
Abstract
We investigated the antiangiogenic effects of Lindera obtusiloba Blume (Hwangmaemok, HMM), which is a plant in the Lauraceae family that is commonly used to treat colds and gastritis. Moreover, given that a recent study reported the inhibitory effects of HMM extract on cancer metastasis, we hypothesized that HMM extract might possess and antiangiogenic function. Thus, this study was conducted to investigate the effects of HMM extract on endothelial cell proliferation, migration, and neovascularization in chick chorioallantoic membrane (CAM), and investigated the molecular mechanism of antiangiogenesis using a ProteoChip-based proteomics technology. To examine the effects of HMM extracts on endothelial cell proliferation and migration, we conducted basic fibroblast growth factor (bFGF)-induced human umbilical vein endothelial cell (HUVEC) proliferation and migration. To assess the molecular mechanism of the antiangiogenic effects of HMM extract, a ProteoChip-based forwarded phase antibody array was employed to identify the differential expression of cell cycle proteins in HMM-treated HUVECs. HMM extract inhibited bFGF-induced HUVEC proliferation and migration in a dose-dependent manner and CAM angiogenesis. The ProteoChip-based antibody microarray data showed upregulation of Nibrin/NBS1 and downregulation of Plk-1 and Cyclin E, which are involved in cell division and controlling the cell cycle in bFGF-induced HUVECs. These data suggest that HMM may be a potent antitumor medicinal herb. The present study demonstrates that the antiangiogenic effect of HMM may be due to suppression of endothelial cell proliferation and migration. Taken together, these results emphasize the potential to use HMM extract as a potent angiogenesis inhibitor to treat cancer.
Introduction
A
Lindera obtusiloba Blume (Hwangmaemok, HMM) is a common plant of South Korea and China that has long been used to treat stomach ache and fever. Recently, the antioxidant and antimicrobial activities of HMM extract have been investigated to determine whether it has the potential for use in inhibition of the inflammatory response. 13 Actifolin, pluviatilol, 5,6-dihydromatairesinol, and (+)-syringaresinol in the extract of HMM have been shown to have antiallergic, antifibrotic, apoptotic, and anti-inflammatory effects. 14 Moreover, HMM extract was recently found to inhibit the cancer metastasis process with Matrix metalloproteinases. 15
In this work, we focused on investigating the antiangiogenic mechanism of HMM through differential expression profiles of cell signaling proteins in endothelial cells using antibody microarray technology.
Materials and Methods
Plant material
The L. obtusiloba Blume was purchased from Daejeon University Korean Medicine Hospital. It was identified and authenticated by Prof. M.R. Jin. Voucher specimens (No. LO-2014-03) have been deposited at the Institute of Traditional Medicine and Bioscience in Daejeon University.
Preparation of HMM extract
HMM (100 g) was boiled at 100°C in 1.0 L of distilled water for 3 h. And the water extract was then concentrated into 15 g (Yield: 15%) at 60°C for 30 min using a Thin Film Evaporator. The extract was lyophilized and then stored at −20°C until use. The HMM was dissolved in DMSO.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were purchased from InnoPharmaScreen, Inc. (Asan, Korea). Cells were cultured in a complete M199 medium (Welgene, Daegu, Korea). HUVECs (at passages 3–5 using) were cultured in a humidified atmosphere of 5% CO2 at 37°C.
In vitro HUVEC proliferation assay
HUVEC proliferation was evaluated by BrdU assay according to the manufacturer's instructions (Roche, Germany). Briefly, HUVECs (2 × 103 cells/well) were added to 96-well tissue culture plates coated with gelatin and cultured overnight. The cells were then treated with HMM extract or Gly-Arg-Gly-Asp-Ser-Pro (GRGDSP) (Peptron, Inc., Korea) in the presence or absence of bFGF and incubated for 72 h. All data were measured in triplicate.
Wound-healing migration assay
A wound area was generated with a sterilized micropipette tip in confluent cultures of HUVECs on six-well culture dish. Cellular debris was washed with phosphate-buffered saline (PBS). And HMM extract was then treated in the cells. The width of wound area was captured and photographed using an inverted microscope (Olympus, Japan). 16
Ex vivo bFGF-induced chorioallantoic membrane angiogenesis
A chorioallantoic membrane (CAM) assay in vivo was performed as previously described. Fertilized eggs were incubated at 37°C in humidified air. Ovalbumin (4–5 mL) was extracted from the bottom of the eggs using a syringe on day 3, and windows were made within the tops of the eggs. The windows were sealed with transparent tape, and the eggs were further incubated until day 9. Thermonox (Nunc, Naperville, IL) containing bFGF (200 ng) or bFGF plus HMM extract was implanted onto the CAM of an individual embryo. After 72 h incubation, intralipose (a white fat emulsion solution) was injected into the CAM for clear visualization of blood vessel formation, and each CAM was observed under a microscope and photographed. 17
Protein extraction and labeling
HUVEC lysates were extracted with Lysis-M™ (Roche) mammalian cell extraction buffer. Each protein was labeled with Cy3 or Cy5 (GE healthcare, United Kingdom) as described in the manufacturer's manual and free dyes were removed by Spin columns (Sigma, USA). Samples were stored at 4°C. 18
Expression profile analysis of cell cycle proteins using IPS forward phase protein chip-based antibody array
We conducted an antibody array using protein A (200 μg/mL)-coated ProteoChip. Specific antibodies were selected against cell cycle-related proteins in HUVECs. Each antibody was dispensed on the chip by 10-fold dilution in 30% glycerol and allowed to stand overnight at 4°C. The chip was blocked with 3% BSA in PBS for 1 h at room temperature. After the chip was rinsed with PBS with Tween 20 (PBST) (0.05% tween-20 in PBS) and dried, it was stored at 4°C until use. Applying the labeled cell lysates diluted in the reaction buffer solution to the antibody array was carried out and incubated for 1 h at 37°C in the dark room. After the reaction, it was rinsed with PBST and dried. The antibody array chips were scanned using a Gene-Pix 4100A microarray Scanner (Axon Instruments, Union City, CA) with 532 and 635 nm lasers. Image analysis was performed with GenePix Pro (version 6.0) software package (Axon instruments). INR (internally normalized ratio) of all spots were calculated as previously described. 18
Western blot analysis
HUVECs were cultured in M199 until 70% confluency and the medium was replaced with fresh low serum medium with or without HMM. The cells were washed with PBS (Welgene, Daejeon, Korea) and cell lysates were prepared in Lysis-M buffer (Roche) containing protease and phosphatase inhibitor cocktails (both from Roche), then clarified by centrifugation. Lysates containing 50 μg of protein were loaded into each well and separated through sodium dodecyl sulfate (SDS)-gel electrophoresis using 4–20% SDS gel grade (Biorad, Hercules, CA). Gels were soaked in transfer buffer (16 mM Tris-HCl, 30 mM glycine, and 20% methanol), after which proteins were transferred to polyvinylidene fluoride (PVDF) membranes (Merck Millipore, Darmstadt, Germany). Nonspecific binding sites were blocked by incubation with 5% nonfat dry milk in PBST (137 mM NaCl, 27 mM KCl, 100 mM Na2HPO4, 20 mM KH2PO4, and 0.05% Tween 20, pH7.4). The PVDF membranes were then incubated with primary antibodies against cyclin E (1: 500) and β-actin (1:10,000) in PBST containing 5% nonfat dry milk at 4°C overnight. Next, membranes were washed with PBST and incubated with secondary antibodies (anti-mouse at 1:10,000, anti-rabbit at 1:16,000). Finally, signals were developed using an enhanced chemiluminescene western blotting detection kit and exposed to X-ray films.
Statistics
All data are representative of three independent experiments with duplicates and expressed as the mean ± standard error of the mean. The values were compared with the control using analysis of variance followed by unpaired Student's t-test. P values of *P < .05, **P < .01, and ***P < .001 were regarded as statistically significant.
Results and Discussion
Inhibitory effects of HMM extract on bFGF-induced HUVECs
To investigate the antiangiogenic function of HMM, we conducted experiments on HUVEC proliferation and migration assay. HMM extract inhibited bFGF-stimulated HUVEC growth in a dose-dependent fashion compared with the stimulated HUVEC control group (Fig. 1). A half-maximal inhibitory concentration (IC50) of HMM was 0.35 ± 0.04 mg/mL dissolved in DMSO (Fig. 1A). We observed that there was no effect on cell proliferation at the concentrations of DMSO solvent (Fig. 1B). To further examine another antiangiogenesis function of HMM, in vitro migration assay was employed. Dose-dependent suppression of bFGF-induced HUVEC migration was observed in response to treatment with HMM. The IC50 value of the migration was determined to occur at 0.243 mg/mL, while complete inhibition occurred at 1.0 mg/mL after 10 h of incubation (Fig. 2). Moreover, HMM extract appeared to be more effective at inhibiting cell migration than the positive control group treated with GRGDSP, an antagonistic peptide against integrin α v β 3. The GRGDSP sequence (Gly-Arg-Gly-Asp-Ser-Pro) is well known as a common binding motif that is recognized by a series of integrin family proteins including integrin α v β 3. 19
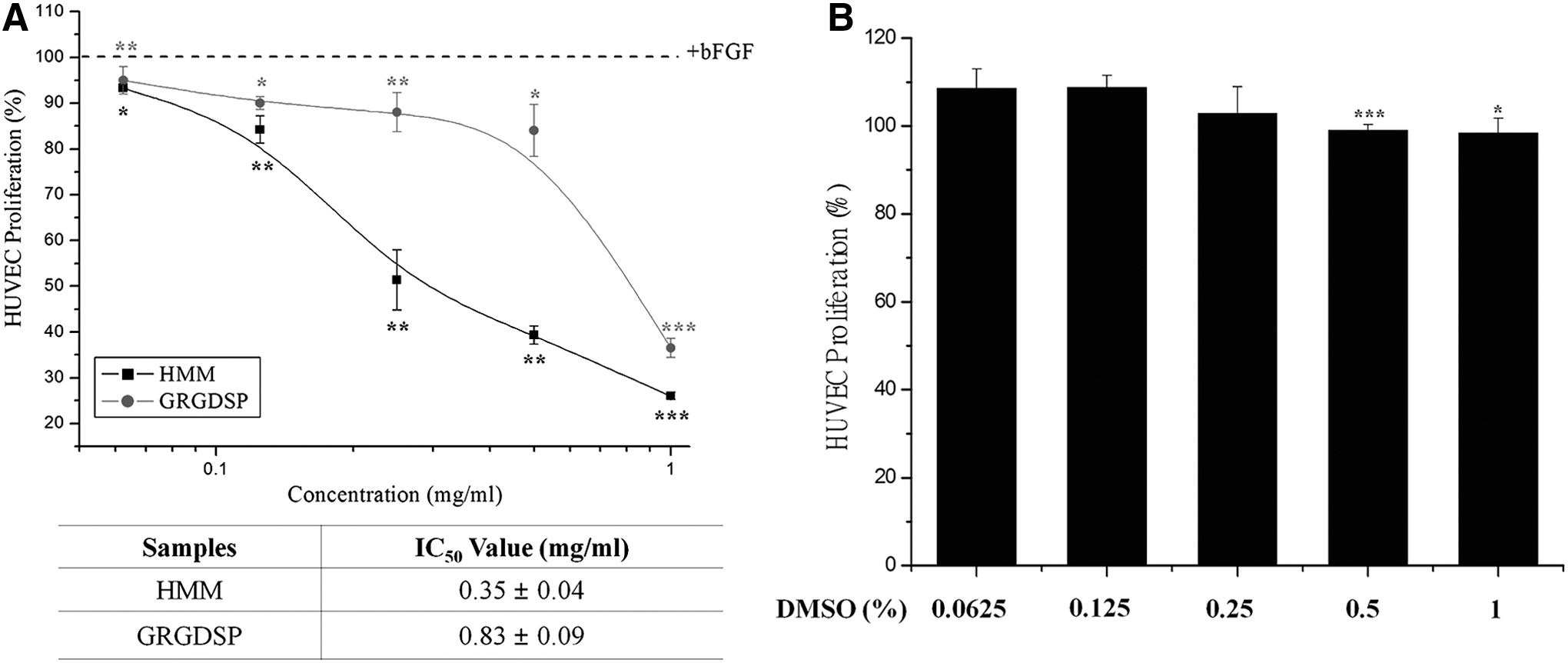
Inhibitory effect of HMM extract on HUVEC proliferation
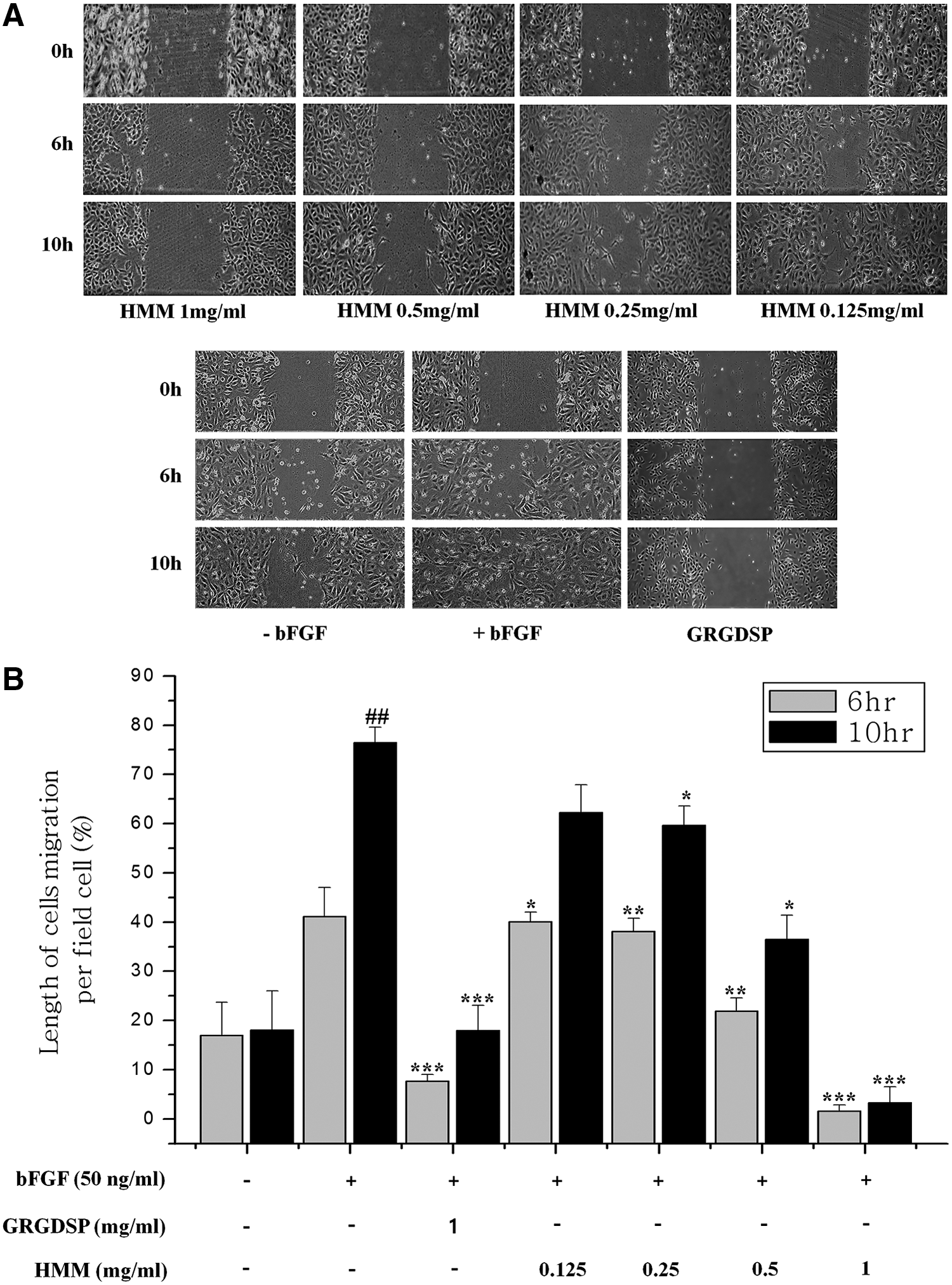
Inhibitory effect of HMM extract on HUVEC migration
Inhibitory effect of HMM extract on CAM angiogenesis ex vivo
To further confirm the antiangiogenic effect of HMM, we employed the CAM angiogenesis assay ex vivo. The area of CAM below the disks without treatment showed a normal density and branching pattern of the blood vessels, indicating that disk weight did not affect their growth. In contrast, the neovascularization of CAM treated with HMM (10 μg/egg) for 72 h was inhibited (Fig. 3). This finding was consistent with previous data obtained from HUVEC proliferation and migration assay. Overall, these results imply that the suppression of HUVEC proliferation and migration by HMM can be an action mechanism of antiangiogenesis of HMM.
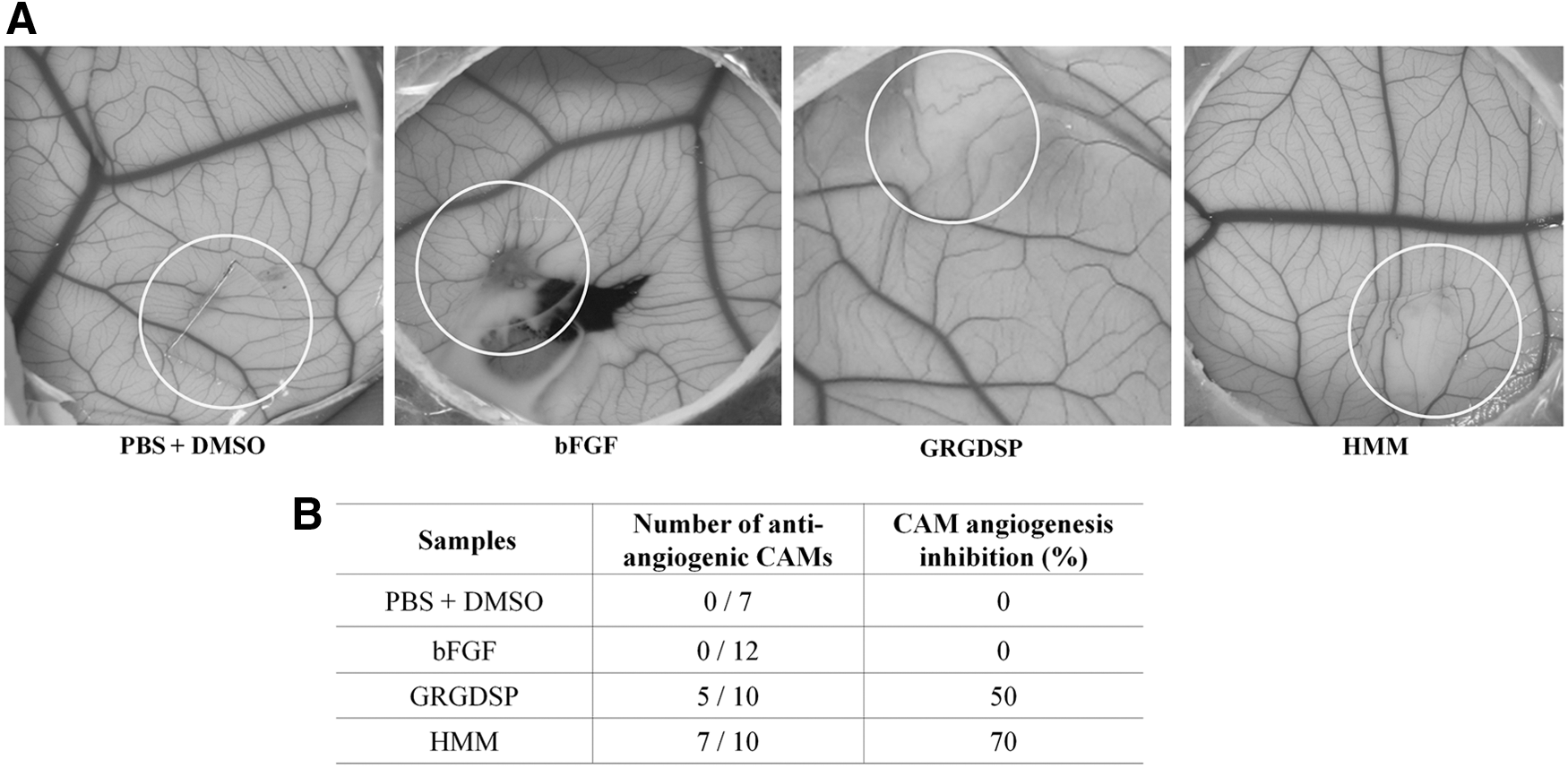
Inhibitory effects of HMM extract on bFGF-induced CAM angiogenesis.
Proteomic analysis of cell signaling protein in HMM-treated HUVECs
To investigate the molecular mechanism of the antiangiogenic function of HMM, we conducted differential expression profiling of cell signaling proteins in HMM-treated HUVECs using an IPS-Forwarded Phase Antibody Array (Fig. 4). HUVEC lysates treated with bFGF alone (Control group) or HMM in the presence of bFGF were labeled with both Cy3 and Cy5. The labeled lysates were then mixed and spotted on antibody microarray. After the reaction, the fluorescence intensity of each spot was measured to determine differential protein expression pattern in the lysates. We identified proteins that indicate changes in protein expression in response to HMM. The distribution of fluorescence intensities for all spots was normalized based on the mean values of the Cy5:Cy3 ratios (Fig. 4A). The results revealed that Plk-1 and Cyclin E were downregulated in HMM-treated HUVECs, while Nibrin/NBS1 was upregulated (Fig. 4B, C). The levels of all other proteins were unchanged based on the antibody microarray analysis. Cyclin B1 and E have been reported to be involved in cell division signaling, and cyclin family members and polo-like kinase (Plk) are important regulators of cell cycle progression through the S phase and mitosis. 17,19,20 The presence of bFGF markedly induced cyclin E-CDK2 activity and S phase entrance. Blockade of the FGF action profoundly inhibited cyclin E expression and cyclin-dependent kinase-2 (CDK2) activity, but not D-type cyclin expression and CDK4 activity. 21 These observations suggest that downregulation of Plk-1 and Cyclin E can suppress cell growth. 22 Nibrin is a DNA double-strand break repair protein that is known to be a marker of various tumors. 23 –26 Our result showed that upregulation of Nibrin/NBS1 is a phenotype of a cellular defense mechanism in cells. Overall, our antibody array data indicate that the inhibitory effects of HMM on the proliferation of HUVECs may be due to the decreased expression of Plk-1 and Cyclin E, which are involved in the induction of cell division.
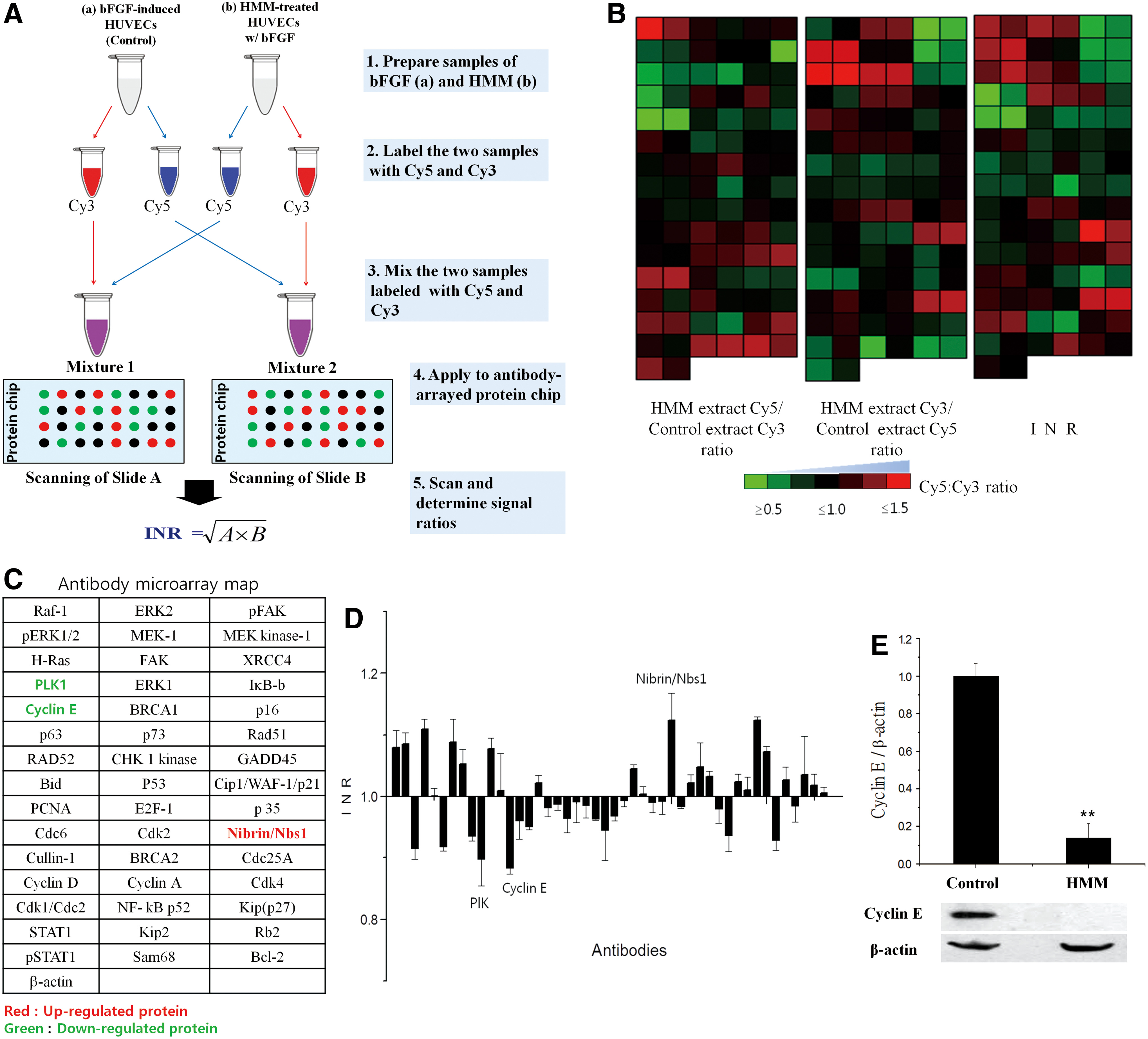
Analysis of differential expression of cell cycle proteins in HUVECs treated with HMM extract using IPS FPPC-based antibody microarray.
Collectively, our study demonstrated that antiangiogenic activity of HMM was due to inhibition of HUVECs proliferation and migration via upregulation of Nibrin/NBS1 and downregulation Plk-1 and Cyclin E in the cells. Further study is needed to further elucidate the antiangiogenic of HMM using animal models.
Footnotes
Acknowledgments
This research was supported in part by grants of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare (grant number: HI14C0849).
AUTHORS' CONTRIBUTIONS
M.K. and Y.M. carried out experiments. I.K. and H.Y. designed the works, drafted, and finalized the article.
Author Disclosure Statement
No competing financial interests exist.