealed for 30 sec, and elongated at 72°C (total 30 cycles). PCR products were separated in 1% agarose gel containing 0.5 μg/mL of ethidium bromide. GAPDH gene product was used as a control.
aP2, adipocyte protein 2 (NM_024406.2); C/EBPα, CCAAT/enhancer binding protein α (NM_001287514.1); FAS, fatty acid synthase (NM_008509.2); GAPDH, glyceraldehyde 3-phosphate dehydrogenase (NM_001289726.1); LPL, lipoprotein lipase (NM_007988.3); PPARγ, peroxisome proliferator-activated receptor-γ (NM_001127330.1); RT-PCR, reverse transcription-polymerase chain reaction; SREBP-1c, sterol regulatory element-binding protein-1c (NM_011480.4).
Statistical analysis
All research data were presented as mean standard error of the mean and were analyzed by one-way analysis of variance using SPSS Version 22 (IBM corp.). Statistical differences between the control and treated groups were analyzed using Tukey's multiple comparison test. A difference was considered statistically significant when the P value was .05 or less.
Results
Changes in body weight and FER of mice fed by HFD and SPE
Before feeding diets, there were no differences in body weights among the groups of mice. After 8 weeks of feeding the diets, the body weight of HFD was increased as much as ∼12.6 ± 1.1 g (320%), compared to increased body weight (3.0 ± 0.4 g) of the normal diet group (chow) (P < .001). On the contrary, the Orlistat positive control and SPE 200 and 400 groups showed significant increases by ∼10.2 ± 1.1 g (81% control), 10.7 ± 1.5 g (84.9% control), and 10.2 ± 1.4 g (81% control) in body weight, respectively, compared to weight increases of HFD mice (Table 2).
The values are presented as mean ± SEM (n = 8–10).
FER (%), feed efficiency ratio = body weight change (g)/food intake (g) × 100.
P < .05, *** P < .001 versus the chow group.
P < .05, ## P < .01, ### P < .001 versus the HFD group.
HFD, high-fat diet; SEM, standard error of the mean; SPE, Salvia plebeia extract.
No significant difference in food intake was observed in Chow-, HFD-, and SPE-treated groups. FER of HFD increased compared to that of Chow in 2–8 weeks (P < .001). However, FER of SPE 200 and SPE 400 had ∼23% and 31% decrease, compared to HFD, respectively (P < .05) (Table 2). Taken together, the antiobesity effect of SPE is supposed to be due to the decrease in FER.
Changes in weights of liver and ATs of mice fed HFD and SPE
After feeding experimental diets for 8 weeks, the distribution and amounts of AT were examined. In addition, liver and epididymal, subcutaneous, mesenteric, retroperitoneal, and kidney ATs were isolated and weighed (Table 3). Total amounts of fat were calculated from a sum of each organ's AT weight. Liver weight of the HFD group increased as much as 1.15 ± 0.05 g, while the increased weight of liver in Chow was ∼0.99 ± 0.06 g (P < .05). Increase of liver weight in the HFD group might be due to accumulation of fat in the liver. Otherwise, liver weights in Orlistat and SPE 200 and 400 groups were 1.01 ± 0.12, 0.97 ± 0.07, and 0.96 ± 0.09 g, which were significantly less than in the HFD group. Ratios of liver weight against whole body weight in Orlistat and SPE 200 and 400 groups decreased significantly in comparison to that of the HFD group (P < .001).
The values are presented as mean ± SEM (n = 8–10).
P < .05, *** P < .001 versus the chow group.
P < .05, ## P < .01, ### P < .001 versus the HFD group.
Weights of each AT in the HFD group increased significantly, compared to the Chow group (P < .001) (Table 3). In the SPE 200 and 400 groups, the fat weights in each tissue declined than the HFD group. In addition, the relative fat weight of HFD was increased than Chow, as similar to total fat weight change (P < .001). Epididymal AT in SPE 200 group and epididymal AT (P < .001), subcutaneous AT (P < .01), and mesenteric AT (P < .01) in SPE 400 group were less than in the HFD group. Taken together, SPE protected against the increase of body fat during high-fat diet. In Orlistat and SPE 400 groups, the relative weight ratios of subcutaneous and epididymal AT decreased more than the ratio in the HFD group (P < .05).
Enzyme activity related to liver function in mice fed by HFD and SPE
Levels of AST in SPE 200 and 400 groups were ∼139.0 ± 38.4 and 124.0 ± 45.0 IU/L, respectively, while levels of AST in Chow and HFS were 71.0 ± 7.0 and 164.0 ± 28.3 IU/L, respectively (Table 4). AST level of HFD mice increased more than that of Chow (P < .001), otherwise AST levels of Orlistat and SPE groups were significantly lower compared to the HFD group.
Chow: normal-diet group, bHFD: high-fat diet group.
The values are presented as mean ± SEM (n = 8–10).
Results were statistically analyzed using the Tukey's multiple comparison test, significantly different *** P < .001 versus the chow group, # P < .05, ## P < .01, ### P < .001 versus the HFD group.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; HDL-c, high-density lipoprotein cholesterol; TC, total cholesterol; TG, triglyceride; VLDL-c, very low-density lipoprotein cholesterol.
ALT activity in Chow was ∼22.3 ± 3.4 IU/L and in HFD was increased to 59.0 ± 14.9 IU/L in a significant difference. SPE 200 and 400 groups showed ∼43.6 ± 9.8 and 30.1 ± 13.8 IU/L of ALT, which were also lower than HFD (P < .001). Decreased ALT levels in the SPE groups are anticipated to be caused by the protective activity of SPE against hepatocyte damages by HFD.
Changes in serum TG and cholesterol in mice fed by HFD and SPE
TG levels (103.7 ± 10.7 mg/dL) in HFD were higher than that (94.8 ± 5.6 mg/dL) in the Chow group (P < .01) (Table 4). The mean TG level in the SPE 400 group was 102.7 ± 10.8 mg/dL, which was less than that in HFD (P < .01).
TC concentrations in serum were shown to be 131.8 ± 2.1 and 218.7 ± 15.8 mg/dL in Chow and HFD, respectively (P < .001). However, SPE 200 and 400 groups had ∼217.8 ± 5.9 and 198.8 ± 16.8 mg/dL of TC content, respectively. Especially, TC content in SPE 400 group was lower than that in the HFD group (P < .05).
Changes in leptin, adiponectin, and glucose levels in mice fed HFD and SPE
Mean serum leptin levels were 7.71 ± 0.556 and 47.29 ± 9.24 ng/mL in Chow and HFD, respectively (Table 4). HFD group had a higher level of leptin in serum than Chow (P < .001). SPE 200 and 400 groups had 49.75 ± 6.30 and 32.08 ± 11.73 ng/mL of leptin, respectively. Although the positive control agent, Orlistat, did not affect the level of leptin, SPE 400 mg/kg caused a decrease in serum leptin levels (32.2%) in mice fed HFD (P < .001). Otherwise, the adiponectin level in SPE groups did not show the increase, compared to Chow and HFD. Glucose levels in serum showed 162.3 ± 2.1 and 220.9 ± 10.3 mg/dL in the Chow and HFD group, respectively (P < .001). Blood glucose levels in Orlistat and SPE were significantly lower compared to HFD (P < .001).
Histological changes in liver and ATs in mice fed by HFD and SPE
Liver and adipocyte tissues fixed with formalin were examined visibly through staining with H&E. As shown in Figure 1A, a lot of lipid droplets were produced in the liver tissue of the HFD group, but the formation of lipid droplets was significantly suppressed by treatment with SPE 400 mg/kg. Liver tissue in HFD had significantly more steatosis and inflammatory cell infiltration (Fig. 1B, C) than in the Chow group (P < .001). However, the steatosis and inflammatory cell infiltration in SPE 400 was significantly compared to the HFD (P < .05).
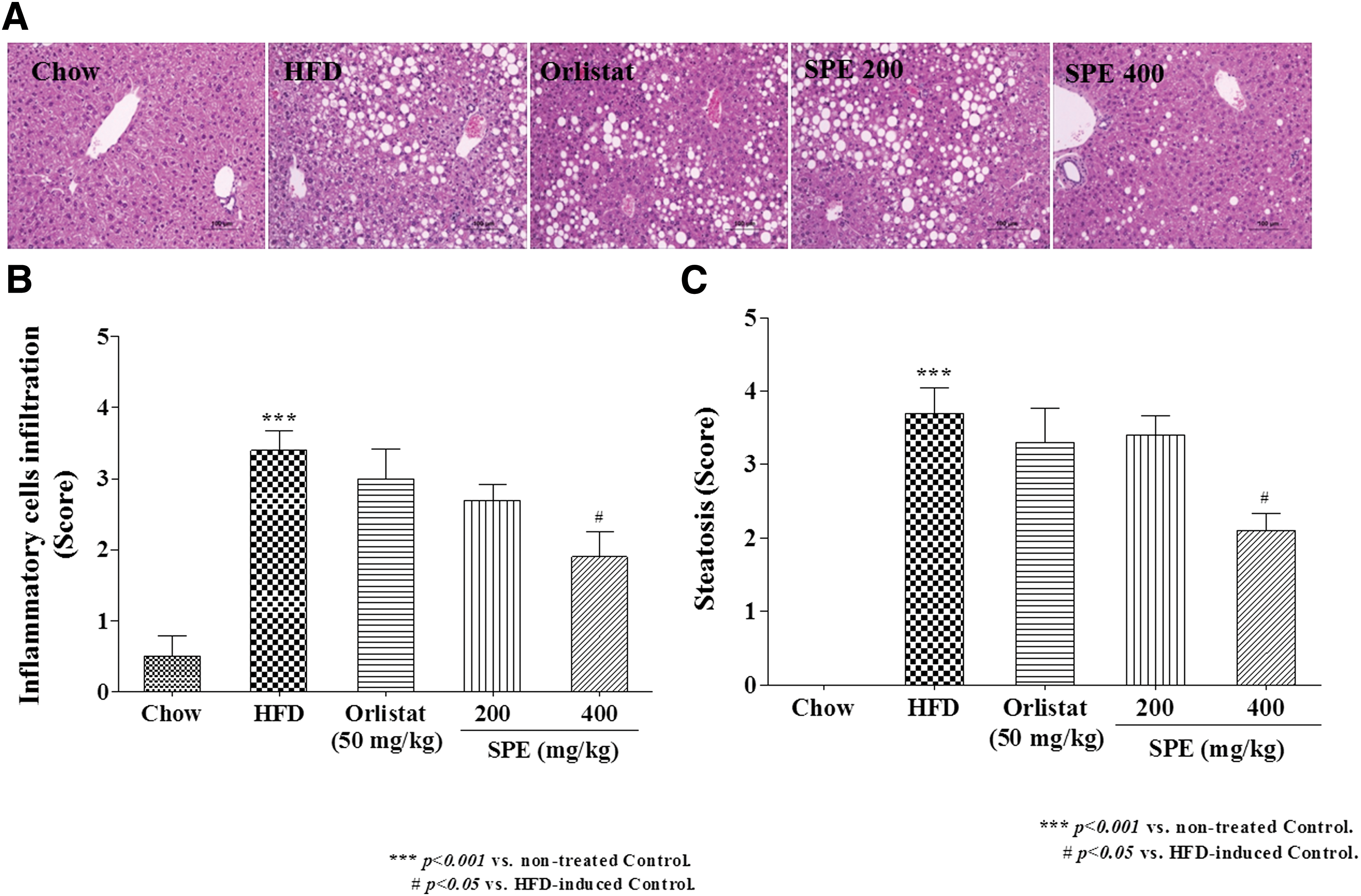
SPE induced histological change in liver of HFD-induced obese mice.
Generally, the sizes of lipid droplets in adipocyte tissue are significantly increased in obesity. 19 Diameter of adipocytes in subcutaneous AT was significantly smaller in the Orlistat and SPE groups (Fig. 2A, B). Adipocytes size in epididymal AT also decreased in SPE 400 group, compared with the HFD group (Fig. 2A, C) (P < .05).

Adipocyte diameter of adipose tissue in high-fat diet-induced obese mice.
Transcriptional changes in genes related to obesity in mice fed HFD and SPE
Epididymal AT and subcutaneous AT were removed from mice with obesity induced by feeding HFD. RT-PCR was performed for evaluating the transcriptional level of several genes related to obesity in removed tissues. Effects of SPE on transcriptional change of target genes and transcription factors related to obesity in tissue were investigated, as shown in Figure 3.
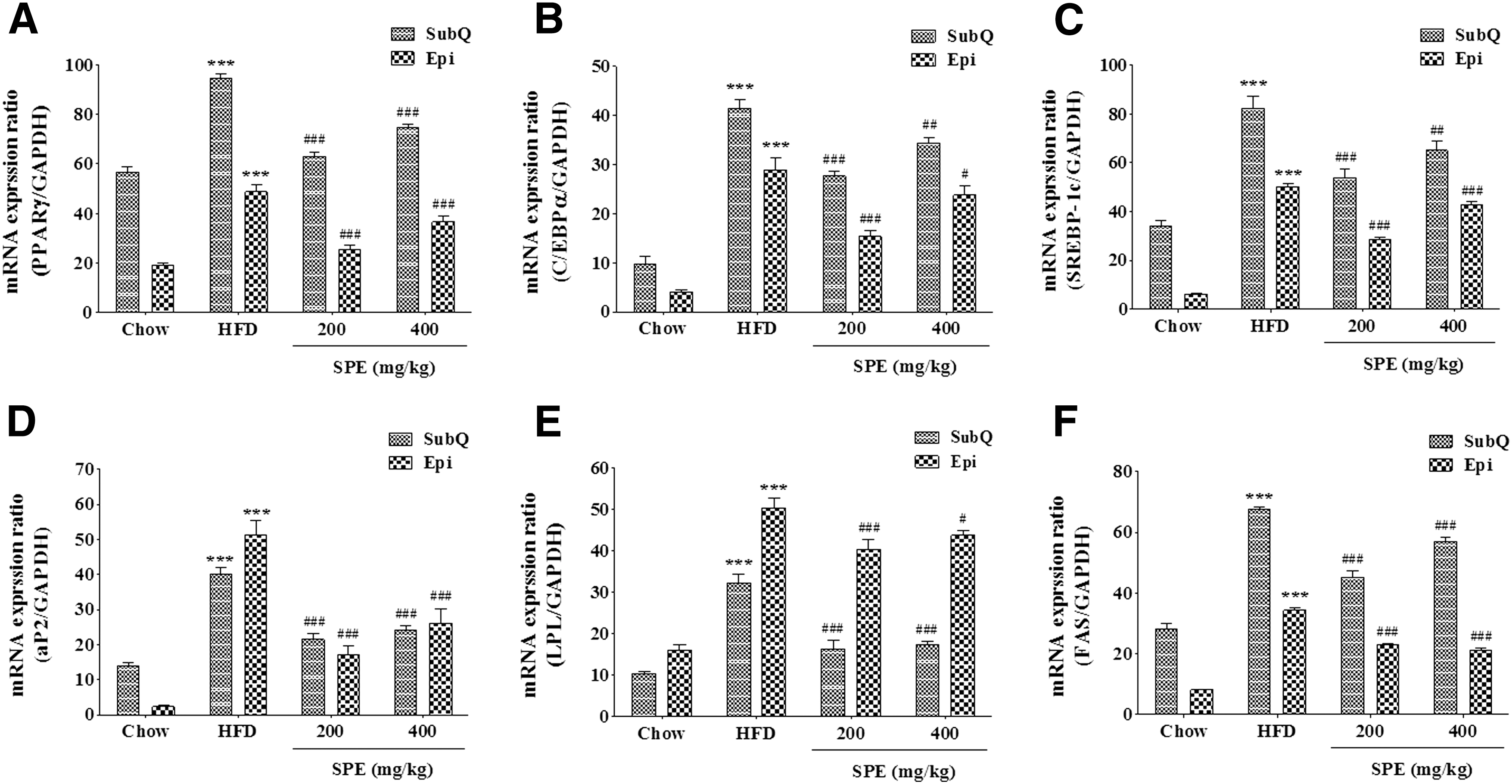
Effects of SPE on expression of transcriptional factors and adipocyte-specific genes in high-fat diet-induced obese mice. Expression of PPARγ
Level of transcription for all genes and transcription factors related to lipogenesis in HFD were increased (P < .001), compared to Chow. SPE treatment suppressed mRNA transcription of peroxisome proliferator-activated receptor (PPAR)γ and CCAAT/enhancer binding protein (C/EBPα) (P < .001). In the adipocyte tissue of the SPE group, adipocyte fatty acid-binding protein 2 (aP2), lipoprotein lipase (LPL), and fatty acid synthase (FAS) genes, which are target genes for obesity, were significantly inhibited transcriptionally by SPE treatment (P < .001). Transcription of the sterol regulatory element-binding transcription factor 1c (SREBP-1c) gene was inhibited by treatment with SPE 200 and SPE 400 mg/kg. Especially, SPE showed >45% inhibitory activity on transcription of aP2 and LPL.
In epididymal AT, the transcription of target genes and transcription factors were induced in HFD, compared to Chow (P < .001), and the treatment with SPE significantly suppressed the induction of mRNA transcription of PPARγ and C/EBPα genes in tissue. In addition, aP2, LPL, FAS, and SREBP-1c gene showed transcriptional inhibition in SPE-treated groups. Especially, transcriptions of aP2 and FAS gene were suppressed by >40% by SPE.
Therefore, SPE was anticipated to reduce the accumulation of TG in adipocyte tissues through transcriptional inhibition of C/EBPα and PPARγ (transcription factor) and aP2, LPL, FAS, and SREBP-1c gene (target gene).
Discussion
Obesity is a metabolic disease characterized by accumulation of body fat and an imbalance between energy intake and expenditure. Recently, the increase in obesity rates in the population is a serious social problem worldwide. However, little is known about the antiobesity activity of Salvia plebeian R. Br.
To evaluate the protective effect of SPE against obesity, the HFD group was fed a diet containing 45% (w/w) fat for 8 weeks. Experimental-treated groups (SPE 200 and 400) were orally administered 200 or 400 mg/kg of 50% SPE with the high-fat diet for 8 weeks and were monitored for changes of body weight, food intake, and feeding efficiency. Consequently, the body weight of HFD increased significantly more than Chow, resulting in obesity caused by the high-fat diet.
Body weights in Orlistat and SPE 200 and 400 groups appeared not to be significantly different against HFD for the first 3 weeks, but were significantly lower by 8 weeks in the SPE 400 group. Body weight gain in Orlistat and SPE 200 and 400 groups was ∼19.0%, 15.1%, and 19.0% less compared to that in HFD.
Positive control agent Orlistat, an inhibitor of lipase, has been used as a medicine for obesity to reduce body weight, due to disruption of the absorption of lipids in the gastrointestinal tract. Activity of Orlistat on reduction of body weight is caused from inhibition of lipid absorption into the body by decreased pancreatic lipase function and induction of and increased fecal excretion of fats. The side effects of Orlistat have been well known to be disturbances in defecation, frequent gas, fat defecation, fecal incontinence, and abdominal distension. 20
Orlistat and SPE 200 and 400 groups did not significantly affect food intake. On the contrary, the feed efficiency and body weight increased much more than other groups and were significantly different. Increased body weight in HFD and concomitant obesity are supposed to be caused by an imbalance of energy control and high-fat diet intake in the body. On the contrary, the SPE-treated group had significantly less feed efficiency than HFD. This is believed to be caused by decreased food digestion/absorption and nutrient bioavailability and increased excretion of fat by SPE treatment. Taken together, mice supplemented with SPE exhibited a regulatory effect of obesity through activities related to decreased feed efficiency.
GOT/AST and GPT/ALT exist abundantly in liver and are released into the blood in acute/chronic hepatitis, fatty liver, alcoholic hepatitis, and liver cancer. Therefore, the levels of AST and ALT in blood are commonly used as an index for denaturation and death of hepatocytes. In the fatty liver, alcoholism, and overweight; the inflammation in the liver and damages of hepatocytes increase AST and ALT in blood. 21 HFD had higher AST and ALT values, which are an index for hepatotoxicity and fatty liver, than Chow, but AST and ALT concentrations in HFD were decreased by SPE treatment.
Cholesterol and TG in serum and tissues are increased in obesity induced by diets and are closely related to the level of obesity. In addition, the chronically high levels of cholesterol and TG cause hypercholesterolemia and hyperlipidemia. 22 TG, Total-cholesterol (c), and very low-density lipoprotein-c, which were indexes for obesity, in HFD had higher values than Chow and were significantly decreased by treatment of 400 mg/kg SPE.
Circulating concentrations of leptin, a hormone produced in adipocytes, were reduced by SPE treatment. Production of leptin is stimulated by overaccumulation of energy in the body. The role of leptin is to reduce the energy intake by decreasing appetite due to signal transfer to the hypothalamus. Leptin is used as an obesity index to regulate body weight and energy balance by decreasing fat storage and energy consumption and therefore is closely related to body weight and body fat formulation. 23,24 Diet-induced obese mice have increased concentrations of leptin in blood, which are proportionate to the amount of fat in the body, 25 and the long period hyperinsulinemia in obesity affects the leptin levels in blood. 26 SPE treatment decreased leptin concentrations in serum, body weight, and decreased fat accumulation in the tissues, all of which were associated with decreases in body weight and body fat by SPE.
Generally, increase in body weight causes various secondary diseases, such as diabetes, cardiac disorders, and hypertension. 27,28 In this study, the high-fat diet mice had high blood glucose levels, compared to the normal diet mice, which are consistent with increases in blood glucose by obesity across species. Treatment with SPE significantly suppressed the level of blood glucose in the HFD group mice.
SPE treatment also resulted in mRNA transcriptional change of genes related to obesity in liver tissue, epididymal AT, and subcutaneous AT. PPARγ and C/EBPα mRNA transcriptions were inhibited significantly in liver and fat tissue of the SPE-treated group. In addition, mRNA transcription of aP2, LPL, FAS, SREBP-1c, and HSL gene suppressed by SPE treatment. PPARγ is distributed mostly in ATs, where it regulates the formation of fat in cells. PPARγ differentiates fat tissues better than other transcription factors and is essential to differentiation of cells. C/EBPα induces the sensitivity to insulin in preadipocyte cells and is important in fat formation, and the latter process of differentiation to adipocyte with PPARγ. C/EBPα is a regulatory factor for transcription of genes related to differentiation. 29
Expression of C/EBPα and PPARγ is stimulated in the latter process of differentiation of preadipocytes to adipocyte and is involved in progression of differentiation. Differentiated cells have cytomorphological features, such as increases in cell size and accumulation of TG in the cell. C/EBPα and PPARγ increase the transcription of aP2, FAS, HSL, SREBP-1c, and HSL, which are subfactors of transcription and biomarkers for adipogenesis, and provide features of adipocytes. 30 In particular, FAS is involved in metabolism of lipids, is a final factor in differentiation of adipocytes, and is expressed most abundantly in adipocyte tissues.
In addition, LPL functions as a transfer enzyme for blood fatty acid into the cells. Fatty acids are converted to TG and accumulate in cells due to the action of LPL in fat tissues. Suppression of LPL transcription and activity inhibits the absorption of undecomposed TG in lipoprotein into the body, which results in decreased energy supply and reduced body weight. 31
In conclusion, the obesity in mice (C57BL/6N) was induced by feeding high-fat diets and was treated by SPE for 8 weeks. Body weight in the HFD group increased significantly compared to the Chow group; SPE-treated groups gained less body weight and had significantly less mesenteric AT and subcutaneous AT than HFD group at the end of the 8 weeks. The SPE group had significantly less enzyme activities or expressions for liver function, TG, and leptin level than the HFD group. Expression of the SREBP-1c gene, which is involved in synthesis of fat and cholesterol, was significantly inhibited in liver and fat tissues of the SPE-treated group compared to the HFD group. Taken together, SPE partially prevented the increase of body weight in a high-fat diet-induced obese mouse model and might be used as a potential antiobesity food material.
Footnotes
Acknowledgments
This research was supported by High Value-added Food Technology Development Program, Ministry of Agriculture, Food and Rural Affairs.
Author Disclosure Statement
No competing financial interests exist.