
Abstract
The antiobesity effect of 5,7-dimethoxyflavone (DMF) was evaluated in 3T3-L1 adipocytes and high-fat diet (HFD)-induced obese C57BL/6J mice. The accumulation of lipid droplets and triglycerides in adipocytes was dose dependently suppressed by DMF through inhibition of adipogenesis. DMF downregulated the adipogenic transcription factors (peroxisome proliferator-activated receptor [PPAR]γ, CCAAT/enhancer binding protein [C/EBP]α, and sterol regulatory element-binding protein-1c [SREBP-1c]) and lipid synthesis enzymes (fatty acid synthase [FAS], acetyl-CoA carboxylase [ACC], lipoprotein lipase [LPL], and HMG-CoA reductase [HMGR]). AMP-activated protein kinase (AMPK) and AMPK related lipolytic proteins in differentiated adipocytes were activated by DMF. In the animal model, oral administration of DMF (50 mg/kg/day for 6 weeks) significantly decreased body weight gain without affecting food intake. Elevated serum levels of total cholesterol and low-density lipoprotein cholesterol were suppressed by DMF. Fat pad masses were reduced in DMF-treated obese mice, as evidenced by reduced adipocyte size. DMF altered the expression of adipogenic transcription factors in epididymal fat tissue. In addition, DMF attenuated HFD-induced nonalcoholic fatty liver disease by decreasing hepatic triglyceride accumulation. Overall, these results suggest that DMF is a potential natural agent for attenuating obesity and other obesity-related metabolic syndromes.
Introduction
O
On the cellular level, obesity is defined as the increase in adipocyte size (hypertrophy) and number (hyperplasia). 3 Adipogenesis is the differentiation process converting preadipocytes to mature adipocytes, which is the essential stage of the onset of obesity. The start of adipogenesis is caused by the change of the expression of transcriptional factors in preadipocytes. 6 The master adipogenic transcription factors, including peroxisome proliferator-activated receptor (PPAR)γ, CCAAT/enhancer binding protein (C/EBP), and sterol regulatory element-binding protein-1c (SREBP-1c), are mostly expressed during adipogenesis. C/EBPβ and C/EBPδ are rapidly expressed in the early phase of differentiation and induce the expression of PPARγ and C/EBPα. These two adipogenic transcription factors are necessary for the terminal adipocyte differentiation and upregulate subsequent adipocyte-specific genes involved in adipogenesis. 6 –8
Activation of AMP-activated protein kinase (AMPK) leads to the inhibition of adipocyte differentiation. The AMPK downregulates lipogenic enzymes, such as fatty acid synthase (FAS), acetyl-CoA carboxylase (ACC), lipoprotein lipase (LPL), and HMG-CoA reductase (HMGR). In contrast, AMPK upregulates lipolytic proteins, including PPARα, PPARγ-coactivator-1α (PGC-1α), and uncoupling proteins (UCPs), which are positively related to fatty acid oxidation. 1,9,10 Thus, AMPK is a promising target by which obesity can be attenuated. 11
5,7-Dimethoxyflavone (DMF), which is isolated from Kaempferia parviflora Well. Ex Baker (commonly referred to as black ginger or Thai ginseng), has been reported to possess anti-inflammatory, anticancer, anticholinesterase, and melanogenic properties. 12 –15 K. parviflora contains abundant amounts of various types of methoxyflavones and polymethoxyflavonoid (PMF)s. Among them, DMF is present at 21.68 mg/g as a major methoxyflavone component. 12 DMF did not show a significant effect on adipogenesis of 3T3-L1 cells in previous reports, 16,17 which is needed to clarify antiobesity effect and its mechanisms' action. Thus, this study investigated the antiobesity effect of DMF by inhibiting adipogenesis in 3T3-L1 adipocytes and high-fat diet (HFD)-induced obese C57BL/6J mice.
Materials and Methods
Isolation of DMF from K. parviflora
Dried rhizomes of K. parviflora were collected in Bangkok, Thailand. A voucher specimen was deposited in the Department of Biotechnology, Yonsei University (Seoul, Korea). DMF (Fig. 1) was isolated from the ethanol extract of K. parviflora and identified as described previously. 15 The purity of DMF was determined to be ≥ 98% by high performance liquid chromatography analysis.
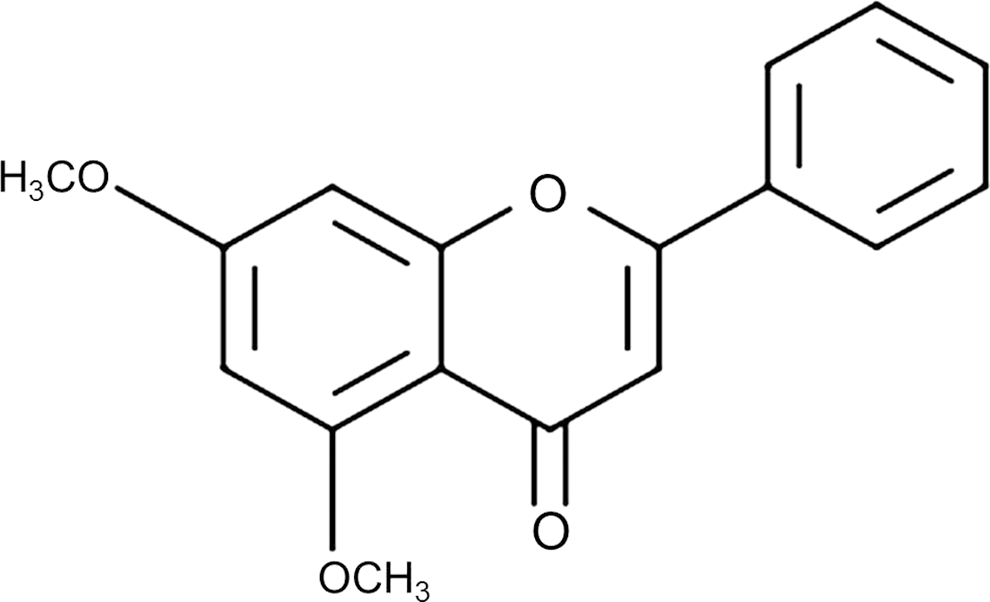
Chemical structure of 5,7-dimethoxyflavone.
Cell culture and differentiation
3T3-L1 preadipocytes were obtained from American Type Culture Collection (ATCC; Manassas, VA, USA) and grown in Dulbecco's modified Eagle's medium (DMEM; HyClone, Logan, UT, USA) supplemented with penicillin (120 U/mL), streptomycin (75 μg/mL), and 10% bovine calf serum (BCS; HyClone). Cells were maintained in an atmosphere of 5% CO2 at 37°C. For the induction of differentiation, 3T3-L1 preadipocytes were incubated with differentiation medium (MDI; 0.5 mM 3-isobutyl-1-methylxanthine (IBMX; Sigma-Aldrich, St. Louis, MO, USA), 0.2 μM dexamethasone (DEX; Sigma-Aldrich), and 1.7 μM insulin (Sigma-Aldrich) in DMEM containing 10% fetal bovine serum (FBS; HyClone)) for 2 days. The medium was replaced with DMEM containing 10% FBS and 1.7 μM insulin, which were changed every 2 days for 8 days. To examine the effect of DMF on adipogenesis, the cells were cultured with differentiation medium in the presence of DMF. DMF was used at various concentrations (5, 10, 20, 40, and 80 μM); no significant cytotoxicity was observed at concentrations up to 40 μM (data not shown). All studies were performed with less than 20 μM DMF. For AMPK inhibition experiments, compound C (AMPK inhibitor; Calbiochem, San Diego, CA, USA) was used. Before the induction of differentiation, 3T3-L1 cells were treated with 10 μM of compound C or serum-free medium for 30 min.
Oil Red O staining
Eight days following adipocyte differentiation induction, the 3T3-L1 cells were washed with phosphate-buffered saline (PBS) and fixed with 10% buffered formalin and stained with Oil Red O solution (0.5 g in isopropanol) for 1 h. After removing the staining solution, the stained lipid droplets were visualized in the cells using an inverted microscope (Nikon TE-200 U, Tokyo, Japan). Quantitation was carried out by extracting the Oil Red O-stained lipid droplets with 100% isopropanol and measuring the absorbance of each solution at 540 nm with a VersaMax tunable microplate reader (Molecular Devices, Inc., Sunnyvale, CA, USA).
Triglyceride content
For quantification of triglyceride (TG) contents, 3T3-L1 adipocytes and homogenized liver tissues were lysed using NP-40 buffer (Elpis Biotech, Daejeon, Korea). The TG contents in each sample were, respectively, measured with a TG Quantification Kit (BioVision, Mountain View, CA, USA) according to the manufacturer's instructions.
Western blot analysis
3T3-L1 adipocytes and homogenized epididymal adipose tissues were lysed using NP-40 buffer (Elpis Biotech) containing a protease inhibitor cocktail (Sigma-Aldrich). The primary antibodies were used against PPARγ, C/EBPα, SREBP-1c, AMPK, phosphorylated AMPK (Thr172), α-tubulin (1:1000 dilution; Cell Signaling, Beverly, MA, USA), phosphorylated ACC, FAS, PPARα, PGC-1α, UCP2, and UCP3 (1:1000 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA). Bound antibodies were detected using goat anti-rabbit or goat anti-mouse horseradish peroxidase-conjugated IgG secondary antibodies (1:5000 dilution; Bethyl Laboratories, Inc., Montgomery, TX, USA) for 2 h. Signals were detected with the enhanced chemiluminescence (ECL) detection system (Amersham Biosciences, Little Chalfont, United Kingdom) and visualized with the G:BOX EF imaging system (Syngene, Cambridge, United Kingdom) and GeneSnap software. α-Tubulin was used as an internal control.
Reverse transcription-polymerase chain reaction
Total RNA was extracted from 3T3-L1 adipocytes and homogenized epididymal adipose tissues using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and then converted to cDNA with reverse transcriptase premix (Elpis Biotech). To quantify mRNA expression, the cDNA was amplified with HiPi PCR PreMix (Elpis Biotech) and primer pairs (Bioneer, Daejeon, Korea) (Table 1). All primers were denatured at 94°C for 5 min before performing PCR amplification. Amplification consisted of 25–30 cycles: denaturing at 94°C for 30 s, annealing at 50–63°C for 1 min, and extending at 72°C for 1 min, followed by a final extending phase at 72°C for 5 min. PCR was performed in a GeneAmp PCR System 2700 (Applied Biosystems, Foster City, CA, USA). PCR products were separated by electrophoresis on 1.5% agarose gels and visualized with the G:BOX EF imaging system (Syngene) and the GeneSnap software. β-Actin was used as an internal control.
Animal experiments
Four-week-old male C57BL/6J mice (Orient Bio, Inc., Seongnam, Korea) were housed in a controlled environment (25°C ± 2°C and 55% ± 5% relative humidity with a 12-h light/12-h dark cycle) in the Yonsei Laboratory Animal Research Center (YLARC; Seoul, Korea). Throughout the experiment, the mice were allowed free access to food and tap water. After 1 week of acclimatization, eighteen mice were randomly divided into three experimental groups (each group, n = 6). Group 1 received a normal chow diet (ND) (Rodent diet D12450B; Research Diets, New Brunswick, NJ, USA; 10% fat, 70% carbohydrate, 20% protein; 3.85 kcal/g); group 2 received a HFD (Rodent diet D12492; Research Diets; 60% fat, 20% carbohydrate, 20% protein; 5.24 kcal/g); and group 3 received HFD with the 50 mg/kg/day DMF (HFD+DMF). Obesity was induced in groups 2 and 3 by HFD feeding for 6 weeks before the 6 weeks of oral sample administration. DMF was dissolved in saline, and animals were orally administrated with DMF at a dose of 50 mg/kg/day using oral feeding needles for 6 weeks. Oral administration volume was approximately 200 μL per mouse. Mice in the ND group and HFD group were given an equal volume of saline. Food intake and body weight were measured twice a week throughout the experiment. At the end of the oral administration period, all mice were sacrificed using CO2 gas. Epididymal and perirenal fat pads were removed, measured, and frozen in liquid nitrogen for storage at −70°C. This study adhered to the Guide for the Care and Use of Laboratory Animals developed by the Institute of Laboratory Animal Resources of the National Research Council. All work was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of YLARC (Permit number is IACUC-201401-120-01).
Micro-CT imaging
Micro-computed tomography (micro-CT) experiments were performed with an animal positron emission tomography (PET)/CT/single photon emission computed tomography (SPECT) system (INVEON, Siemens, Washington, DC, USA) at the Center for Evaluation of Biomaterials at Pohang Technopark.
Analysis of blood biochemical parameters
Whole blood samples were collected from mice by heart puncture. Serum was prepared by centrifugation at 1500 g for 15 min and stored at −70°C until analysis. Serum lipid profiles and serum levels of hepatotoxicity markers were analyzed using an automated biochemical analyzer (Mindray, Shenzhen, China) according to the method established by the International Federation of Clinical Chemistry (IFCC).
Histological analysis
After sacrificing the mice, epididymal adipose and liver tissues were fixed in 10% formalin for 24 h and stained with hematoxylin and eosin (H&E). The stained sections were analyzed using an Eclipse TE2000 U inverted microscope equipped with twin CCD cameras (magnification, × 200; Nikon, Tokyo, Japan). Adipocyte size was quantified using ImageJ software, version 1.47 (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All experiments were repeated at least thrice. Results are presented as mean ± standard deviation (SD). Statistical analysis was performed using SPSS 21.0 (SPSS, Inc., Chicago, IL, USA). One-way analysis of variance (ANOVA) followed by a Scheffe's test for in vitro testing and group differences was assessed by unpaired Student's t-test for in vivo test. P values <.05 were considered statistically significant.
Results
Effect of DMF on differentiation of 3T3-L1 adipocytes
DMF significantly inhibited lipid accumulation by reducing TG accumulation in 3T3-L1 adipocytes (Fig. 2A, B). PPARγ, C/EBPs, and SREBP-1c are the master transcription factors in the regulation of adipogenesis. 18,19 DMF effectively downregulated PPARγ, C/EBPα, and SREBP-1c protein and mRNA expression levels (Fig. 2C, D). In addition, DMF reduced the mRNA expression of adipocytokines such as adiponectin (Fig. 2E). These results indicated that DMF suppresses adipocyte differentiation through inhibition of the transcriptional factors involved in the differentiation.
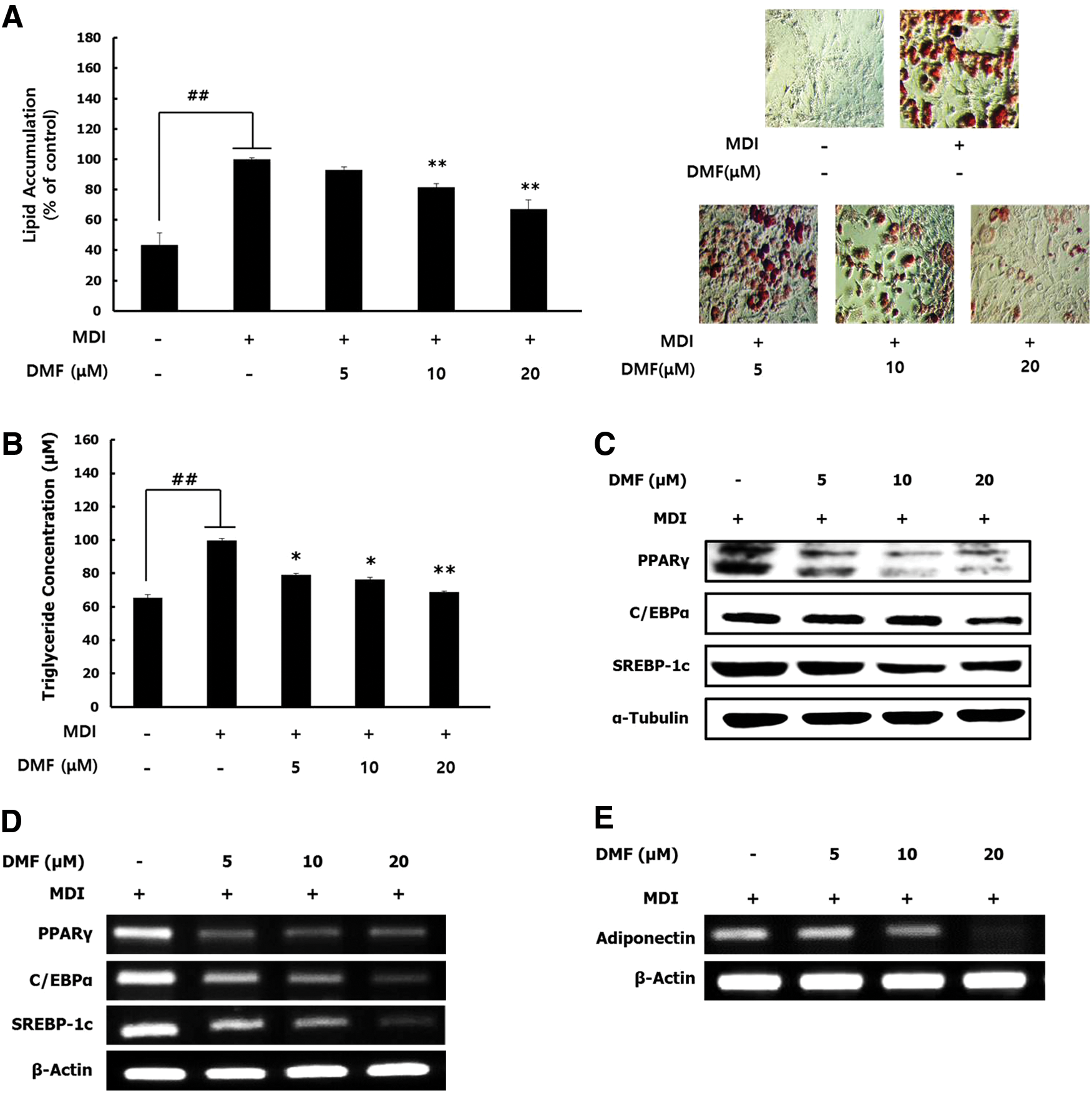
Effect of DMF on lipid accumulation and the expression of differentiation-related transcriptional factors in 3T3-L1 adipocytes. The cells were cultured with the differentiation medium in the various concentrations of DMF for 8 days.
Effect of DMF on lipid metabolism-related enzymes through AMPK activation in 3T3-L1 adipocytes
To test whether DMF affects the AMPK activity, 3T3-L1 adipocytes were treated with DMF in the presence or absence of compound C, an AMPK inhibitor. The lipid content was decreased by DMF treatment and this effect was significantly reversed in the presence of compound C (Fig. 3A). DMF effectively increased AMPK and ACC phosphorylation, while no change was observed in AMPK and ACC total protein expression (Fig. 3B). DMF reduced the mRNA expression of lipogenic enzymes such as LPL, ACC1, FAS, and HMGR (Fig. 3C). In addition, DMF upregulated the PPARα, PGC-1α, and UCPs and mRNA expression levels (Fig. 3D, E), which are lipolytic proteins involved in fatty acid oxidation. 9 These results suggest that DMF modulates lipid metabolism and increases fatty acid oxidation through the AMPK signaling pathway in 3T3-L1 adipocyte.
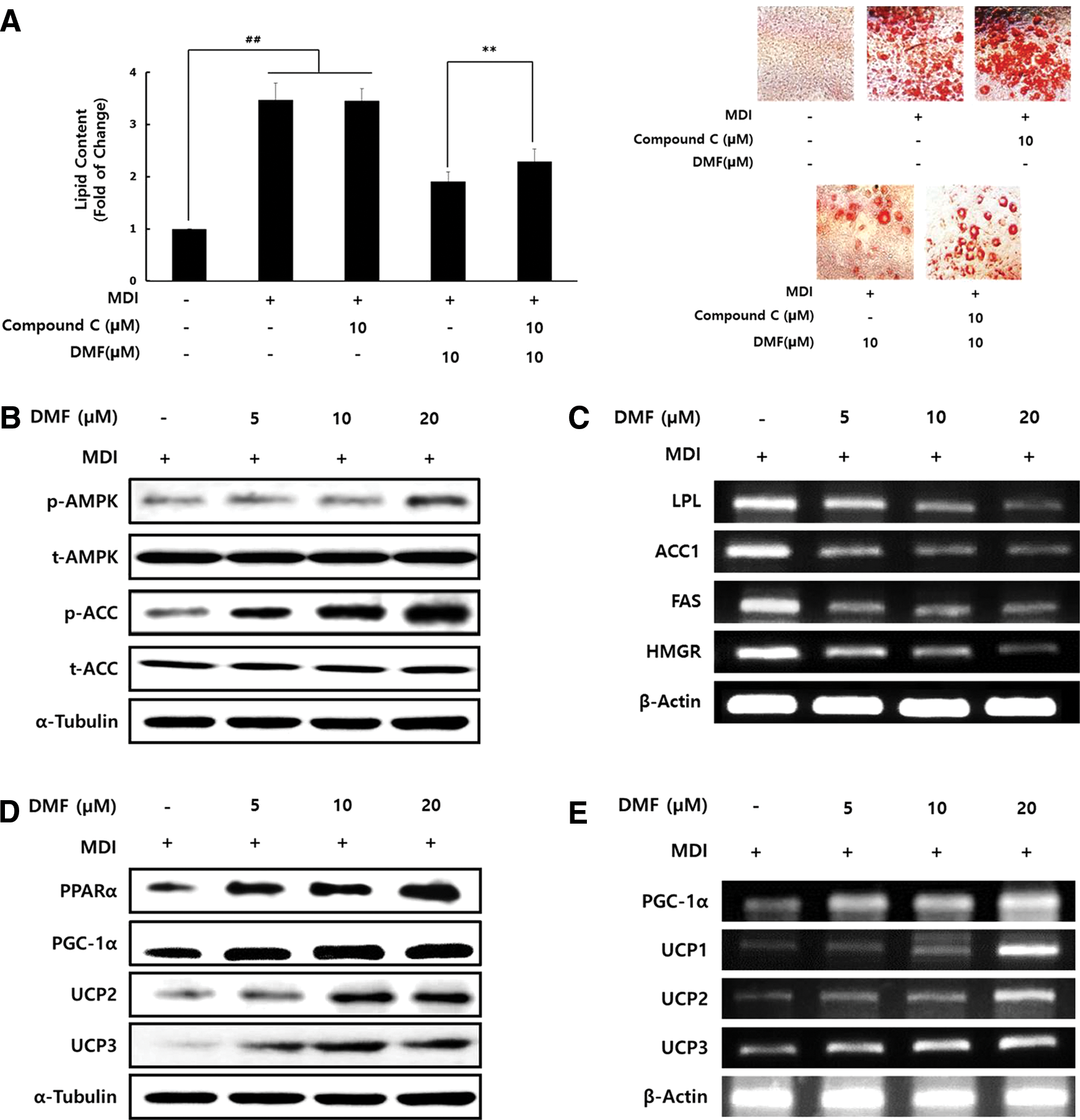
Effect of DMF on AMPK activation and lipid metabolism-related enzymes in 3T3-L1 adipocytes. The cells were cultured in the differentiation medium with various concentrations of DMF for 8 days.
Effect of DMF on HFD-induced body weight gain and adiposity in mice
The body weights of the HFD group increased markedly more compared with those of the ND group; this difference was gradually increased until 12 weeks. The HFD+DMF group showed consistently less body weight gain without affecting food intake, compared with the HFD group (Fig. 4A). Gross appearance and micro-CT imaging showed that body shape and body fat composition were remarkably smaller in HFD+DMF group than in the HFD group (Fig. 4B, C). These results indicate that the oral administration of DMF reduces body weight gain and body size by decreasing whole body fat accumulation without changing food intake.
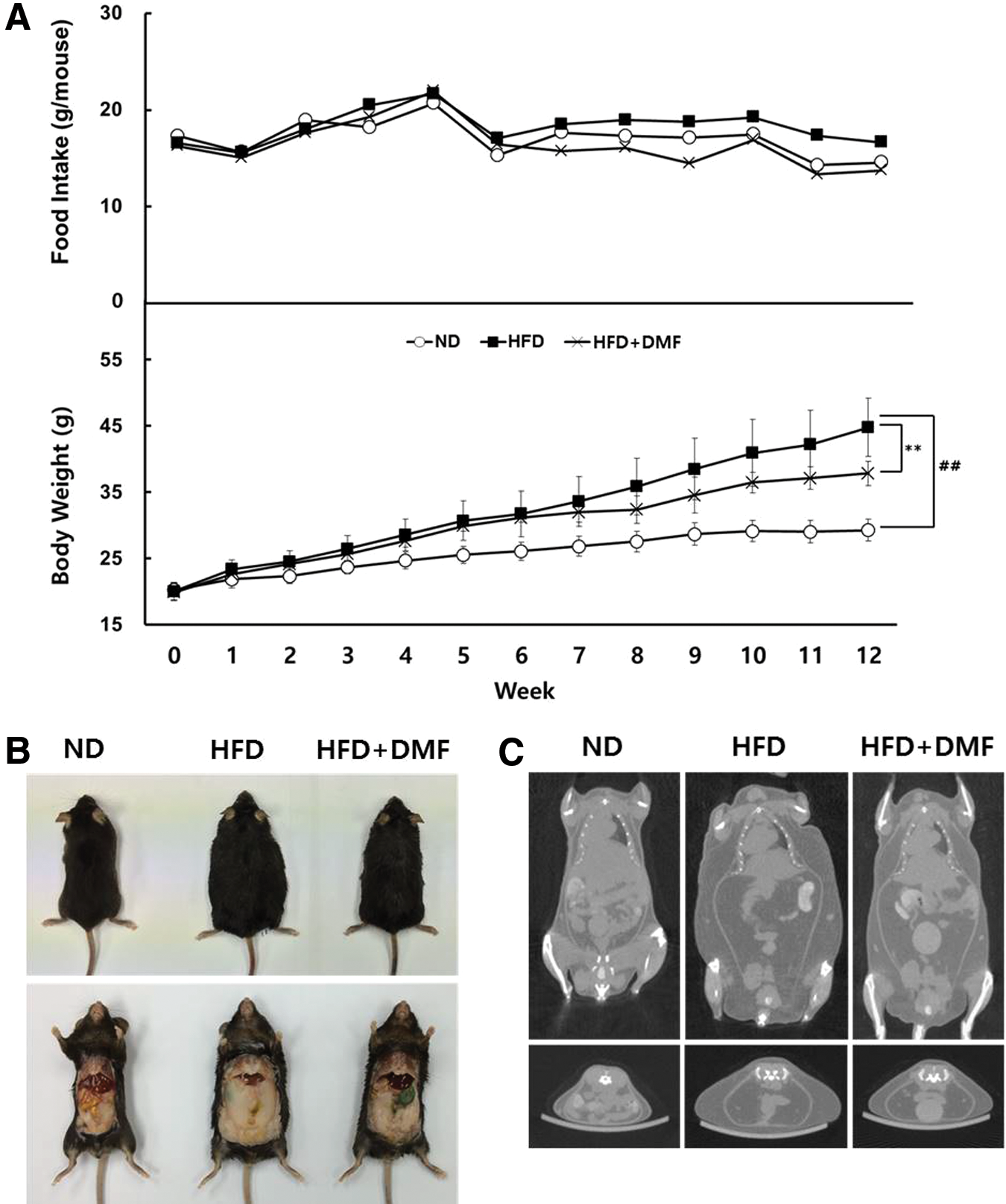
Effects of DMF treatment on body weight gain and adiposity in HFD-induced obese mice.
Effect of DMF on HFD-induced hypercholesterolemia in mice
The HFD group showed the increase of total cholesterol (Total-C) and low-density lipoprotein cholesterol (LDL-C) compared with the ND group and the parameters were effectively lowered in the HFD+DMF group. The level of high-density lipoprotein cholesterol (HDL-C) was not significantly different among all groups (Fig. 5A). The ratio of HDL-C/Total-C was significantly lower in the HFD group, but was recovered in the HFD+DMF group (Fig. 5B). Furthermore, the levels of serum hepatotoxic markers, including AST and ALT, were not significantly altered (Fig. 5C). These results show that DMF prevents HFD-induced hypercholesterolemia and the risk of other related diseases by decreasing LDL-C and Total-C without hepatotoxicity.
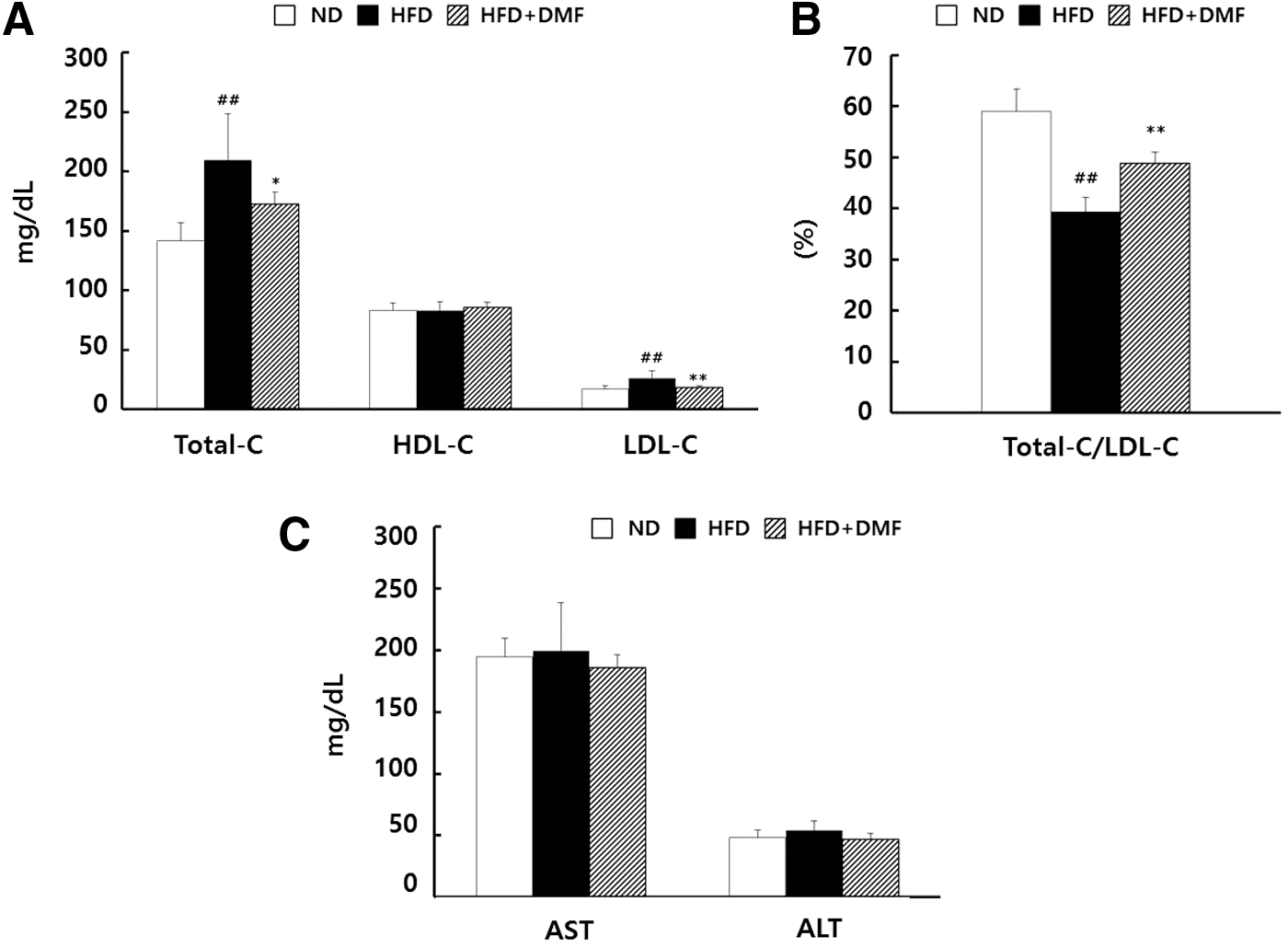
Effects of DMF treatment on serum lipid profiles and hepatotoxicity markers in HFD-induced obese mice.
Effect of DMF on HFD-induced fat mass, fat size, and the expression of adipogenic transcriptional factors in mice
Epididymal and perirenal fat pad masses were increased in HFD group compared to ND group, but were decreased in HFD+DMF group compared with HFD group (Fig. 6A). Histological analysis of epididymal fat indicated that the fat pad size was increased in the HFD group, but significantly decreased in the HFD+DMF group (Fig. 6B). Furthermore, the protein expression of major adipogenic transcriptional factors, including PPARγ, C/EBPα, and SREBP-1c, was decreased in HFD+DMF group compared with HFD group (Fig. 6C). Taken together, the data indicate that the DMF-mediated decrease in body weight gain is related to the decrease of fat masses and fat size, due to modulation of the adipogenic transcriptional factors.
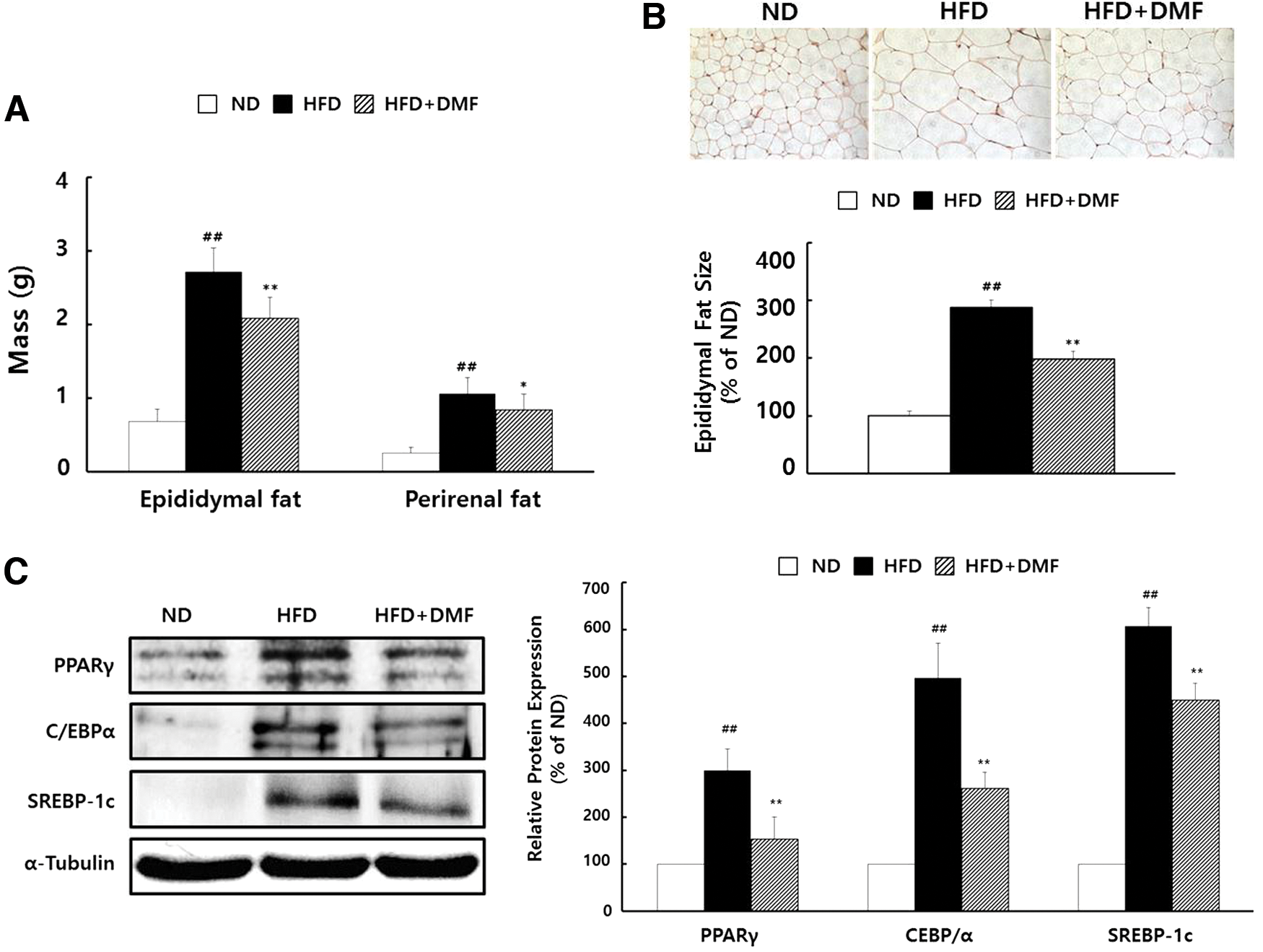
Effects of DMF treatment on fat pad masses, adipose tissue histology, and the expression of adipogenic transcriptional factors.
Effect of DMF on HFD-induced nonalcoholic fatty liver disease in mice
Liver weight was higher in the HFD group and lower in HFD+DMF group; similar results were obtained regarding the TG content and lipid droplet size in liver (Fig. 7A, B). These results indicate that DMF alleviated HFD-induced fatty liver by decreasing liver TG content and lipid droplet size in liver.

Effects of DMF treatment on liver mass, histological analysis, and TG contents.
Discussion
Previously, it was reported that DMF did not induce the differentiation of 3T3-L1 cells and did not affect the suppression of intracellular lipid accumulation in mature adipocytes. 16,17 In contrast, this study showed that DMF significantly decreased the accumulation of intracellular lipid during adipogenic differentiation (Fig. 2A). This difference may be due to different state of adipocyte differentiation when DMF was used to treat the 3T3-L1 cells. PPARγ has been suggested to be a master regulator of adipogenesis and is necessary for adipocyte differentiation. 20 Activation of PPARγ immediately leads to the induction of C/EBPα and SREBP-1c expression, which subsequently activates transcription of a series of adipocyte-specific genes responsible for terminal differentiation. 21 In present study, DMF downregulated adipogenic transcription factor expression in both 3T3-L1 adipocyte (Fig. 2C, D) and epididymal adipose tissue (Fig. 6C), which led to suppressed accumulation of intracellular lipid droplets (Fig. 2A) and reduced body weight gain by decreasing the fat mass and fat size (Figs. 4A and 6A, B). Adiponectin is one of the multiple adipocytokines, which led to accelerated adipocyte differentiation by upregulating the expression of PPARγ, C/EBPα, and SREBP-1c. 22 Antiadipogenic effect of DMF by decreasing the expression of PPARγ, C/EBPα, and SREBP-1c may be involved in the reduced mRNA expression of adiponectin (Fig. 2E). These results indicate that the antiobesity ability of DMF is mediated by its inhibition of adipogenesis.
AMPK plays an important role by directly phosphorylating the downstream target ACC at Ser79. This phosphorylation event inhibits its lipid biosynthesis enzymatic activity and causes a subsequent decrease in fatty acid synthesis. 11 In addition, AMPK increases thermogenesis by inducing PGC-1α and UCPs. 23 DMF significantly increased AMPK and ACC phosphorylation (Fig. 3B), which was accompanied by decreased lipid accumulation and increased thermogenesis in differentiated 3T3-L1 adipocytes. In addition, 3T3-L1 preadipocytes were not able to develop into mature adipocytes in the presence of DMF, but this effect was reversed when the cells were pretreated with compound C, an AMPK inhibitor (Fig. 3A). These results indicate that DMF inhibits adipocyte differentiation and lipid accumulation through the AMPK pathway.
DMF also resulted in decreased weight gain, decreased fat mass and fat size, lower levels of LDL-C and Total-C, and reduced liver weight and hepatic lipid accumulation in obese mice without reducing appetite and toxicity (Figs. 4 –7). Toda et al. 24 reported that DMF significantly enhanced the mRNA expression of glucose transporter 4 (GLUT4) in C2C12 skeletal muscle cells, leading to the improvement of insulin stimulated glucose uptake. Akase et al. 25 also reported that K. parviflora extract (KPE), containing DMF as a major bioactive compound, effectively attenuated obesity and insulin resistance by suppressing fat accumulation, hyperinsulinemia, glucose intolerance, hypertension, and insulin resistance in obese type 2 diabetic mice. These results indicate that DMF can improve obesity-related metabolic dysfunctions and insulin sensitivity in obese mice.
Serum levels of hepatotoxic biomarkers were not significantly altered among the groups, which indicated safety probability of DMF (Fig. 5C). Higher hepatic fat accumulation induced by HFD leads to increased hepatotoxic biomarker, which causes fatty liver disease and hepatic inflammation and results in imbalance of lipid metabolism. 26 DMF leads to decreased liver triglyceride content, lipid droplet size, and liver weight (Fig. 7), which protected against obesity-induced hepatic injury. These results indicate that DMF can potentially be used to treat nonalcoholic liver disease.
The present study shows that DMF exerts an antiobesity effect by suppressing adipogenesis. DMF effectively attenuated lipid and TG accumulation in 3T3-L1 cells and decreased body weight gain, fat masses, and fat size in obese mice. Furthermore, DMF improved serum lipid profiles in obese mice and exerted hypolipidemic effects on hepatocytes without any liver damage. Taken together, our data suggest that DMF is a novel candidate for treating HFD-induced obesity.
Footnotes
Acknowledgment
This research was supported by the Commercializations Promotion Agency for R&D Outcomes (COMPA) funded by the Ministry of Science, ICT and Future Planning (MSIP) (2016K000107).
Author Disclosure Statement
The authors declare that there are no conflicts of interest.