
Abstract
The aim of our experiment was to evaluate the anticancer effect of bamboo salt (BS) on C57BL/6 mice in an azoxymethane (AOM)/dextran sodium sulfate (DSS)-induced colon cancer model. BS, solar salt, and purified salt were evaluated for their protective effects during AOM/DSS-induced colon carcinogenesis in C57BL/6 mice. BS, especially after baking for nine separate intervals (BS9x), suppressed colon carcinogenesis in the mice. BS9x decreased colon length shortening, weight-to-length ratios, and tumor counts. Pathological evidence from histological evaluation by hematoxylin and eosin staining also revealed suppression of tumorigenesis. BS9x lowered serum levels of proinflammatory cytokines (TNF-α, IL-6, and IL-1β) to close to those of the Normal group. Additionally, BS9x suppressed colon mRNA expression of proinflammatory factors and significantly regulated mRNA levels of the apoptosis-related factors, Bax and Bcl-2, and the cell cycle-related genes, p21 and p53. Additionally, immunohistochemistry showed that BS promoted p21 expression in the colon. Taken together, the results indicate that BS exhibited anticancer efficacy by modulating apoptosis- and inflammation-related gene expression during colon carcinogenesis in mice, and repetition in baking cycles of BS enhanced its anticancer functionality.
Introduction
B
Colorectal cancer is known to result from environmental factors, including diet-derived carcinogens, chronic inflammation and pathogens, and genetic components. 11 Additionally, a significant connection between table salt and colorectal cancer has been reported by Shikata et al., 12 and consuming salted shellfish or meat can increase the incidence of colorectal cancer. 13
Even though many functions of BS were unveiled through the studies referenced above, the preventive functions of BS against colorectal cancer were evaluated using only HT-29 human colon cancer cells. 6 Thus, we used the azoxymethane (AOM)/dextran sodium sulfate (DSS)-based mouse colorectal cancer model to evaluate the preventive effect of BS and reinforce the previous results in vitro by direct observations of changes in colon conditions (colon length, weight, and tumor counts) and alterations in serum cytokine profiles.
Materials and Methods
BS baked three times (BS-3x) and nine times (BS-9x) was purchased from Insanga Co. (Hamyang, Gyeongsangnam-do, Korea), and the solar salt was provided by the Yeongbaek salt field (Shin-An, Jeollanam-do, Korea; SS-Yb; this solar salt was used as a raw material to manufacture BS). Purified salt (PS) was purchased from Hanju Co. (Ulsan, Korea).
AOM was purchased from Sigma Co. (St. Louis, MO, USA), and dextran sulfate sodium (DSS; molecular weight = 36,000–50,000) was obtained from the MP Biomedical Co. (Solon, OH, USA). ELISA kits for analysis of tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), and interleukin 1-beta (IL-1β) were purchased from Biolegend Co. (San Diego, CA, USA). TRIzol, reverse transcriptase reagents, and agarose powder were purchased from Invitrogen Co. (Carlsbad, CA, USA). A PCR thermal cycler used for this experiment was manufactured by Invitrogen (Singapore, Singapore). All primers, TAE buffer, and ethidium bromide were purchased from the Bioneer Co. (Daejeon, Korea).
Animal studies
Six-week-old C57BL/6J male mice, purchased from Samtach Co. (Osan, Gyeonggi-do, Korea), were acclimatized for 1 week before the main experiment. After acclimatization, the mice were divided into the following experimental groups: Normal, Control (AOM/DSS treatment only), purified salt-fed (PS), solar salt-fed (SS-Yb), three-time baked bamboo salt-fed (BS-3x), and nine-time baked bamboo salt-fed (BS-9x) groups. Mice from all of the experimental groups, except the Normal group, were administered AOM by intraperitoneal injection at a dose of 10 mg/kg on the first day of week 1. One week later, the mice were given 0.02 g/mL DSS dissolved in drinking water for an entire week, on week 3, and week 6. Beginning at the time of the AOM injection, a salt-supplemented diet with 0.47% salt (provided by Dooyeol Biotech, Seoul, Korea), which is equivalent to a human salt intake of 5 g/day, was provided to animals as prescribed for each experimental group. At the end of the experiment (week 8), the mice were put on a 12-h fasting regimen and sacrificed. Blood was collected from the abdominal aorta and stored overnight at 4°C, followed by centrifugation at 3,000 g for 15 min. Serum was collected from these samples and stored at −70°C until further analysis. Colons were surgically removed and assessed for length, weight, and tumor numbers, 14 followed by storage at −70°C for molecular analysis and a portion stored in 10% formalin for further histological studies. The protocol used for this experiment was approved by the Institutional Animal Care and Use Committee (PNU-IACUC; approval number PNU-2013-0450) of Pusan National University (Busan, South Korea).
Histological evaluation by hematoxylin and eosin staining
The colon tissues were fixed in 10% (v/v) neutral buffered formalin for 24 h. The formalin-fixed colon sample was embedded into a paraffin block and sliced into multiple sections at a thickness of 5 μm. Sections were then stained with hematoxylin and eosin (H&E) reagent, and the images were acquired using an Olympus BX 51 microscope equipped with a digital camera system of cell sense entry (Olympus, Japan).
Analysis of serum proinflammatory cytokines
Cytokines, including TNF-α, IL-6, and IL-1β, were analyzed using enzyme-linked immunosorbent assay (ELISA) kits according to the appropriate manufacturer protocol. The optical density of each well that had undergone an antigen–antibody reaction was measured by an ELISA reader at a wavelength of 450 nm.
Analyses of colon mRNA expression of cancer-related genes
mRNA expression levels of proinflammatory cytokines (TNF-α, IL-6, and IL-1β), apoptosis-related factors (Bcl-2 and Bax), and cell cycle-related factors (p53 and p21) in the colon mucosa of mice were measured using a reverse transcription (RT)-PCR assay. 10 Total RNA was isolated from the colon tissue (100 mg) using TRIzol reagent according to the manufacturer's recommendations and centrifuged at 12,000 g for 15 min at 25°C before chloroform (0.2 mL) was added. Isopropanol was then added to the supernatant at a 1:1 ratio, and the RNA was pelleted by centrifugation (12,000 g for 15 min). After washing the pellet with ethanol, the RNA was solubilized in diethylpyrocarbonate (DEPC)-treated RNase-free water and quantified by measuring the absorbance at 260 nm using a UV-2401PC spectrophotometer (Shimadzu, Kyoto, Japan). Equal amounts of RNA (1 μg) were reverse transcribed in a master mix, containing reverse transcriptase buffer, 1 mM dNTPs, 500 ng of each oligo dT18 primer, 140 U MMLV reverse transcriptase, and 40 U RNase inhibitor for 45 min at 42°C. PCR was then carried out in an automatic thermocycler for 25 cycles (94°C for 30 sec, 55°C for 30 sec, and 72°C for 40 sec), followed by an 8-min final incubation at 72°C. The PCR products were separated using 2% agarose gels and visualized by ethidium bromide staining. GAPDH expression was used for normalization, and gene expression was quantified using ImageJ software.
Immunohistostaining of p21 antibody in colon tissues
Immediately after fixation of colon tissues with 10% (v/v) neutral buffered formalin for 24 h, 4-μm-thick sections were cut consecutively from the blocks, followed by deparaffinization in xylene and rehydration in a graded alcohol series. The tissue samples were analyzed with a BOND-MAX autoimmunostaining system (Leica Microsystems, Wetzlar, Germany). Antigen retrieval was performed in pH 6.0 citrate phosphate buffer using a microwave. The primary antibody for this study was a polyclonal antibody against p21(1:100; Dako, Denmark). The slides were counterstained with Meyer's hematoxylin. Negative controls were treated similarly without primary antibodies. Images were acquired using a Zeiss Axioskop 2 Plus microscope (Carl Zeiss MicroImaging, Thornwood, NY, USA) equipped with an AxioCamMRc5 CCD camera (Carl Zeiss MicroImaging). The images obtained from those processed were analyzed using I-solution (Martin Microscope Company, Easley, SC, USA).
Statistical analyses
All data are presented as the mean ± SD, and differences between the mean values for individual groups were assessed using a one-way ANOVA, followed by Duncan's multiple range tests. P-values <.05 determined using a software package (SAS Institute, Inc., Cary, NC, USA) were considered statistically significant.
Results
Changes in colon length, weight-to-length ratio, and tumor counts
The colon length of each experimental group is shown in Figure 1a. The average colon length of the Control group was 5.6 ± 0.6 cm, which was markedly shorter than that in the Normal group (6.2 ± 0.5 cm), due to AOM/DSS-driven ulcerative colon cancer. The average colon length of the BS9x-treated mice was 6.8 ± 0.0 cm, which was close to that of the Normal group, followed by BS3x (5.8 ± 0.2 cm), SS-Yb (5.5 ± 0.3 cm), and PS (5.0 ± 1.4 cm; P < .05).
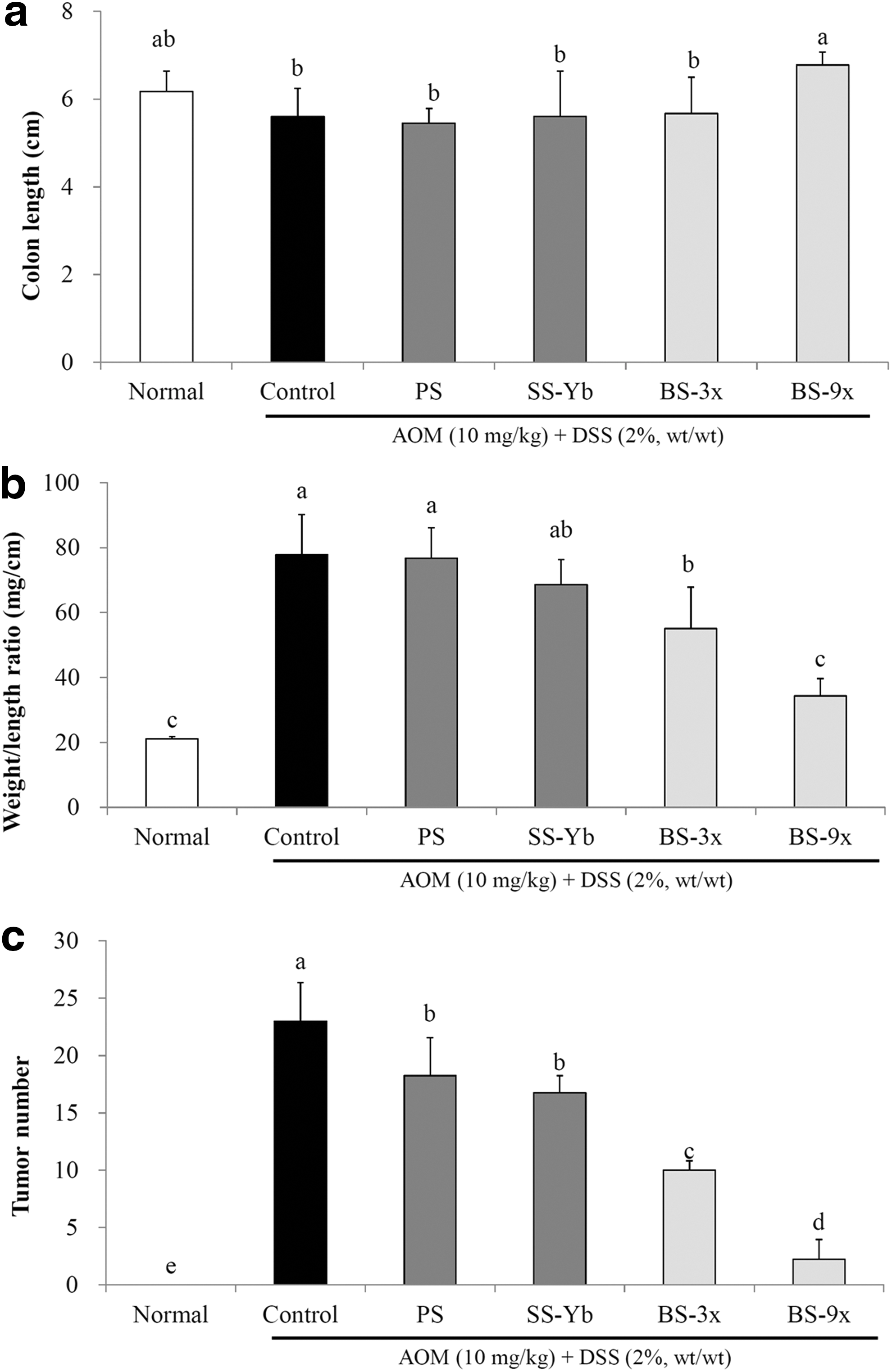
Effects of salt samples on
The colon weight-to-length ratio was observed by dividing colon weight by length as previously described. The average colon weight/length ratio of the Control mice was 77.8 ± 12.3 mg/cm, which was significantly higher than the 21.0 ± 0.7 mg/cm colon length in the Normal group (P < .05). BS9x resulted in an average colon weight/length ratio of 34.3 ± 5.4 mg/cm, which was the lowest among the AOM/DSS-treated groups and closest to that of the Normal group, followed by BS3x (55.0 ± 12.7 mg/cm), SS-Yb (68.5 ± 7.8 mg/cm), and PS (76.7 ± 9.3 mg/cm. Fig. 1b).
The average colon tumor count of the Control mice was 23.0 ± 3.3, whereas no tumors were found in the Normal group. The average colon tumor count from the BS9x-treated mice was 2.2 ± 1.7, which was significantly lower than that of the AOM/DSS-treated groups and close to that of the Normal group (P < .05). In addition, the average colon tumor counts of the other groups were as follows: BS3x (10.0 ± 0.8), SS-Yb (16.7 ± 1.5), and PS (18.2 ± 3.3; Fig. 1c).
Colon histopathology
Pathological changes following salt administration are shown in Figure 2. In comparison with the Normal group, colon tissues that underwent colon cancer induction following the AOM/DSS protocol exhibited loss of intestinal gland cells, tumors on the mucosa, and even invasion of cancer cells into tissues. Such pathogenesis was found to be severe in the Control, PS, and SS-Yb mice. The BS3x-treated mice showed precancerous lesions of adenocarcinoma when compared with those of the Control group. In the picture of BS9x, normal tissue was observed in the right side of colon tissue with goblet cells, whereas cancerous tissue starting to heal, determined by breakage of nucleus, was observed in the left side of colon tissue (identified by the green arrow).
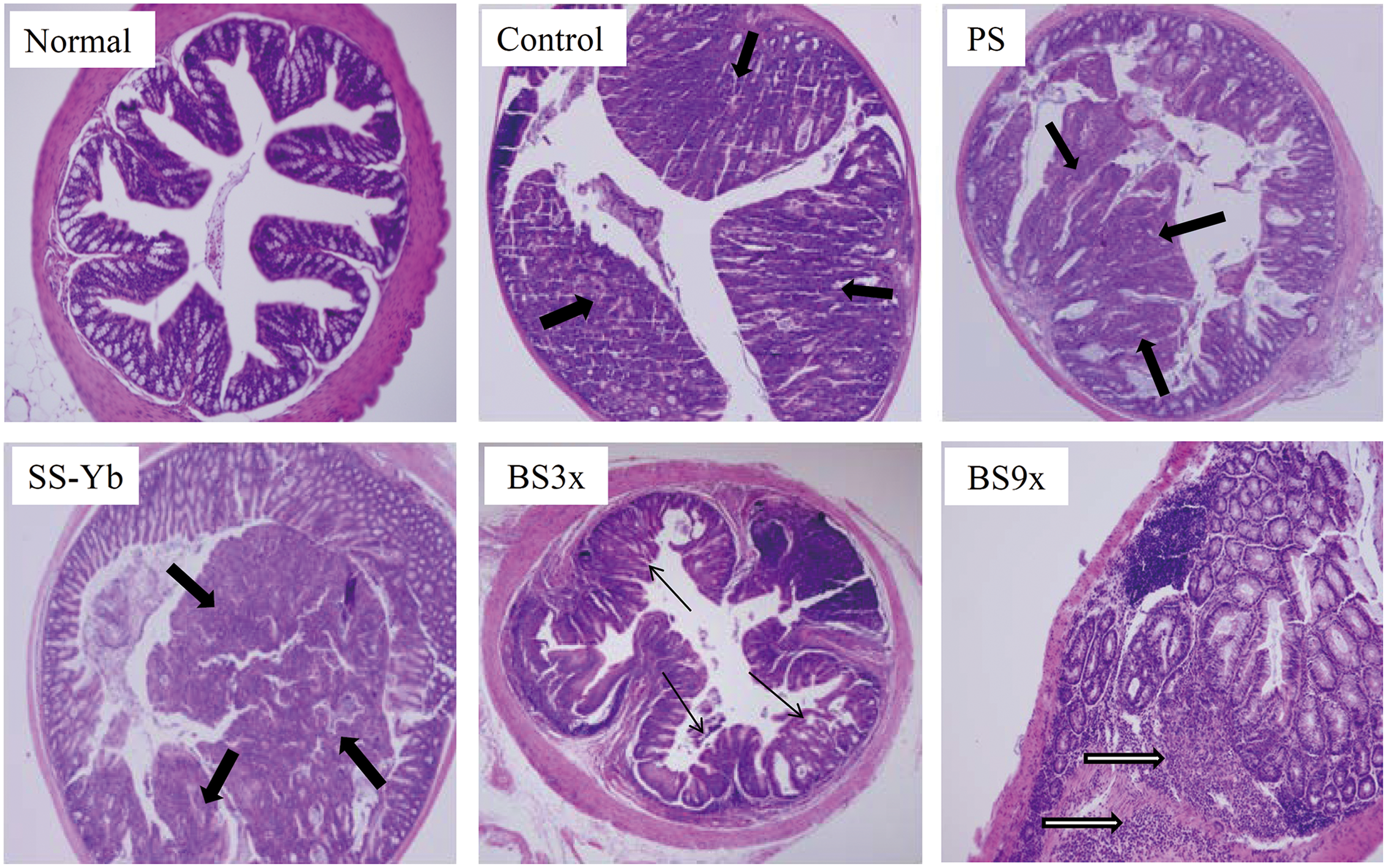
Histological analysis of the colonic mucosae of mice with AOM/ DSS-induced colon cancer using hematoxylin and eosin staining. The thick black arrows in the figure indicate areas of carcinoma, and the thin black arrows in the figure indicate adenoma. In addition, the bordered black arrows indicate the cancerous tissues on the way to healing. Group abbreviations are as described in the legend of Figure 1. Color images available online at
Serum cytokine concentrations
The serum concentrations of TNF-α, IL-6, and IL-1β in the salt-supplemented mice were found to be lower than those of the Control group, as shown in Figure 3.
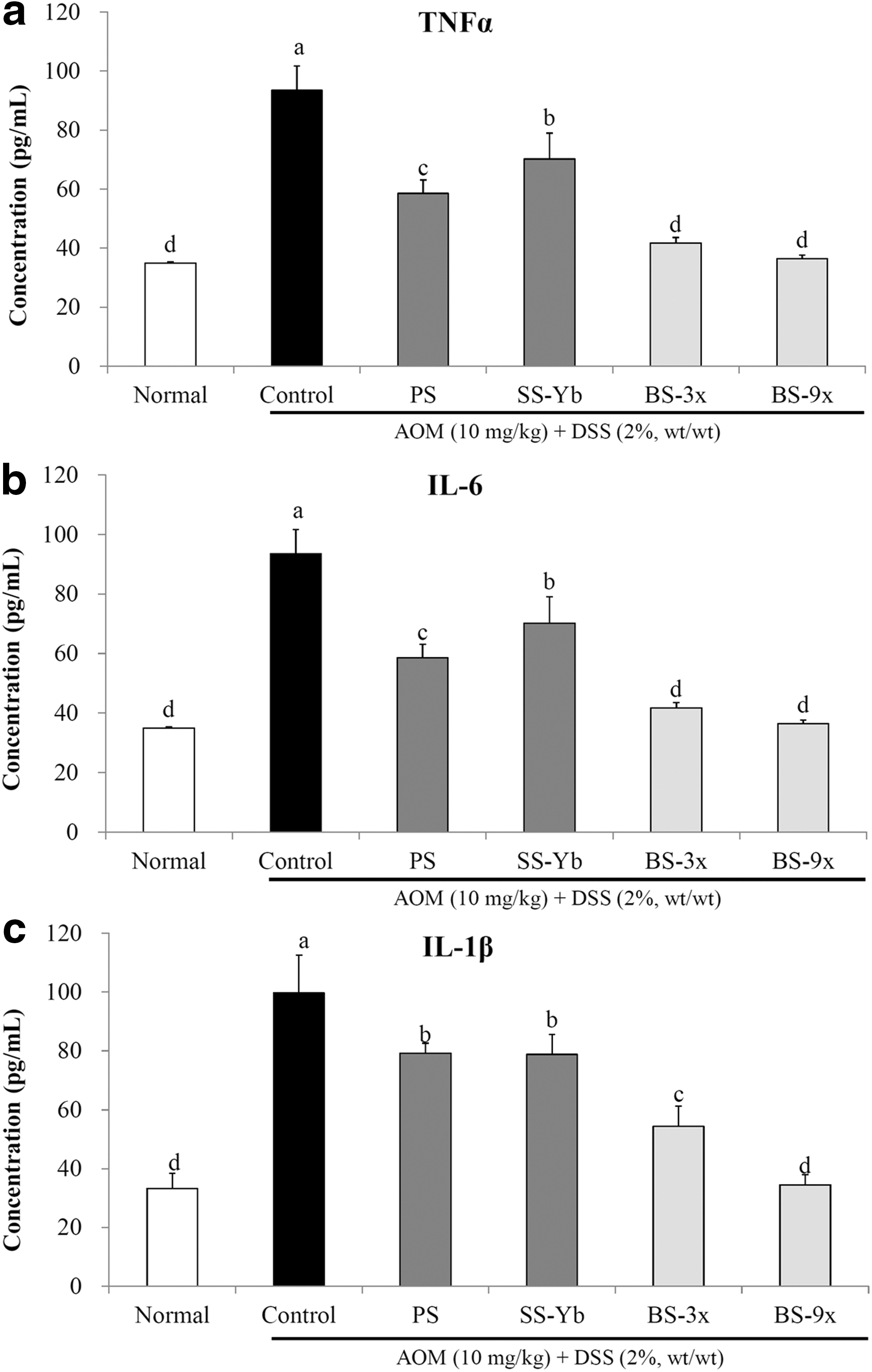
Serum cytokine levels of
The serum TNF-α concentration in the Control group was higher at 97.0 ± 4.1 pg/mL than that of the Normal mice (27.7 ± 4.4 pg/mL) due to the administration of AOM/DSS. Conversely, the BS3x- and BS9x-treated mice were found to have significantly lower concentrations at 50.5 ± 4.7 and 36.7 ± 3.2 pg/mL, respectively (p < 0.05), followed by PS (53.9 ± 7.6) and SS-Yb (65.9 ± 1.6; Fig. 3a).
The serum IL-6 concentration in the Control group was 93.4 ± 8.1 pg/mL, which was higher than the Normal group (34.9 ± 0.4 pg/mL), due to administration of AOM/DSS. BS9x treatment resulted in the lowest concentration of IL-6 (36.4 ± 1.2 pg/mL), followed by BS3x (41.6 ± 1.9), PS (58.6 ± 4.4), and SS-Yb (70.1 ± 8.8; Fig. 3b).
Additionally, a significantly low IL-1β concentration was achieved by BS9x treatment (34.5 ± 3.4 pg/mL), which was close to that of Normal (33.3 ± 5.0), followed by BS3x (54.3 ± 6.8), SS-Yb (78.8 ± 6.8), and PS (79.1 ± 3.4; P < .05. Fig. 3c).
Changes in mRNA expression in colon tissues
mRNA expression of inflammation-related factors in colon tissues
Colon mRNA expression of proinflammatory cytokines is shown in Figure 4. Generally, mRNA expression levels of TNF-α, IL-6, and IL-1β significantly increased in the Control group compared with those in the Normal group (P < .05). Every salt-administered group was found to have a lower expression level of TNF-α, IL-6, and IL-1β than did the Control group; in particular, the BS9x-treated group was found to have an even lower expression level than did the Normal group.
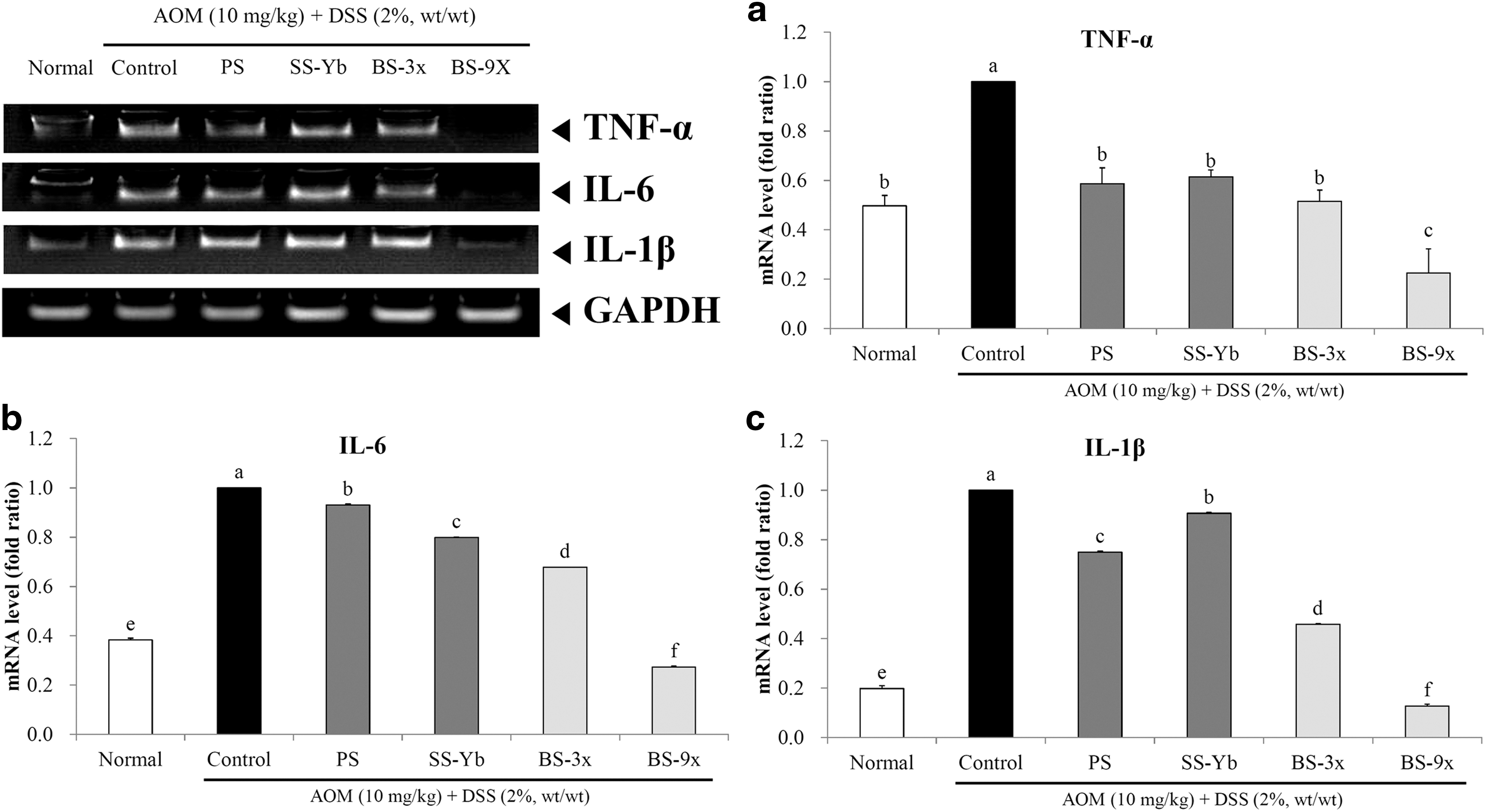
mRNA expression levels of
The mRNA expression of TNF-α, in comparison with Control, was also suppressed in the salt-administered groups. Especially, BS9x achieved the lowest expression level, 0.23 ± 0.10 times that of Control; followed by BS3x (0.52 ± 0.05) and SS-Yb (0.61 ± 0.03; Fig. 4a).
mRNA expression of IL-6 was suppressed in the salt-administered groups when compared with the Control group. In particular, BS9x achieved the lowest expression level (0.27 ± 0.00) compared with that of the Control group, followed by BS3x (0.68 ± 0.00) and SS-Yb (0.80 ± 0.00; Fig. 4b).
The mRNA expression of IL-1β, in comparison with the Control, was also suppressed in the salt-administered groups. Especially, BS9x achieved the lowest expression level, 0.13 ± 0.00 of that of Control; followed by BS3x (0.46 ± 0.00) and SS-Yb (0.91 ± 0.00; Fig. 4c).
mRNA expression of apoptosis-related factors in colon tissues
Colon mRNA expression of apoptosis-related factors (Bax and Bcl-2) was observed and is shown in Figure 5.
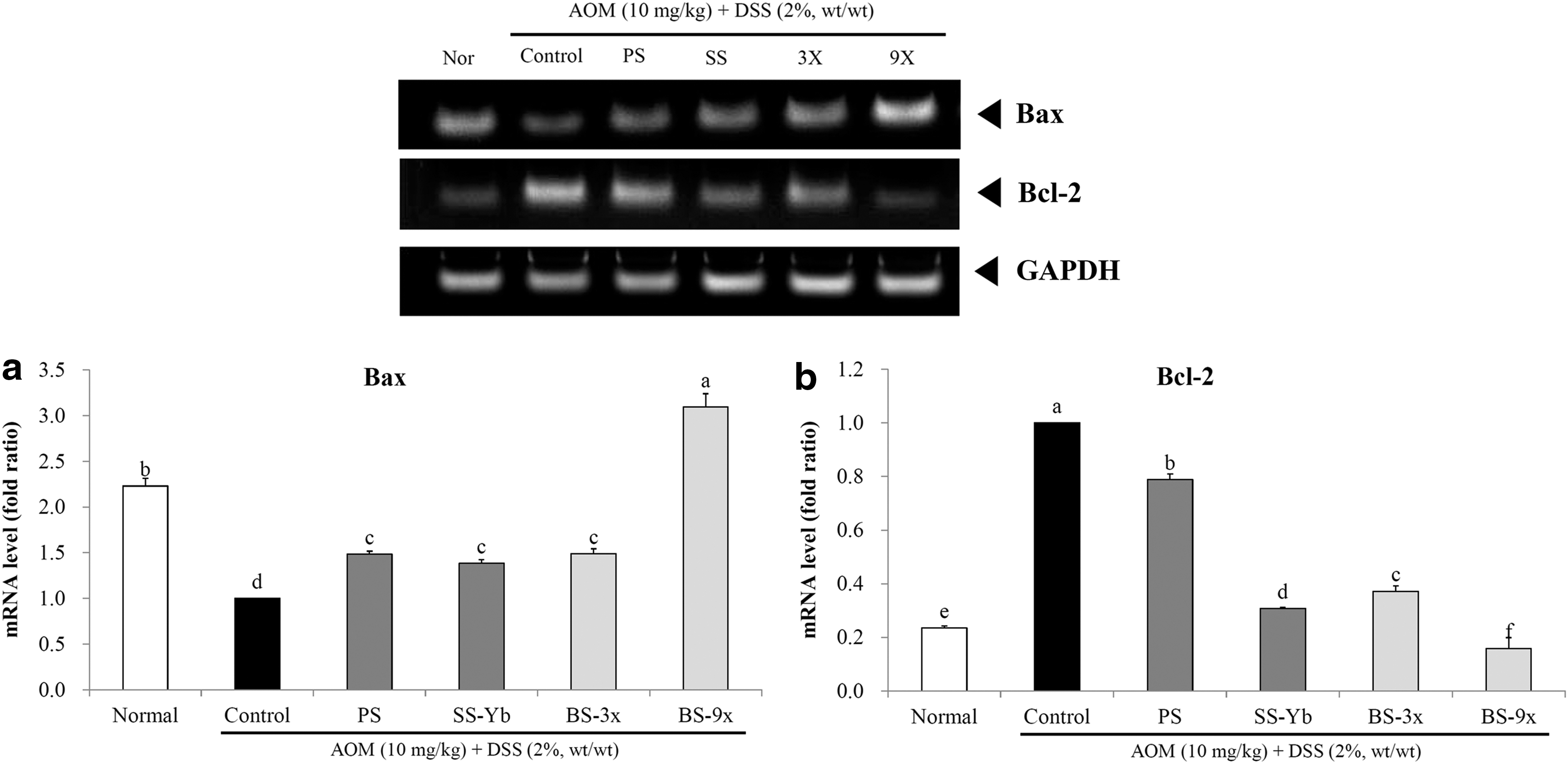
mRNA expression levels of
The mRNA expression of Bax was increased in the salt-treated groups when compared with the Control. Especially, BS9x achieved the highest expression level, 3.09 ± 0.15 times that of the Control, followed by BS3x (1.49 ± 0.05) and SS-Yb (1.38 ± 0.04; Fig. 5a).
The mRNA expression of Bcl-2 was suppressed in the salt-treated groups when compared with that of the Control group. In particular, BS9x achieved the lowest expression level, 0.16 ± 0.01 of that of Control, followed by BS3x (0.37 ± 0.00) and SS-Yb (0.31 ± 0.00; Fig. 5b).
mRNA expression of cell cycle-related factors in colon tissues
The mRNA expression of cell cycle-related factors is shown in Figure 6 for p21 and p53.
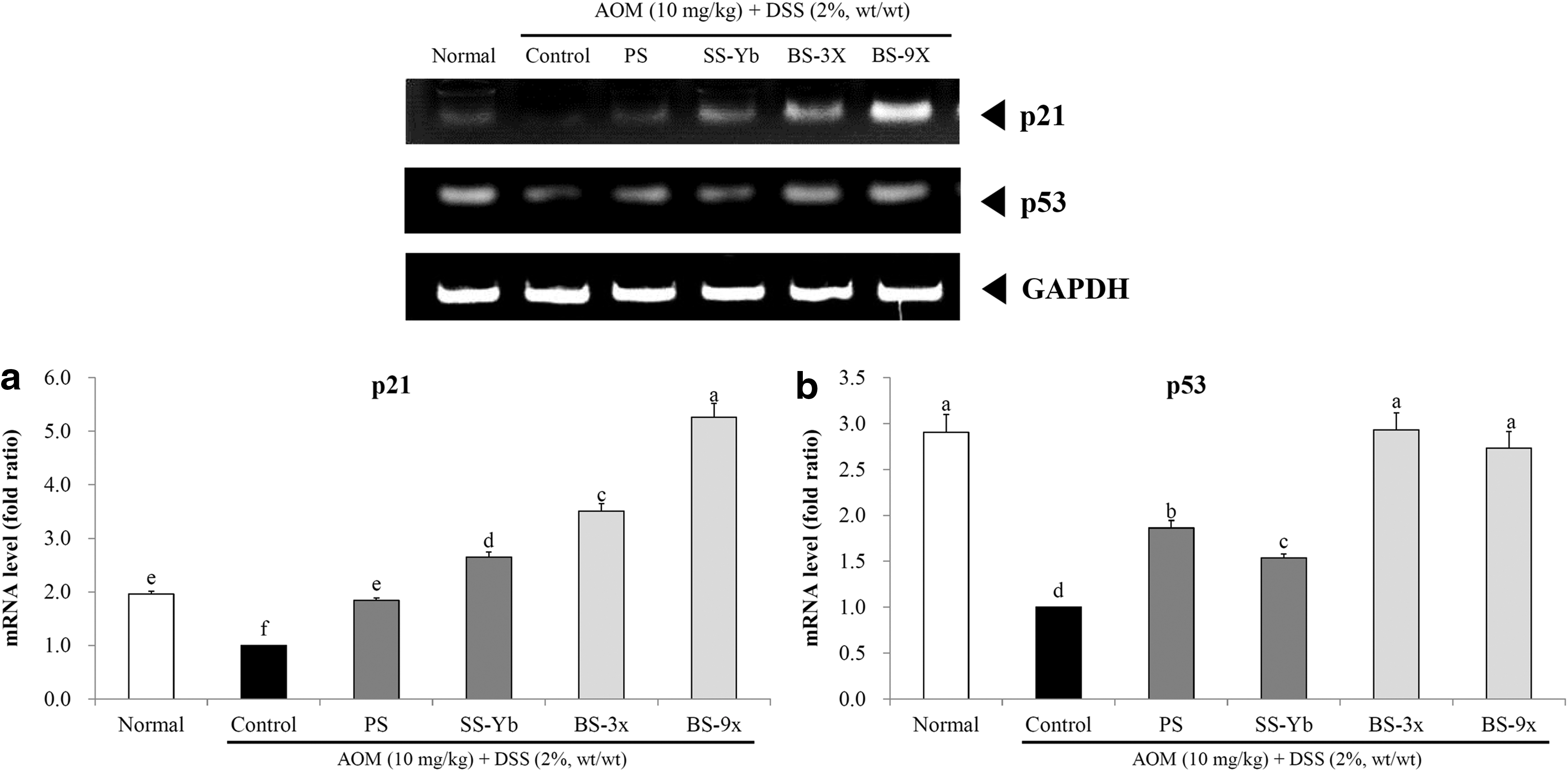
mRNA expression levels of
The mRNA expression of p21, in comparison with the Control, was increased in the salt-treated groups. In particular, BS9x achieved the highest expression level, 5.26 ± 0.26 times that of the Control, followed by BS3x (3.51 ± 0.14) and SS-Yb (2.65 ± 0.09; Fig. 6a).
The mRNA expression of p53 was increased in the salt-treated groups compared with that of the Control. In particular, both BS9x (2.73 ± 0.18 times that of the Control) and BS3x (2.93 ± 0.19 times that of the Control) achieved the highest expression levels, followed by PS (1.86 ± 0.08) and SS-Yb (1.54 ± 0.04; Fig. 6b).
Colon tissue immunohistology
Histopathological changes following salt treatment were confirmed by immunohistochemistry to determine the enteric protein expression of p21, as shown in Figure 7. p21 was found to be partially expressed in the interstitial space of the Normal group, while the colon tissues of the Control group were scarcely stained. PS group showed just a dim staining on the colon tissue, but the stained portions of the interstitial space and the enteric epithelium were increasingly observed in the colon of the SS-Yb group, and its intensity was strengthened in the BS3x-treated mice. The colon tissues of the BS9x-treated mice were found to have most stained portions in the interstitial space of all the salt-treated groups, much stronger than the Control group.
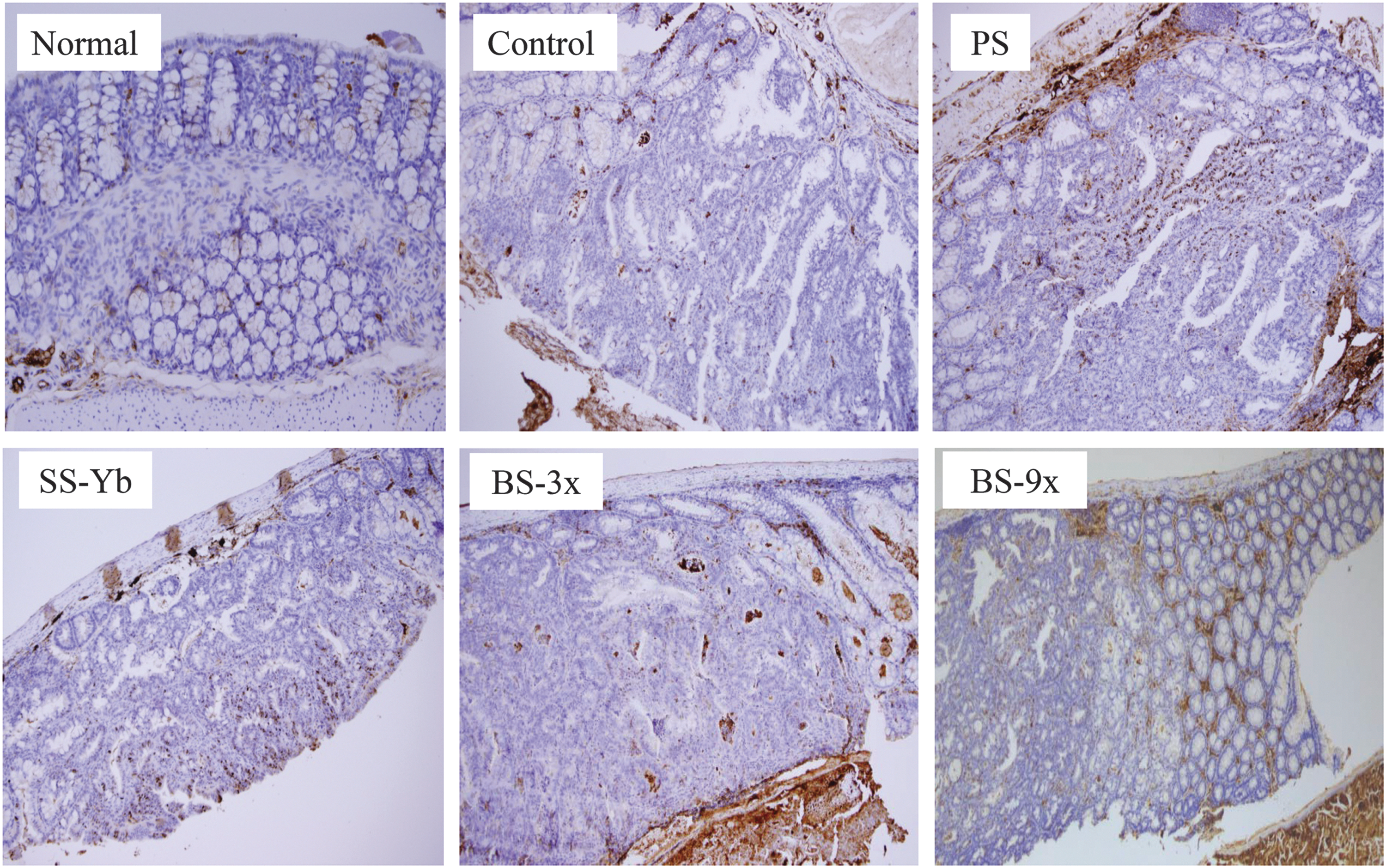
Immunohistochemical analysis of the colonic mucosae of mice with AOM/DSS-induced colon cancer, using p21 antibody. The brown-colored nuclei in the figure indicate areas of p21 expression. Group abbreviations are as described in the legend of Figure 1. Color images available online at
Discussion
Induction of colon cancer by AOM and DSS was found to shorten the colon length, cause edema, change morphology, harden the colon tissue, and to eventually increase colon weight and tumor counts. 14 The lower the colon weight-to-length ratio, the better prevention induced by suppression of tumorigenesis. Actually, our observations showed that increased baking times for processing the solar salt into BS reduced the ratio and suppressed the increased tumor counts induced by AOM and DSS. Additionally, BS is reported to have antioxidative efficacy owing to higher OH and potassium concentrations than other types of salts, which presumably means stronger cancer prevention. 3,7 It has also been reported that AOM-DSS-based colon cancer induction leads to decreased body weight during the DSS phase. 14 Our observations also showed that the average mouse body weight of the Normal group constantly increased throughout the entire experimental period due to the absence of AOM/DSS treatment, whereas body weights of all AOM/DSS-treated groups were found to decrease at week 5 when the second DSS session began. This effect was ameliorated by administration of BS3x or BS9x, whereas treatment with other salts (PS and SS-Yb) failed to ameliorate AOM-DSS-driven weight loss (data not shown).
Inflammation of the colon mucosa, found during inflammatory bowel disease (IBD), results from hyperactive immune cells, which boost proinflammatory cytokines, including TNF-α, IL-6, and IL-1β, to damage colon tissues, all of which are important contributors to inflammation. 15 Additionally, the cytokines tend to increase in the AOM-DSS-driven colon cancer model due to preinitiation of colitis-related colon cancer by AOM before DSS administration. 16 Thus, lower proinflammatory cytokine levels may reflect better efficacy against inflammation-related cancer. Based upon the facts referenced above, increased baking times for BS from SS may strengthen the anti-inflammatory efficacy when compared with that of SS-Yb.
Colon tissues are found to have severe destruction, inflammatory cell infiltration, and ulceration in the DSS-driven IBD model, all of which can be accelerated by pretreatment with AOM before administration of DSS for induction of IBD-related colorectal cancer. 14 The enteric tissues of the Control group showed loss of gland cells, tumors on the mucus layer, and infiltration of cancer cell into the tissues; however, the bamboo salt-treated groups showed suppression of such pathological changes, whereas PS and SS-Yb did not suppress these changes when compared with the Control group. In particular, increases in the baking cycles of BS were found to restore enteric tissues to close to those in the Normal group. Additionally, BS is reported to have more hydroxyl functional groups and more antioxidative effects than purified and solar salts. 8,9 Such special properties of BS might have partially contributed to the anti-inflammatory efficacies.
Apoptosis results from the expression of multiple factors, such as the Bcl family, which is known to play an important role. The major factors in the Bcl family include Bcl-2, Bcl-xl, Bax, and Mcl-1; of these factors, Bax promotes apoptosis, whereas Bcl-2 suppresses it, and both determine survival or death of cells under tight molecular control, including dimerization. 17 Based upon these facts and our results above, the bamboo salt-treated groups were found to suppress the expression of Bcl-2, while promoting expression of Bax, and such efficacy was found to be reinforced as the bamboo baking times were lengthened, whereas the efficacies of PS and SS-Yb could hardly approach those of BS treatment. Thus, tumors may undergo more frequent apoptosis as the baking time of BS is increased. Such results shown by RT-PCR were similarly reflected in H&E staining, showing that BS9x was the strongest stimulator of apoptosis induction of all the salt-treated experimental groups.
p53 is known to be important for cell cycle management, DNA recovery, and even apoptosis. p21 is well known as a suppressor of cyclin-dependent kinase; it suppresses tumor progression by halting the G1 to S and G2 to M phase transitions when DNA damage is detected. 18 In addition, it is also known that deactivation of p53 results in suppression of p21 and Bax. 19,20 Based upon these facts, the bamboo-treated groups were found to have partial anticancer effects presumably by such cell cycle arrest mechanism that promotes p53 and p21. This efficacy was strengthened as the baking times of the BS from SS-Yb increased, whereas PS and SS-Yb fell short of the BS-9x in p53- and p21 expression. Additionally, our observation of p21 expression by immunohistochemical assay showed that administration of BS9x strongly promoted expression of p21 in comparison with Control, whereas PS and SS-Yb fell behind BS-9x, reflecting the results obtained from RT-PCR.
In conclusion, supplementation with BS suppressed shortening of the colon length, reduced the colon weight-to-length ratio and tumor counts, and reduced the serum concentrations of proinflammatory cytokines, such as TNF-α, IL-6, and IL-1β, presumably resulting from reduced enteric expression of inflammation-related factors and modulation of apoptosis- and cell cycle-related factors (Bcl-2, Bax, p21, and p53), all of which were reflected by histopathological and immunohistochemical observations. Furthermore, the increase in baking times of BS promoted its anticancer efficacy.
Footnotes
Acknowledgment
This work was supported by grant 20130290 to Solar Salt Research Center of Mokpo National University from Ministry of Oceans and Fisheries of Korea.
Author Disclosure Statement
No competing financial interests exist.