
Abstract
This study investigated the effects of ethanol extract from Gynostemma pentaphyllum (GP-EX) on memory deficits in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-lesioned mouse model of Parkinson's disease (PD) (MPTP-lesioned mice). MPTP (30 mg/kg/day, 5 days)-lesioned mice showed deficits of habit learning memory and spatial memory, which were further aggravated by treatment with L-3,4-dihydroxyphenylalanine (L-DOPA) (25 mg/kg, 21 days). However, treatment with GP-EX (50 mg/kg, 21 days) ameliorated memory deficits in MPTP-lesioned mice treated with L-DOPA (25 mg/kg): GP-EX prevented the decreases in retention latency time in the passive avoidance test and tyrosine hydroxylase-immunopositive cells and dopamine levels in the nigrostriatum. GP-EX also reduced increases in retention transfer latency time of the elevated plus-maze test and expression of N-methyl-D-aspartate (NMDA) receptor and improved decreases in phosphorylation of extracellular signal-regulated kinase (ERK1/2) and cyclic AMP-response element binding protein (CREB) in the hippocampus in the same models. By contrast, L-DOPA treatment (10 mg/kg, 21 days) ameliorated memory deficits in MPTP-lesioned mice, which were further improved by GP-EX treatment. These results suggest that GP-EX ameliorates habit learning memory deficits by activating dopaminergic neurons and spatial memory deficits by modulating NMDA receptor-ERK1/2-CREB system in MPTP-lesioned mice treated with L-DOPA. GP-EX may serve as an adjuvant phytonutrient for memory deficits in PD.
Introduction
P
The dopaminergic neurons in the substantia nigra and striatum are involved in habit learning memory in humans and animals. 5 –7 L-DOPA treatment in early PD may improve certain cognitive functions. 7 By contrast, L-DOPA administration enhances verbal learning and high-level cognitive deficits in patients with PD. 8,9 N-Methyl-D-aspartate (NMDA) receptors of the CA1 hippocampal region are also involved in learning and memory, including spatial memory, in human and rats. 10 –13 In addition, the extracellular signal-regulated kinase (ERK1/2) and cyclic AMP-response element binding protein (CREB) are an essential component of NMDA receptor signaling transduction-mediated spatial memory in the hippocampus. 14,15
Gynostemma pentaphyllum (Cucurbitaceae) has been used as a herbal tea and contains various bioactive gypenoside derivatives and flavonoids. 16 Recently, the ethanol extract from G. pentaphyllum (GP-EX) has been shown to exert protective effects against neurotoxicity by reducing tyrosine hydroxylase (TH) neuronal cell death and L-DOPA-induced dyskinesia in 6-hydroxydopamine (6-OHDA)-lesioned rat model of PD. 17,18 GP-EX also shows anxiolytic effects on affective disorders and neuroprotective effects in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-lesioned mouse model of PD. 19
This study, therefore, investigated the effects of GP-EX on memory deficits in the MPTP-lesioned mouse model of PD treated with L-DOPA by examining the behavioral changes of the latency time and the biochemical influences on the dopaminergic neuronal system and the NMDA receptor system, including ERK1/2 and CREB.
Materials and Methods
GP-EX preparation
G. pentaphyllum was obtained from the Wonkwang Food Manufacturing Co. (Geochang, Korea), and a voucher specimen of the herbal leaves of G. pentaphyllum was deposited at the herbarium of the College of Pharmacy, Chungbuk National University (Cheongju, Korea). The air-dried leaves of G. pentaphyllum (1 kg) were extracted with ethanol (80%, v/v), and the ethanol extracts were evaporated to dryness under reduced pressure and temperature (GP-EX, 95.3 g, yield, 9.5%, w/w). 17
Experimental design
Mice (C57BL/6, male, 20–25 g) were obtained from Samtako Co. (Animal Breeding Center, Osan, Korea) and maintained as described previously. 18,19 All procedures were approved by the Animal Ethics Committee of Chungbuk National University (Approval No. CBNUA-872-15-02), and the experiments were performed in accordance with the guidelines of the NIH for Care and Use of Laboratory Animals and Chungbuk National University Laboratory Animal Research Center.
The experiments were performed in two separate subsets of six groups (8–10 animals/group), including (1) the step-through passive avoidance test and TH-immunohistochemical analysis and (2) the elevated plus-maze test, including biochemical and Western blot analyses. The control group received 0.9% saline. The MPTP-lesioned group (MPTP) was injected with MPTP (30 mg/kg/day, daily for 5 days, intraperitoneal [i.p.]). 20 The L-DOPA-treated group (MPTP+L-DOPA) was treated with L-DOPA (10 or 25 mg/kg with benserazide, 15 mg/kg, i.p.) daily for 21 days after the 5 days of MPTP injections, and the GP-EX-treated group (MPTP+GP-EX+L-DOPA) received GP-EX (50 mg/kg, p.o.) daily for 21 days 3 h before L-DOPA treatment in the L-DOPA-treated group. After the final treatments, all mice were subjected to behavioral tests. The mice were then anesthetized (Zoletil 50, 100 mg/kg, i.p.; Virbac, Carros, France) and sacrificed to obtain brain tissues for TH-immunohistochemical, biochemical, and Western blot analyses.
Latency time in the step-through passive avoidance and elevated plus-maze tests
After the final treatments, all mice were subjected to behavioral tests. Each mouse was placed in the illuminated chamber on the first day after the habituation periods using the step-through passive avoidance apparatus (Med Associates, Inc., St. Albans City, Vermont, USA). An inescapable electric shock (0.5 mA, 3 sec) was delivered to mice. The initial latency time of entrance into the dark chamber was recorded, and the initial latency times >180 sec were excluded from the experiment. Twenty-four hours later, each mouse was placed in the illuminated chamber to measure the retention latency time. 21
The elevated plus-maze apparatus consisted of four arms (30 × 5 cm): two open and two closed arms of the same size, with 16-cm-high black walls elevated 45 cm. Each mouse was placed on the open arm facing outward. The time to enter the closed arm in the first trial was noted as initial transfer latency. A second trial was performed 24 h after the first trial and measured for retention transfer latency. The results are expressed as the ratio of the retention transfer latency time to initial transfer latency time (%ITL). 22
TH immunohistochemistry
Mice were intracardially perfused with a paraformaldehyde solution (4% in 0.1 M phosphate buffered saline, pH 7.4). Coronal brain sections (30 μm) were made through the cell bodies of dopaminergic neurons of the substantia nigra (Vibratome; Leica Microsystems GmbH, Wetzlar, Germany). The sections were processed for TH-immunohistochemical staining using TH primary antibody (1:200, in 0.3% Triton X-100; Chemicon Int., Temecula, CA, USA) overnight at 4°C and biotinylated goat anti-rabbit secondary antibody (1:250; Vector Laboratories, Burlingame, CA, USA). Photomicrographs of TH immunoreactivity and digitized bright-field images and the TH-immunopositive cell counting were performed as described previously. 19
Dopamine levels
The striatum tissue samples of mice were homogenized in 300 μL of 0.1 M perchloric acid containing 100 mM EDTA and 10 μM isoproterenol and then centrifuged (13,000 g, 15 min, 4°C). The supernatant was passed through a filter (Millex-GV, 0.45 μm, Waters), and the filtrate (100 μL) was injected into an high-performance liquid chromatography (HPLC) system with an electrochemical detector (+0.85 V, Ag/AgCl reference electrode; Model 2465; Waters) and a Waters 120 ODS-BP column (5 μm, 50 × 4.6 mm) as described previously. 19 The dopamine levels were expressed as a percentage of the control group.
Western blot analysis
The phosphorylation of NMDA receptor (Ser 897; Sigma-Aldrich, St. Louis, MO, USA), ERK1/2 (Thr 202/Tyr 204), CREB (Ser 133), and β-actin (Cell Signaling Tech, Danvers, MA, USA) was determined by western blot analysis. After the hippocampal tissue samples were homogenized and added the sample loading buffers, the blot analysis on proteins in samples (30 μg in each lane) was conducted using primary antibodies (1:1000 using TBS-T) at 4°C and secondary antibodies (1:5000 using TBS-T) as described previously. 19
Statistical analysis
The results are analyzed by one-way analysis of variance (ANOVA) followed by Tukey's test for evaluating the effects of GP-EX and two-way ANOVA followed by Tukey's test for analyzing the retention latency time in the passive avoidance test. The results are expressed as the mean ± standard error of the mean, with a P value <.05 being considered statistically significant.
Results
Latency time for habitual memory in the step-through passive avoidance test
The latency time in the step-through passive avoidance test is shown in Table 1. No overall baseline differences were observed between the control group, MPTP-lesioned group, and MPTP-lesioned group treated with L-DOPA or/and GP-EX (average of initial latency time from each group, 24.3 sec). The MPTP-lesioned group showed a significant impairment as a decreased retention latency time (decrease to 60.6 sec, P < .01, F = 31.7), compared with the control group (126.3 sec). The 10 mg/kg L-DOPA-treated group increased the retention latency time to 90.2 sec (P < .05, F = 23.8) compared with the MPTP-lesioned group, whereas the 25 mg/kg L-DOPA-treated group further decreased the retention latency time to 56.9 sec. By contrast, treatment with GP-EX (50 mg/kg) improved the retention latency time to 104.5 sec (P < .05, F = 16.2) and 83.9 (P < .05, F = 22.4), respectively, compared with the MPTP-lesioned group treated with L-DOPA (10 and 25 mg/kg).
Treatments with GP-EX (50 mg/kg) and L-DOPA (10 and 25 mg/kg) in MPTP-lesioned mice and the behavioral test were performed according to the Materials and Methods section. The results are expressed as the mean ± SEM for 8–10 animals per group.
P < .01 compared with the control group; # P < .05 compared with the MPTP group; § P < .05 compared with the MPTP+L-DOPA (10 and 25 mg/kg) group, respectively (two-way ANOVA followed by Tukey's test).
ANOVA, analysis of variance; GP-EX, ethanol extract from Gynostemma pentaphyllum; L-DOPA, L-3,4-dihydroxyphenylalanine; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; SEM, standard error of the mean.
Transfer latency time for spatial memory in the elevated plus-maze test
The ratio of the retention transfer latency time to initial transfer latency time (%ITL) was significantly increased to 152.3% (P < .01) in the MPTP-lesioned group, compared with the control group (Fig. 1). Treatment with L-DOPA (10 mg/kg) in the MPTP-lesioned group decreased %ITL to 136.6% (P < .05) compared with the MPTP-lesioned group, however, treatment with L-DOPA (25 mg/kg) increased %ITL to 153.4% (Fig. 1). By contrast, treatment with GP-EX (50 mg/kg) in the MPTP-lesioned group treated with L-DOPA (10 and 25 mg/kg) decreased %ITL to 121.9% (P < .05) and 134.3% (P < .05), respectively, compared with the MPTP-lesioned group treated with each L-DOPA alone (Fig. 1).
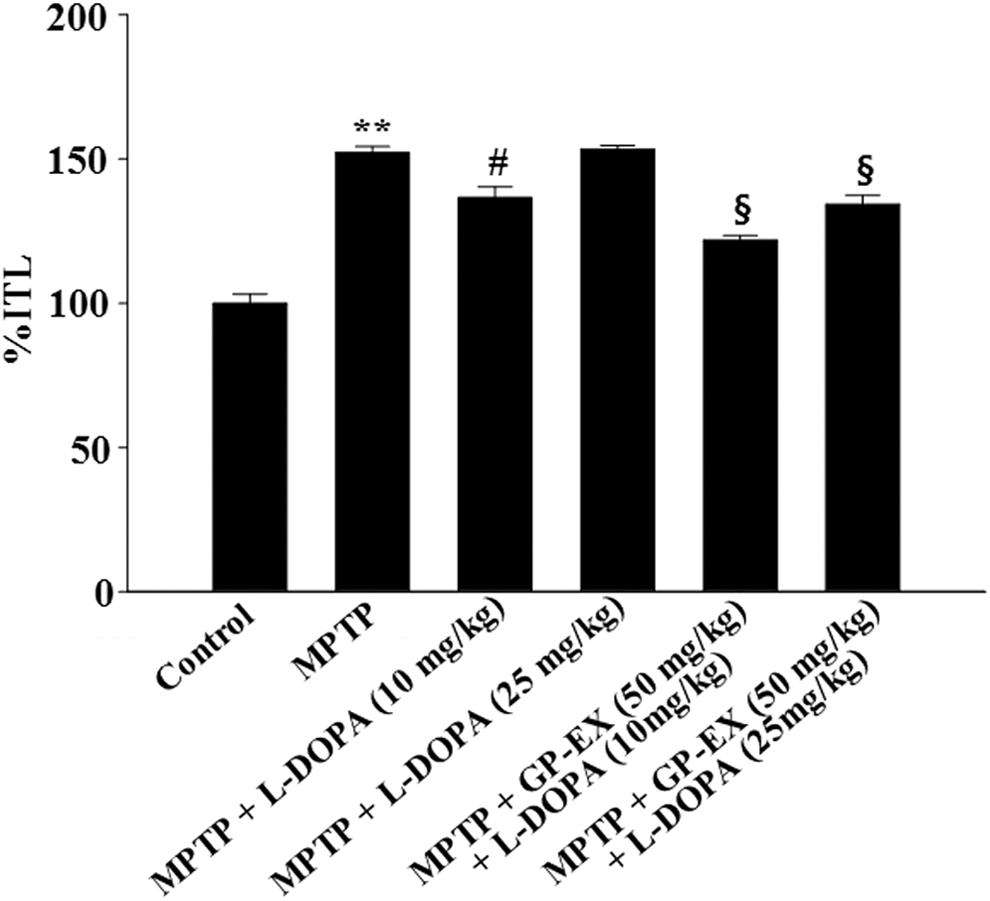
Effects of GP-EX on retention transfer latency time (%ITL) in the elevated plus-maze test. Treatments with GP-EX (50 mg/kg) and L-DOPA (10 and 25 mg/kg) in MPTP-lesioned mice were performed according to the Materials and Method section. The results are expressed as the ratio of the retention transfer latency time to initial transfer latency time (%ITL) and as the mean ± SEM for 8–10 animals per group. **P < .01 compared with the control group, # P < .05 compared with the MPTP group, § P < .05 compared with the MPTP+L-DOPA (10 and 25 mg/kg) group, respectively (one-way ANOVA followed by Tukey's test). ANOVA, analysis of variance; GP-EX, ethanol extract from Gynostemma pentaphyllum; L-DOPA, L-3,4-dihydroxyphenylalanine; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; SEM, standard error of the mean.
TH-immunopositive cells in the substantia nigra
Representative photomicrographs showed that TH-immunopositive cells were significantly reduced to 41.4% (P < .01) in the MPTP-lesioned group, compared with the control group (Fig. 2). The TH-immunopositive cells that were reduced by MPTP lesion were increased to 57.7% (P < .05) after treatment with 10 mg/kg L-DOPA compared with the MPTP-lesioned group, however, TH-immunopositive cells reduced by MPTP lesion were further decreased to 38.9% after treatment with 25 mg/kg L-DOPA (Fig. 2). By contrast, treatment with GP-EX (50 mg/kg) in the MPTP-lesioned group treated with L-DOPA (10 and 25 mg/kg) increased TH-immunopositive cells to 78.8% (P < .05) and 74.4% (P < .05), respectively, compared with the MPTP-lesioned group treated with L-DOPA alone (Fig. 2).
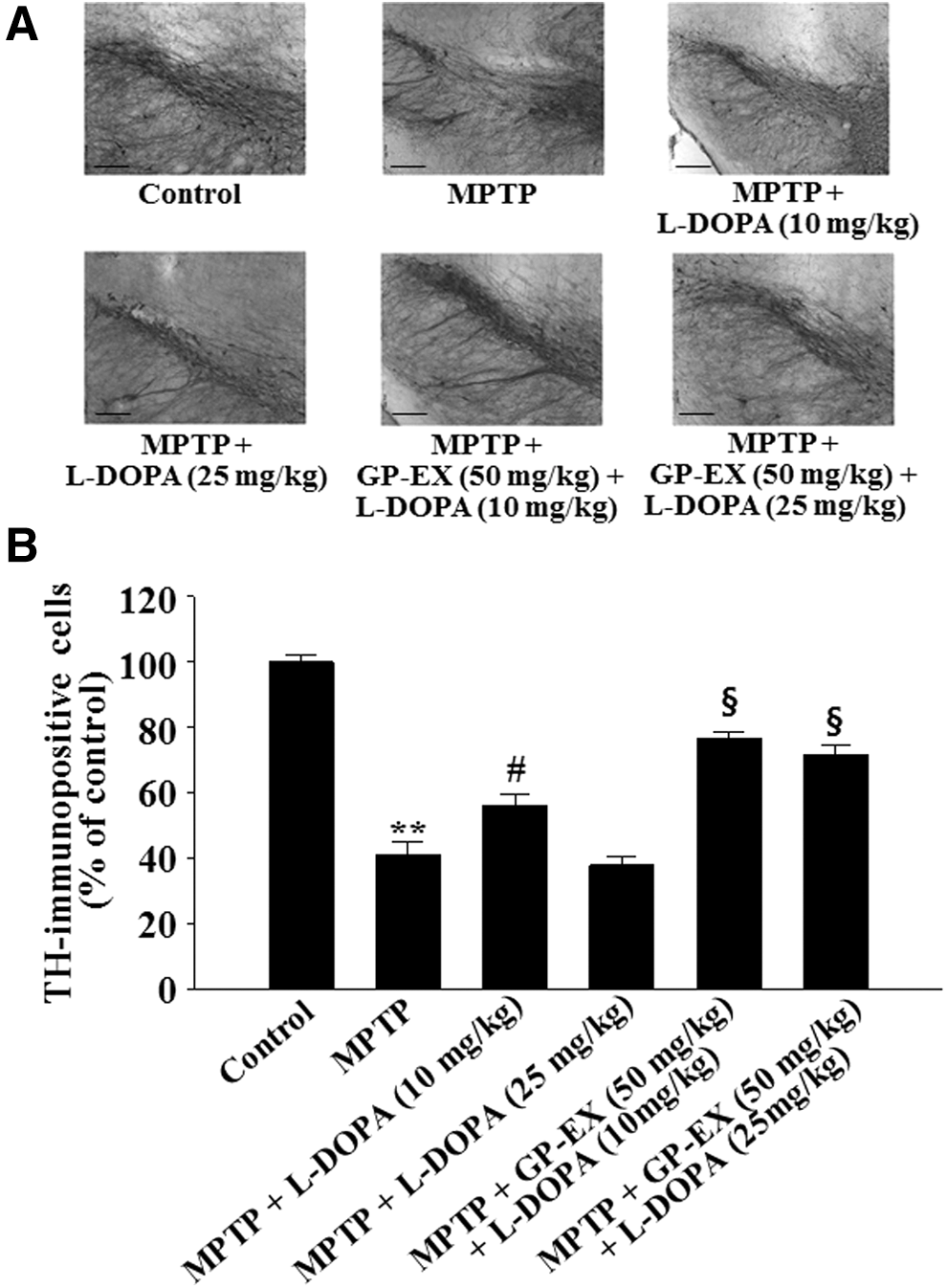
Representative photographs illustrating the effects of GP-EX on TH-immunohistochemistry
Dopamine levels in the striatum
The levels of dopamine were significantly decreased to 49.6% (P < .01) in the MPTP-lesioned group, compared with controls (Fig. 3). The decrease in dopamine levels by MPTP lesion was restored to 62.9% (P < .05) by treatment with 10 mg/kg L-DOPA, while the dopamine levels were further decreased to 47.7% by treatment with 25 mg/kg L-DOPA (Fig. 3). However, treatment with GP-EX (50 mg/kg) in the MPTP-lesioned group treated with L-DOPA (10 and 25 mg/kg) increased dopamine levels to 76.3% (P < .05) and 61.6% (P < .05), respectively, compared with the MPTP-lesioned group treated with each L-DOPA alone (Fig. 3).
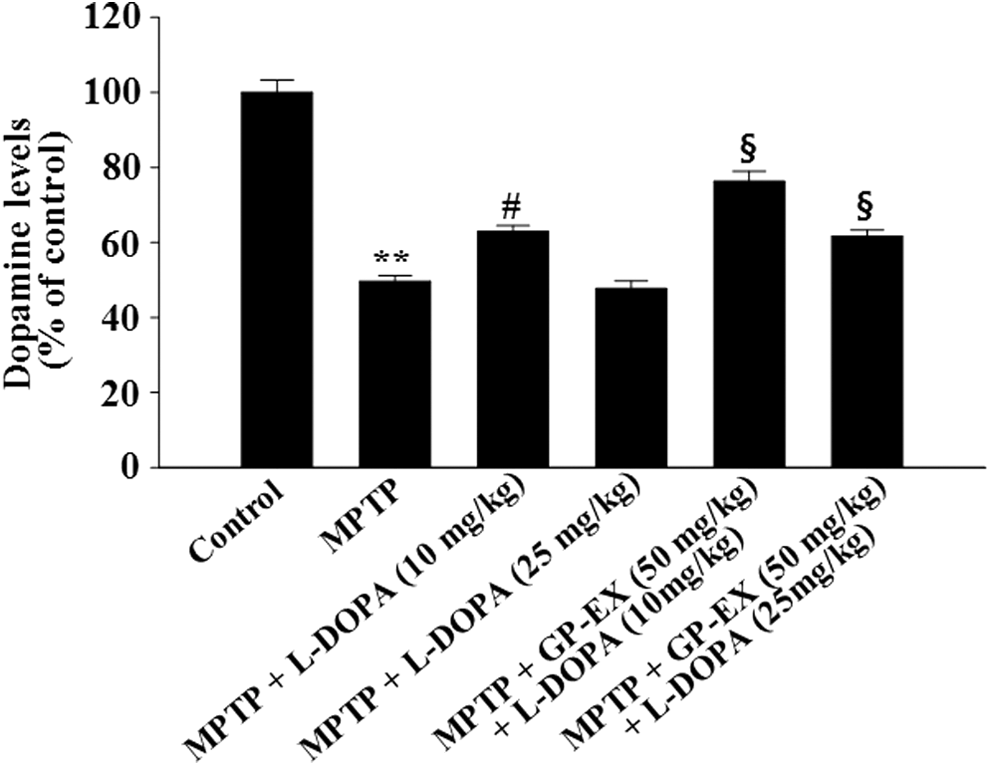
Effects of GP-EX on dopamine levels in the striatum. Treatments with GP-EX (50 mg/kg) and L-DOPA (10 and 25 mg/kg) in MPTP-lesioned mice were performed according to the Materials and Method section. The dopamine levels of control group were 8.38 ± 0.72 ng/mg tissue. The results were expressed as the mean ± SEM for 8–10 animals per group. **P < .01 compared with the control group, # P < .05 compared with the MPTP group, § P < .05 compared with the MPTP+L-DOPA (10 and 25 mg/kg) group, respectively (one-way ANOVA followed by Tukey's test).
Phosphorylation of NMDA receptor, ERK1/2, and CREB in the hippocampus
The phosphorylation of NMDA receptor (p-NMDAR1) was significantly induced to 1.65-fold (P < .01) by MPTP lesion, compared with the control group (Fig. 4). Treatment with 10 mg/kg L-DOPA in the MPTP-lesioned group reduced p-NMDAR1 to 1.22-fold over baseline (P < .05) compared with the MPTP-lesioned group, whereas treatment with 25 mg/kg L-DOPA increased p-NMDAR1 to 1.71-fold (Fig. 4). In addition, treatment with GP-EX (50 mg/kg) in the MPTP-lesioned group treated with L-DOPA (10 and 25 mg/kg) reduced p-NMDAR1 to 1.10-fold (P < .05) and 1.49-fold (P < .05), respectively, compared with the MPTP-lesioned group treated with L-DOPA alone (Fig. 4).
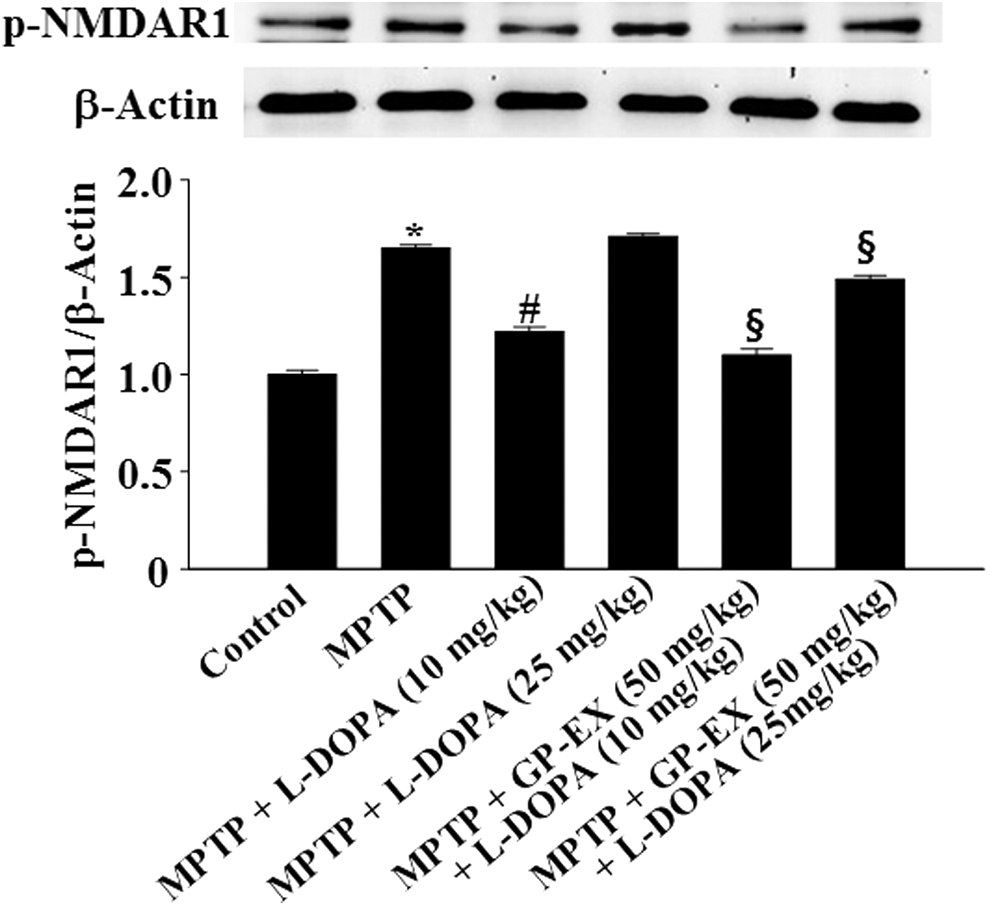
Effects of GP-EX on NMDA receptor (type 1) (NMDAR1) phosphorylation in the hippocampus. Treatments with GP-EX (50 mg/kg) and L-DOPA (10 and 25 mg/kg) in MPTP-lesioned mice were performed according to the Materials and Method section. The value of the relative density ratio of phosphorylation of NMDAR1 (p-NMDAR1)/β-actin is expressed in arbitrary units. The results were expressed as the mean ± SEM for 8–10 animals per group. *P < .05 compared with the control group, # P < .05 compared with the MPTP group, § P < .05 compared with the MPTP+L-DOPA (10 and 25 mg/kg) group, respectively (one-way ANOVA followed by Tukey's test). NMDA, N-methyl-D-aspartate.
p-ERK1/2 was reduced to 0.56-fold (P < .01) by MPTP lesion, compared with the control group (Fig. 5). Treatment with 10 mg/kg L-DOPA increased p-ERK1/2 to 0.80-fold (P < .05) compared with the MPTP-lesioned group, while treatment with 25 mg/kg L-DOPA in the MPTP-lesioned group reduced p-ERK1/2 to 0.52-fold (Fig. 5). However, treatment with GP-EX (50 mg/kg) in the MPTP-lesioned group treated with L-DOPA (10 and 25 mg/kg) increased p-ERK1/2 to 0.88-fold (P < .05) and 0.77-fold (P < .05), respectively, compared with the MPTP-lesioned group treated with L-DOPA alone (Fig. 5).
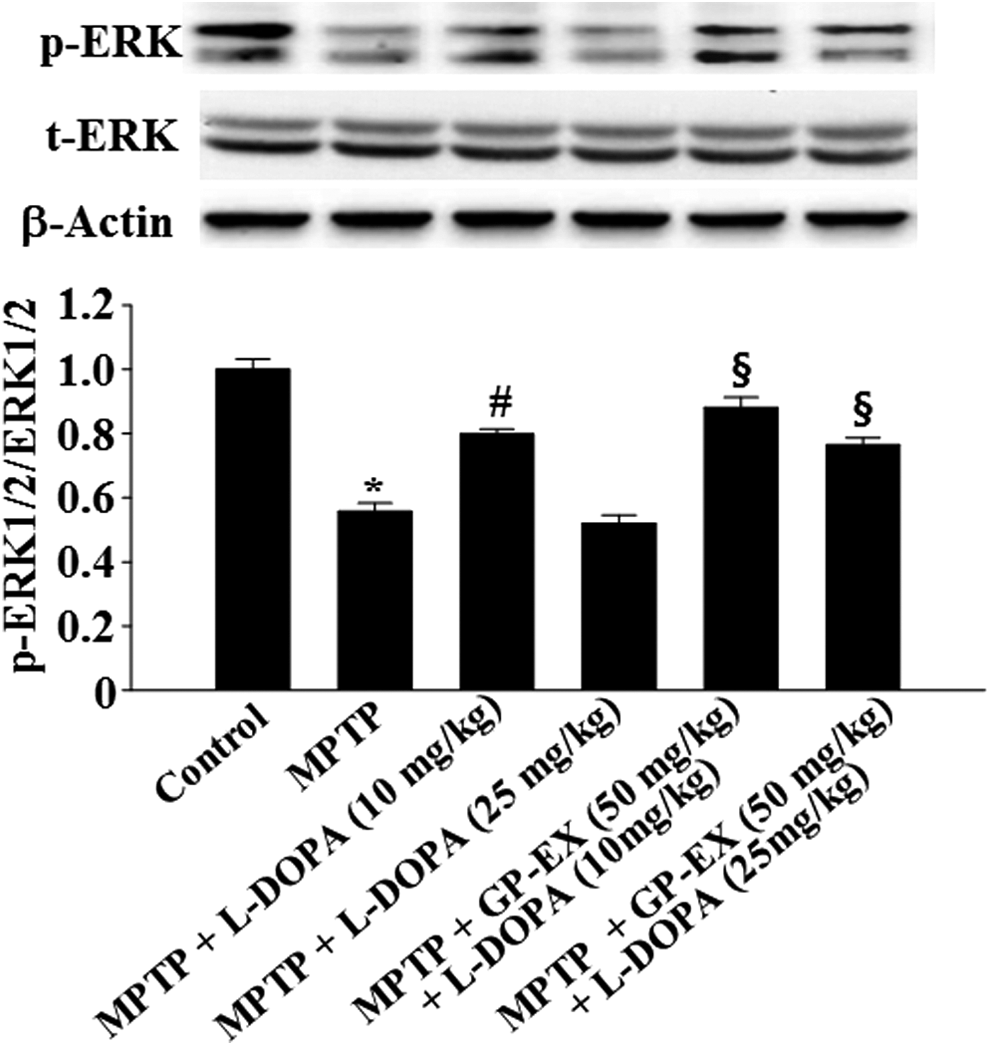
Effects of GP-EX on ERK1/2 phosphorylation in the hippocampus. Treatments with GP-EX (50 mg/kg) and L-DOPA (10 and 25 mg/kg) in MPTP-lesioned mice were performed according to the Materials and Method section. The value of the relative density ratio of phosphorylation of ERK1/2 (p-ERK1/2)/total ERK1/2 (t-ERK1/2) is expressed in arbitrary units. The results were expressed as the mean ± SEM for 8–10 animals per group. *P < .05 compared with the control group, # P < .05 compared with the MPTP group, § P < .05 compared with the MPTP+L-DOPA (10 and 25 mg/kg) group, respectively (one-way ANOVA followed by Tukey's test). ERK1/2, extracellular signal-regulated kinase.
p-CREB was decreased to 0.53-fold (P < .01) by MPTP lesion, compared with controls (Fig. 6). Treatment with 10 mg/kg L-DOPA in the MPTP-lesioned group increased p-CREB to 0.66-fold (P < .05) compared with the MPTP-lesioned group, but L-DOPA treatment (25 mg/kg) reduced p-CREB to 0.48-fold (Fig. 6). By contrast, treatment with GP-EX (50 mg/kg) in the MPTP-lesioned group treated with L-DOPA (10 and 25 mg/kg) increased p-CREB to 0.85-fold (P < .05) and 0.72-fold (P < .05), respectively, compared with the MPTP-lesioned group treated with each L-DOPA alone (Fig. 6).

Effects of GP-EX on CREB phosphorylation in the hippocampus. Treatments with GP-EX (50 mg/kg) and L-DOPA (10 and 25 mg/kg) in MPTP-lesioned mice were performed according to the Materials and Method section. The value of the relative density ratio of phosphorylation of CREB (p-CREB)/total CREB (t-CREB) is expressed in arbitrary units. The results were expressed as the mean ± SEM for 8–10 animals per group. *P < .05 compared with the control group, # P < .05 compared with the MPTP group, § P < .05 compared with the MPTP+L-DOPA (10 and 25 mg/kg) group, respectively (one-way ANOVA followed by Tukey's test). CREB, cyclic AMP-response element binding protein.
Discussion
GP-EX at 25 and 50 mg/kg ameliorated the effects of the 6-OHDA- and MPTP-lesioned animal model of PD. 17 –19 Recently, treatment with GP-EX (50 mg/kg) for 21 days in MPTP-lesioned mice improved the retention latency time to 60.1% (P < .05) and reduced the retention transfer latency time (%ITL) to 114.7% (P < .05), compared with the MPTP-lesioned group (data not shown). In this study, a dosage of 50 mg/kg of GP-EX was selected by considering the previous results. 17 –19 Treatment with GP-EX (50 mg/kg) in the control group did not alter the latency time in the passive avoidance and elevated plus-maze tests (data not shown).
The passive avoidance test is commonly used for examining habit learning memory deficits in MPTP-lesioned mice. 21 The elevated plus-maze test is used for anxiety testing by determining time spent on open arms and number of open arm entries. 23 The transfer latency time in the elevated plus-maze test is used to evaluate the parameters of spatial memory, 22,24 which is reported to have several advantages. 24 The elevated plus-maze method has also been applied to study spatial memory deficits. 22,25 We therefore used the passive avoidance and elevated plus-maze tests to evaluate the effects of GP-EX on habit learning memory and spatial memory deficits in MPTP-lesioned mice.
It has been demonstrated that MPTP injection causes dopaminergic neuronal degeneration in the substantia nigra and striatum, which induces deficits of learning and memory. 4,26,27 MPTP may also lead to neuroinflammation, including microglial activation and cell loss in the hippocampus of the rat model of PD. 28,29 In addition, chronic L-DOPA administration induces the dopamine receptor dysregulation in the hippocampus of rat brain. 30 In this study, the retention latency time in the passive avoidance test was decreased in MPTP-lesioned mice treated with L-DOPA (25 mg/kg), which was improved by treatment with GP-EX (50 mg/kg) (Table 1). Both TH-immunopositive cells and dopamine levels, which were decreased by 25 mg/kg L-DOPA treatment in MPTP-lesioned mice, were increased after treatment with GP-EX (50 mg/kg) (Figs. 2 and 3). These results suggest that GP-EX ameliorates deficits of habit learning memory by preventing dopaminergic neuronal cell death in the substantia nigra and striatum of MPTP-lesioned mice treated with L-DOPA (25 mg/kg).
Impairments of spatial working memory are induced by MPTP and 6-OHDA in rat models of PD. 31 NMDA receptor-mediated signaling transduction and ERK1/2 and CREB components in the NMDA receptor play a key role in learning and memory, including spatial memory in the hippocampus. 10 –15 By contrast, excessive activation of NMDA receptors leads to cell death due to calcium overload in acute central nervous system (CNS) injury syndromes such as ischemia and trauma. 32 NMDA receptor expression is also increased in the striatal region of 6-OHDA-lesioned rats treated with L-DOPA (25 mg/kg). 33 In this study, %ITL was significantly increased by MPTP lesion, which was further aggravated by treatment with L-DOPA at 25 mg/kg (Fig. 1). However, treatment with GP-EX (50 mg/kg) decreased %ITL in the L-DOPA (25 mg/kg)-treated MPTP-lesioned group (Fig. 1). Furthermore, GP-EX (50 mg/kg) suppressed the increased NMDA receptor expression and reversed the suppression of ERK1/2 and CREB expression in MPTP-lesioned mice treated with L-DOPA (25 mg/kg). These results suggest that GP-EX improves impairment of spatial memory by modulating the expression of NMDA receptors and the activity of ERK1/2 and CREB.
In addition, MPTP-lesioned mice exhibit anxiety disorders. 19 It has been reported that GP-EX (50 mg/kg) ameliorated the anxiolytic effects in a chronic stress-induced mouse model 23 and MPTP-lesioned mouse model of PD. 19 Chronic stress may injure the dorsal striatum, with effects on hippocampus-dependent memory functions in mice and humans, 34,35 suggesting that the ameliorating effects of GP-EX on chronic stress-induced anxiety disorders may improve memory deficits. 34,35
It has been reported that L-DOPA has dual effects on neurotoxicity and dopamine biosynthesis in PC12 cells and dopaminergic neuronal cells. L-DOPA at low concentrations (10–20 μM) does not show cytotoxicity in PC12 cells, however, L-DOPA at high concentrations (100–200 μM) causes oxidative-induced cytotoxicity in PC12 cells, which leads to decreases in dopamine biosynthesis. 36 Low doses of L-DOPA (6–15 mg/kg) are also a benefit and increase dopamine levels in the brain, 37,38 whereas long term or high doses of L-DOPA (20–30 mg/kg) may cause oxidative-induced neurotoxicity of dopaminergic neuronal cells in MPTP- or 6-OHDA-lesioned animal model of PD. 19,39 In addition, L-DOPA may improve certain cognitive functions that are associated with the severely depleted dorsal striatum. 7 By contrast, L-DOPA administration enhances high-level cognitive deficits in patients with PD, 8,9 and high levels of L-DOPA (200 mg/kg) do not improve learning and memory dysfunctions in MPTP-lesioned rats. 40 In the present study, MPTP-induced deficits of habit learning memory and spatial memory were improved by the low concentration of L-DOPA (10 mg/kg), but not L-DOPA (25 mg/kg), for 21 days (Figs. 2 and 3). Furthermore, treatment with GP-EX (50 mg/kg) improved habit learning memory and spatial memory in MPTP-lesioned mice treated with L-DOPA (both 10 and 25 mg/kg) (Figs. 2 and 3).
Recently, long-term treatments with low doses of L-DOPA (10 mg/kg) were shown to be toxic in dopaminergic neurons in the 6-OHDA-lesioned rat model of PD through the ERK-c-Jun (Ser 73) system. 41 Repeated treatments with nontoxic L-DOPA (10 mg/kg) induced oxidative-induced cytotoxicity through the Epac-sustained ERK system in both dopaminergic neuronal and PC12 cells. 36,42,43 Furthermore, chronic and high levels of L-DOPA induced complicated severe motor symptoms, including dyskinesia in PD models. 18,44 These results indicate that long-term treatment with L-DOPA can cause neurotoxicity in dopaminergic neuronal cells, although L-DOPA (10 mg/kg) ameliorated the memory deficits in this study. This hypothesis needs to be studied further.
L-DOPA induces the formation of reactive oxygen species in dopamine neurons and PC12 cells. 2,36 L-DOPA increases nitric oxide production in the striatum, which is associated with PD. 45 Daily repeated L-DOPA administration also increases the nitric oxide generation through activation of neural nitric oxide synthase. 46 GP-EX protects against chronic stress by modulating c-Fos expression. 23,47 GP-EX also shows prophylactic effects on 6-OHDA-induced oxidative cell death in the 6-OHDA-lesioned rat model of PD during long-term L-DOPA treatment. 18 The dose of GP-EX treatment (50–400 mg/kg) does not exhibit adverse effects, such as weight loss, diarrhea, vomiting, and death. 47 With respect to these results, a hypothesis that the protective functions of GP-EX on oxidative stress-induced neuronal cell death play a role in the improvement of habit learning memory and spatial memory in MPTP-lesioned mice treated with L-DOPA can be proposed.
In conclusion, our results are summarized in Figure 7. Treatment with GP-EX (50 mg/kg) showed protective effects on deficits of both habit learning memory and spatial memory by activating the dopaminergic neuronal system and modulating the NMDA receptor-ERK-CREB system in the MPTP-lesioned mouse model of PD treated with L-DOPA (10 and 25 mg/kg). GP-EX may serve as an adjuvant phytonutrient for memory deficits in patients with PD receiving long-term L-DOPA treatments.
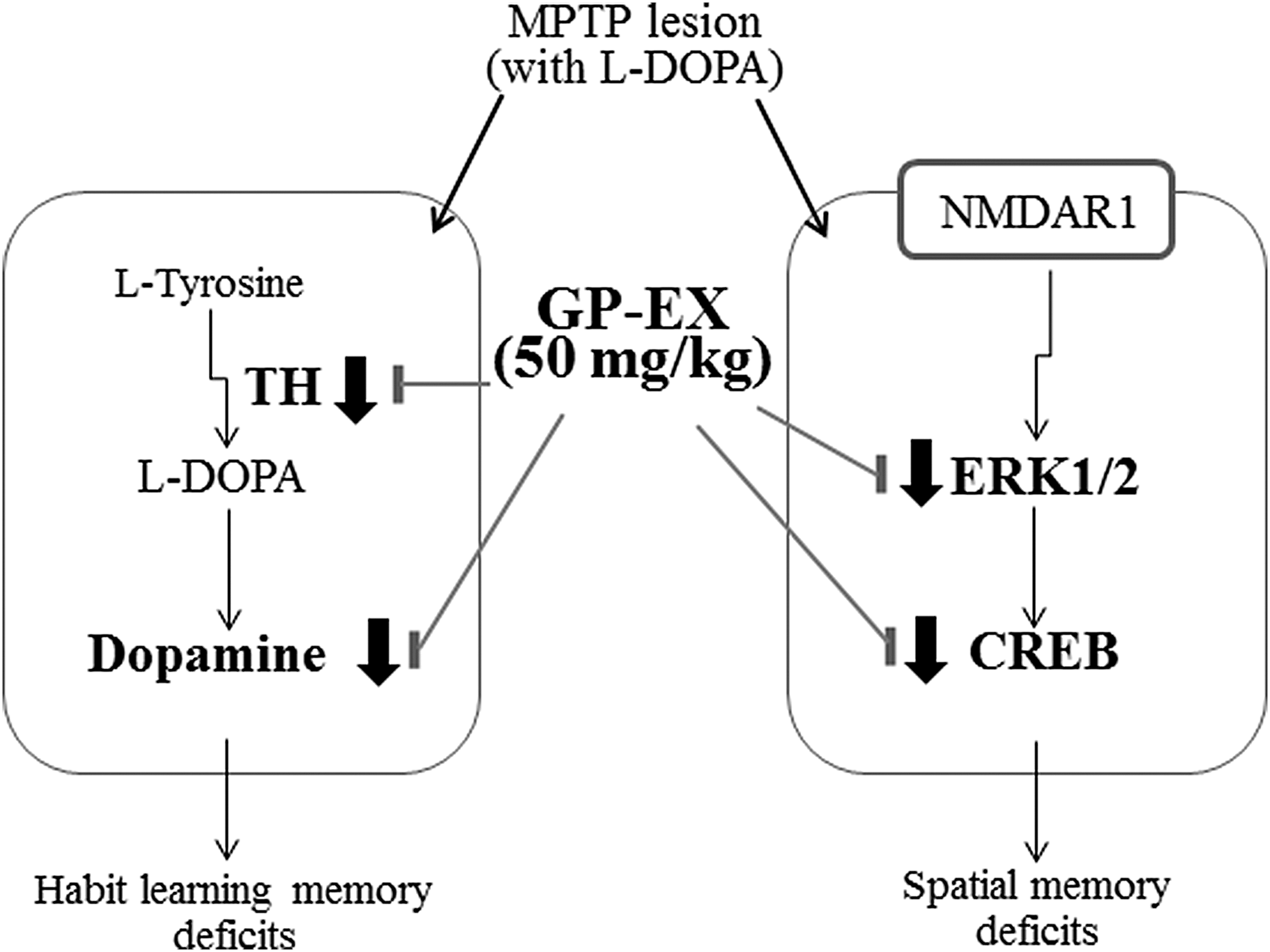
MPTP-lesioned mice showed habit learning memory deficits by reducing TH expression and dopamine levels in the nigrostriatal region and also showed spatial memory deficits by modulating NMDA receptor (NMDAR1)-mediated signaling system in the hippocampal region, which were further aggravated by treatment with L-DOPA (25 mg/kg) (Table 1 and Fig. 1). In this study, treatment with GP-EX (50 mg/kg) showed protective effects on deficits of both habit learning memory and spatial memory by activating the dopaminergic neuronal system and modulating the NMDAR1-ERK-CREB system in MPTP-lesioned mouse model of PD treated with L-DOPA (25 mg/kg). PD, Parkinson's disease.
Footnotes
Acknowledgment
This research was financially supported from the National Research Foundation of Korea (Grant No. 2013R1A1A2058230, 2015–2016), Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.