
Abstract
Uncaria rhynchophylla is one of the major components included in Traditional Chinese Medicine prescriptions for hypertensive treatment. Previous studies have suggested that U. rhynchophylla might contain vasodilation-mediating active compounds, especially indole alkaloids. Hence, this study was carried out to determine the vasodilatory effects of U. rhynchophylla, which was extracted by different solvents. The most effective extract was then further studied for its signaling mechanism pathways. The authenticity of U. rhynchophylla was assured by using modernized tri-step Fourier transform infrared (FTIR), including conventional 1D FTIR, second derivative scanning combined with 2D-correlated IR spectroscopy. Results obtained proved that the fingerprint of U. rhynchophylla used was identical to the atlas. Isolated aortic rings from male Sprague–Dawley rats were preconstricted with phenylephrine (PE) followed by cumulative addition of U. rhynchophylla extracts. The signaling mechanism pathways were studied by incubation with different receptor antagonists before the PE precontraction. In conclusion, the 95% ethanolic U. rhynchophylla extract (GT100) was found to be most effective with an EC50 value of 0.028 ± 0.002 mg/mL and an R max value of 101.30% ± 2.82%. The signaling mechanism pathways employed for exerting its vasodilatory effects included nitric oxide/soluble guanylyl cylcase/cyclic guanosine monophosphate (NO/sGC/cGMP) and PGI2 (endothelium-derived relaxing factors), G protein-coupled M3- and β 2 receptors, regulation of membrane potential through voltage-operated calcium channel, intracellular Ca2+ released from inositol triphosphate receptor (IP3R), and all potassium channels except the Kca channel.
Introduction
H
As stated in the hypertension Clinical Practice Guideline (CPG), combination drug therapy is more effective than monotherapy because it targets multiple signaling pathways. Therefore, the discovery of new drugs that are capable of employing multiple vasodilative-mediated signaling mechanism pathways is highly desired. 2 Traditional Chinese Medicine (TCM) could serve as a resource for pharmaceutical research. The therapeutic values of TCM established since the beginning of human civilization have inspired researchers to continue the discovery of hidden medicinal values of herbs. 3
In TCM, one of the important ingredients prescribed for the treatment of cardiovascular diseases 3 and hypertension 4 –6 is Uncaria rhynchophylla. The ancient medical practitioners created TCM prescriptions for treating hypertension by utilizing the stems and hooks of U. rhynchophylla, which were known as Tianma gouteng yin, Gouteng tang, Gouteng san, and Gouteng yin in Chinese herbal medicine. 5 The Uncaria genus is widely distributed in tropical regions, especially in South America, Africa, and Southeast Asia. 4 TCM principles defined the disharmony patterns of hypertension according to the Chinese Medicine Guidelines into several types, which include hyperactivity of liver-yang, yin deficiency and yang hyperactivity, deficiency of both yin and yang, and liver-wind with turbid phlegm. Generally, all these disharmony patterns are accompanied by a few other symptoms, including dizziness, distending headache, and fatigue.
According to the Compendium of Materia Medica, U. rhynchophylla is able to extinguish wind, clear heat, pacify liver, and arrest convulsion in treating hypertension. 5,7,8 There are more than 200 chemical constituents present in Uncaria species, which include triterpenes, flavonoids, phenylpropanoids, and indole alkaloids. 5 Currently, a total of 123 alkaloids have been discovered in plants of the Uncaria genus, which constitutes 0.2% of the total content. 5 Among these alkaloids, rhynchophylline, 9 isorhynchophylline, corynoxeine, 9 and isocorynoxeine are believed to contribute >50% of the total alkaloids found in Uncaria spp., especially in U. rhynchophylla. 10 –14 Studies also showed that among these four major alkaloids, rhynchophylline and isorhynchophylline contributed ∼28–50% and 5%, respectively, to the total alkaloid content in U. rhynchophylla. 5 The other alkaloids, namely corynantheine, dihydrocorynantheidine, corynoxeine, rhynchophylline, akuammigine, isorhynchophylline, isocorynoxeine, hirsuteine, and hirsutine, were found to constitute 0.09%, 0.21%, 0.67%, 0.79%, 0.89%, 1.60%, 1.63%, 2.01%, and 2.34%, respectively, of the total alkaloids of U. rhynchophylla. 4,11
Rhynchophylline and isorhynchophylline were suggested to be the main hypotensive constituents of U. rhynchophylla 6 , with pD2 values of 4 for both in vasodilation studies. 15 Current studies also showed that the vasodilatory effect of rhynchophylline involved the calcium and potassium-linked channels. 3 However, some researchers claimed that the anti-hypertensive effect of U. rhynchophylla is mostly contributed by cadambine, 3α-dihydrocadambine, and 3β-isodihydrocadambine rather than rhynchophylline. 14,16,17 In addition, corynoxeine, geissoschizine methyl ether, and villocarine A isolated from U. rhynchophylla showed vasorelaxant effects with pD2 values of 4.8, 6.13, and 4.52, 12,18 –20 respectively, in the aortic ring model. It is interesting to note that most of the bioactive compounds were present in the alcoholic extract of U. rhynchophylla. 21 Hence, this study was conducted to determine the vasodilatory effect of the U. rhynchophylla ethanolic extract, as we believe that either the alcoholic or aqueous extracts could induce a more potent vasodilatory effect than any of the single compounds.
According to Loh et al., studies on vasodilatory effects were most frequently conducted by using the isolated rat aortic ring model, which is known as the “golden tool” in pharmacological research due to its high reliability. 22,23 Therefore, this experimental model was utilized for the investigation on the signaling mechanism pathways of U. rhynchophylla's vasodilatory effects. On the other hand, tri-step Fourier transformed infrared (FTIR) spectroscopy, which includes conventional FTIR (1D-IR) and second-derivative scanning combined with 2D-infrared correlation analysis, was used as a rapid tool for authenticating the herb and analyzing the complicated mixture of components in U. rhynchophylla.
Materials and Methods
Plants and chemicals
The radix of U. rhynchophylla was purchased from the local medical hall. The authenticity of the purchased U. rhynchophylla was authenticated by Dr. Yam Mun Fei (expert in Chinese Materia Medica, Universiti Sains Malaysia). Phenylephrine (PE), acetylcholine (Ach), and nifedipine (Nif) were purchased from Acros Organics (Belgium); 4-Aminopyridine (4-AP) and potassium bromide (KBr) were purchased from Merck (Germany). Methylene blue (MB) and ethylene glycol tetraacetic acid (EGTA) were purchased from Promedipharm Sdn. Bhd (Malaysia) and Calbiochem (Germany), respectively. Tetraethylammonium (TEA), 1H-[1,2,4]oxadiazolo[4,3,-a]quinoxalin-1-one (ODQ), barium chloride (BaCl2), Nω-nitro-
Animals
The experiments were performed on male Sprague–Dawley rats between the ages of 8 to 10 weeks and weighing 180 to 250 g that were housed at room temperature with a 12-h light-dark cycle and granted free access to water and food. The investigation obeyed the Guideline in Care and Use of Laboratory Animal by Universiti Sains Malaysia [animal ethnic approval: USM/2016/(103)/(777)].
Preparation of extracts
U. rhynchophylla was sliced into 2–3 cm pieces and dried in the oven at a temperature of 50°C for 48 h. While drying, the weight of sliced U. rhynchophylla was measured daily until a constant weight was achieved and then the dried plant was ground into fine powder. About 200 g of ground powder was soaked in 95% ethanol, a mixture of 50–50% ethanol and distilled water, and distilled water, in a water bath at 50°C for 48 h. The maceration process was repeated three times. Filtration was carried out to separate the filtrate and residue. The filtrate was concentrated by using a rotary evaporator under reduced pressure, and the resultant extract was subsequently freeze dried. The yields of U. rhynchophylla water extract (GTW), 50% of ethanolic extract (GT50), and 95% of ethanolic extract (GT100) were 5.98%, 8.93%, and 5.09%, respectively. The stock extracts were kept in a desiccator at 4°C for future use.
Herb characterization by Tri-step FTIR
The experiment began with a tablet of KBr as blank. The herb samples were pulverized and passed through a 200-mesh sieve to obtain fine herb powder before the experiment. About 1 mg of herb powder was mixed evenly with 100 mg of KBr and pressed as a tablet at ∼10 psi. The spectra were recorded at 0.2 cm/s of optical path speed with 16 co-added scans at resolutions of 4 cm−1 between wavelengths 4000 and 400 cm−1 by using a Spectrum 400 FT-IR spectrometer (v 6.3.5) that was equipped with a deuterated triglycine sulfate (DTGS) detector (Perkin-Elmer, USA). The disturbances caused by carbon dioxide and water were eradicated online while scanning. The FTIR spectrum was assumed valid when a transmission of 60% or higher was achieved or else the test had to be repeated by adding more sample or KBr. 24
The second derivative IR (SD-IR) spectra were achieved after Savitzky–Golay polynomial fitting (13-point smoothing). The 2D-IR spectra were obtained by placing the sample tablet into the sample holder with a programmable heated jacket controller (Model GS20730; Specac). The dynamic spectra were recorded at temperatures ranging from 50°C to 120°C at an interval of 10°C. The 2D-IR correlation spectra were obtained by treating the series of dynamic spectra with 2D-IR correlation analysis software developed by Tsinghua University (Beijing, China). 24 –26
Preparation of aorta rings
A petri dish filled with Krebs–Henseleit (Krebs') solution (118.0 mM NaCl, 4.7 mM KCl, 25.0 mM NaHCO3, 1.25 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, and 11.0 mM glucose, pH 7.4) was prepared and aerated with carbogen (95% O2 and 5% of CO2). A male Sprague–Dawley rat was euthanized by an overdose of CO2 inhalation. An incision was made through its sternum to locate the aorta. The aorta was excised and placed in the petri dish containing the Krebs' solution immediately. The excessive blood, connective tissues, and fat adhering to the aorta were removed. After that, the aorta was cut into 3–5 mm sections and mounted carefully within a tissue chamber containing 10 mL of standard Krebs' solution by using a small hook and an L-shaped brace. Throughout the experiment, the chamber was continuously aerated with carbogen gas at 37°C. One of the hooks was mounted at the bottom of the tissue chamber whereas another was connected to the force-electricity transducer (GRASS Force-Displacement Transducer FT03 C Isometric Force Measurements). The suspended aortic rings were left to equilibrate for 45 min at a tension of 1.0 g. The Krebs' solution was replaced every 15 min, and the tension was readjusted back to 1.0 g if necessary. The validity of the aortic rings was assured by exposure to the contractile agent, PE (1 μM) followed by the relaxing agent, Ach (1 μM). Validity was assured with at least 60% vasoconstriction and 60% vasorelaxation responding to their respective agonist. After that, the rings were rinsed with Krebs' solution for three times so that the tension returned to baseline. Subsequently, PE was applied to induce precontraction. The herb extract (100 μL) was then added cumulatively (0.0025–0.08 mg/mL, which is equal to a final concentration of 0.1575 mg/mL) into the tissue chamber at 20 min intervals. 27 Three types of herb extracts (GTW, GT50, and GT100) were tested. Concentration-response curves were constructed based on the contractile response measured by the force-electricity transducer. The signals were amplified by Quad bridge amp (AD instrument, Australia) and converted into digital signals by powerLab 26T (AD instrument). At this stage, the extract that exhibited the highest vasodilatory effect was subjected to further mechanism studies. 27,28
Investigations of endothelium dependency of GT100's vasodilatory effects
Mechanism studies of vasodilatory effects usually start by determining the endothelium dependency. After isolating the aorta from the Sprague–Dawley rat, the endothelium was mechanically removed by gently rubbing the lumen of the aorta with a stick. The complete removal of the endothelium was confirmed by exposing it to Ach, which caused no relaxation. The concentration-response curve was constructed by the cumulative addition of GT100 extract (100 μL of 0.25–8 mg/mL, which equals a final concentration of 2.5–80 μg/mL). The cumulative concentration-response curve of GT100 in endothelium-denuded aortic rings assay were constructed and compared with the endothelium-intact concentration-response curve.
Investigation of GT100's vasodilatory effects in the presence of endothelium-derived relaxing factors and its cascade inhibitors
Well-characterized endothelium-derived relaxing factors (EDRFs), nitric oxide (NO) and prostacyclin (PGI2) were studied by incubating the endothelium-intact aortic rings with endothelial NO synthase (eNOS) inhibitor, L-NAME (10 μM) or nonselective cyclooxygenase (COX) inhibitor, indomethacin (10 μM). The antagonist was incubated with the aortic rings for 20 min before they were primed with PE. Down the NO cascade, the effects of soluble guanylyl cylcase (sGC) and cyclic guanosine monophosphate (cGMP) were studied by incubating the endothelium-intact aortic rings with selective sGC inhibitor, ODQ (1 μM) or selective cGMP inhibitor, MB (10 μM), respectively, for 20 min before the rings were primed with PE. The cumulative concentration-response curves of GT100 in the presence of inhibitors were constructed and compared with a control set of experiments that had not been treated with those inhibitors.
Investigation of GT100's vasodilatory effects in the presence of G protein-coupled receptor inhibitors
The endothelium-dependent G protein-coupled muscarinic receptor (M3) and endothelium-independent G protein-coupled β-adrenergic receptor (β 2) were studied by pretreating the endothelium-intact aortic rings with nonselective M3 receptor blocker, atropine (1 μM) or nonselective β 2 receptor blocker, propranolol (1 μM), respectively, for 20 min before the addition of PE for precontraction. The cumulative concentration-response curves of GT100 in the presence of inhibitors were constructed and compared with control sets of experiments that had not been treated with these inhibitors.
Investigation of GT100's vasodilatory effects with the involvement of potassium channels
Generally, four types of potassium (K+) channels were studied in vasodilation experiments, which include the calcium-activated K+ channel (Kca), ATP-sensitive K+ channel (KATP), inward-rectifier K+ channel (Kir), and voltage-activated K+ channel (Kv). Endothelium-intact aortic rings were pretreated with nonselective Kca channel blocker, TEA (1 mM); selective KATP channel blocker, glibenclamide (10 μM); nonselective Kir channel blocker, BaCl2 (10 μM); or nonselective Kv channel blocker, 4-AP (1 mM), respectively, for 20 min before priming with PE. The cumulative concentration-response curves of GT100 in the presence of inhibitors were constructed and compared with control sets of experiments that had not been treated with those channel blockers.
Investigations of the involvement of extracellular calcium influx channels in GT100's vasodilatory effects
Commonly, the extracellular calcium influx channel is referred to as a voltage-operated calcium channel (VOCC). Before investigating the involvement of VOCC in GT100's vasodilatory effects, a preliminary set of experiments was carried out. The endothelium-intact aortic rings were primed with KCl (80 mM) to induce precontraction. Subsequently, cumulative concentration of GT100 was applied. The cumulative concentration-response curve was constructed and compared with a concentration-response curve where the precontraction was induced by PE. To determine whether the involvement of VOCC affected GT100's vasodilatory effects, three sets of experiments were performed in endothelium-intact aortic rings, which include the control, nifedipine, and GT100 groups. In the control set of experiments, aortic rings were left to stabilize in normal Krebs' solution and the integrity of the aortic rings was assessed. Subsequently, the normal Krebs' solution was replaced by calcium-free Krebs' solution that contains EGTA (0.2 mM) for 10 min to remove the Ca2+, and it was washed twice with K+-rich calcium-free Krebs' solution (91.04 mM NaCl, 50.0 mM KCl, 11.9 mM NaHCO3, 1.05 mM MgSO4, and 5.5 mM glucose, pH 7.4) at a 10 min interval and left to stabilize. After that, Ca2+ was added cumulatively at concentrations of 0.01–10 mM. The cumulative concentration-response contraction curve was constructed. This protocol was repeated with aortic rings primed with nifedipine (0.1, 0.3, and 1 μM) or GT100 (0.005, 0.02, and 0.08 mg/mL), respectively, for 20 min before the cumulative addition of Ca2+. The cumulative concentration-response contraction curves were compared with the control set of experiments. 29
Investigation of the involvement of intracellular calcium release receptor in GT100's vasodilatory effects
The calcium stored in the sacroplasmic reticulum (SR) is released into the cytosol through the inositol triphosphate receptor (IP3R) and causes vasoconstriction. To determine the involvement of IP3R, three sets of experiments were carried out, which includes control, 2-APB (positive control), and GT100 groups. In the control, endothelium-denuded aortic rings were stabilized in K+-rich Ca2+-free Krebs' solution containing 0.2 mM of EGTA for 10 min to remove the Ca2+ followed by two times of washing with K+-rich Ca2+-free Krebs' solution (without EGTA) at 10 min intervals. Subsequently, PE (1 μM) was applied and the rings were left for 20 min to induce a transient contraction. After that, the aortic rings were rinsed for 10 min with normal Krebs' solution to allow for re-absorption of Ca2+ into the SR store. In the positive control and GT100 groups, the aortic rings were incubated with 2-APB (100 μM) or GT100 (0.005, 0.02, and 0.08 mg/mL), respectively, for 20 min before the addition of PE. Then, the PE-induced contractions in the presence of 2-APB and GT100 were then compared with the control group. 29
Statistical analysis
The data were expressed as means ± standard error of the mean. Results from the controls and those pretreated with blockers were compared by one-way analysis of variance (ANOVA) and post hoc Dunnett's test by using SPSS version 20 software. The KCl-pretreated and endothelium-denuded groups were compared with the control by using t-tests. All tests were two tailed, and significance was set at P < .05.
Results
Tri-step FTIR macro-fingerprint of U. rhynchophylla and its extracts
The study used the tri-step FTIR identification method, which can gradually enhance the resolution and amplify the fingerprint feature of the samples. The 1D FTIR spectra of UR and its extracts are shown in Figure 1a, and all their characteristic peaks are assigned in Table 1. As shown in Figure 1a, the spectra of GT100, GT50, and GTW were very similar; therefore, high-resolution SD-IR and 2D-correlation IR spectroscopy were performed to further analyze and distinguish their active components. The SD-IR spectra of UR, GT100, GT50, and GTW within the range of 1800–750 cm−1 are shown in Figure 1b. The spectrum obtained for GT100 was different than those of GT50 and GTW. However, the differences between GT50 and GTW were more obscure.
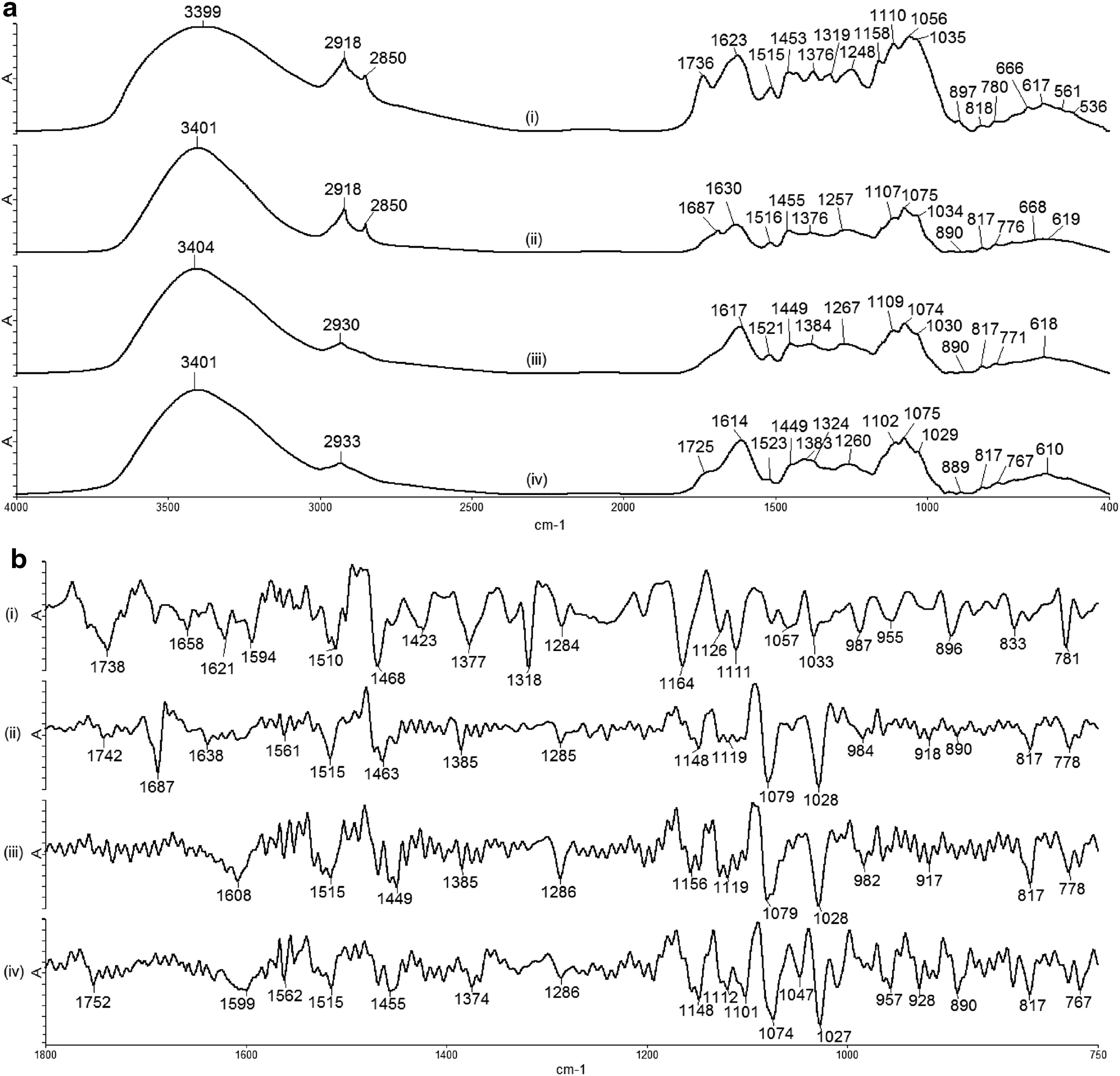
Representative
ν, stretching; ν s, symmetrical stretching; ν as, asymmetrical stretching; δ, bending.
FTIR, Fourier transform infrared.
According to Sun et al. 30 , the analysis of the 2D-correlation IR spectra of UR and its extracts were separated into two ranges such as 850–1100 cm−1 (Fig. 2) and 1200–1500 cm−1 (Fig. 3). The 2D-IR spectra of GT100 showed more obvious differences compared with the spectra of GT50 and GTW in the range of 850–1100 cm−1. This indicated the uniqueness of the macro-fingerprinting features of the tri-step FTIR identification method in identifying and distinguishing the herb and its extracts, and, thus, the spectra of UR, GT100, GT50, and GTW were used as their fingerprints.
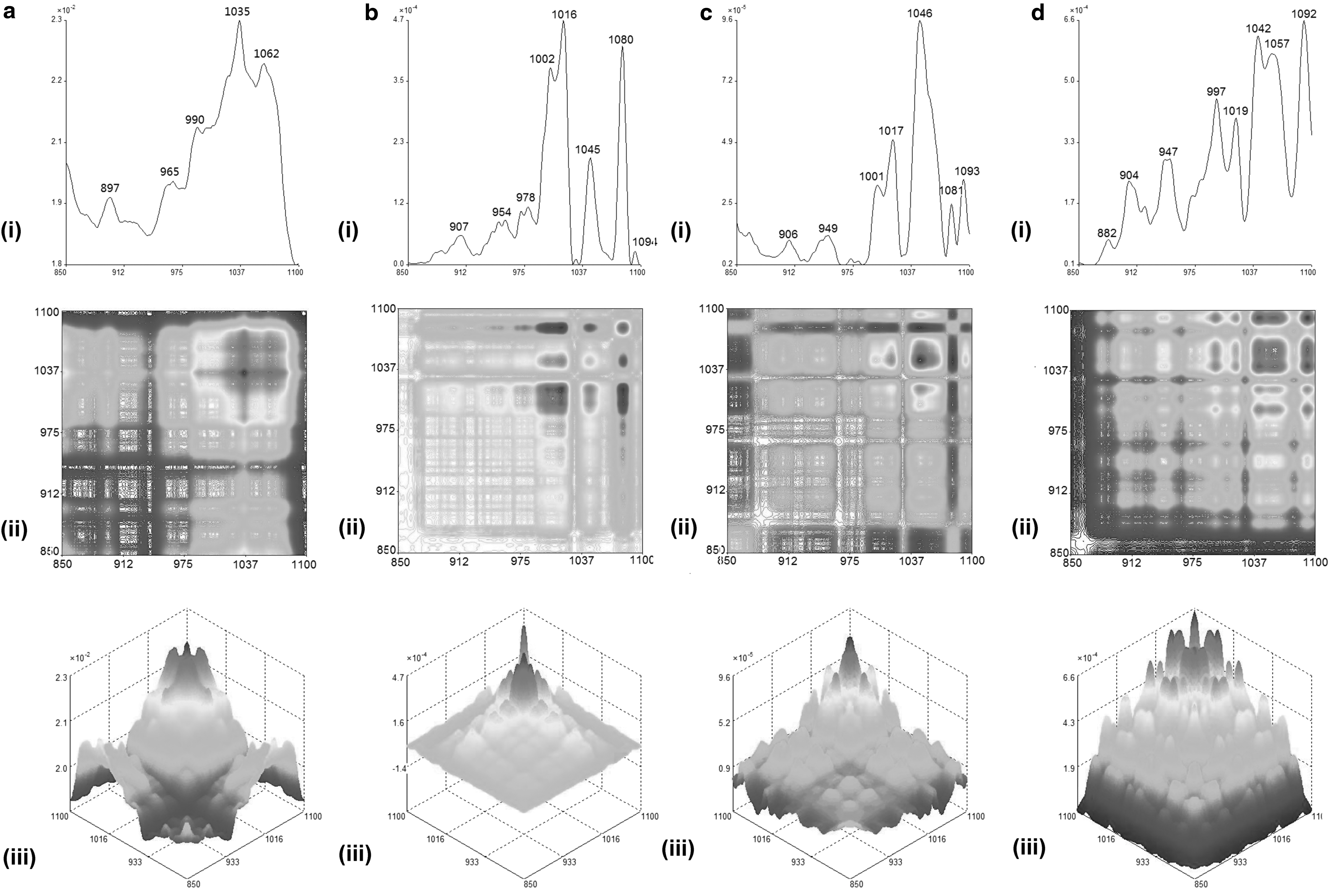
Representative (i) autopeak, (ii) synchronous 2D-IR, and their (iii) 3D-IR spectra in range of 850–1100 cm−1 for
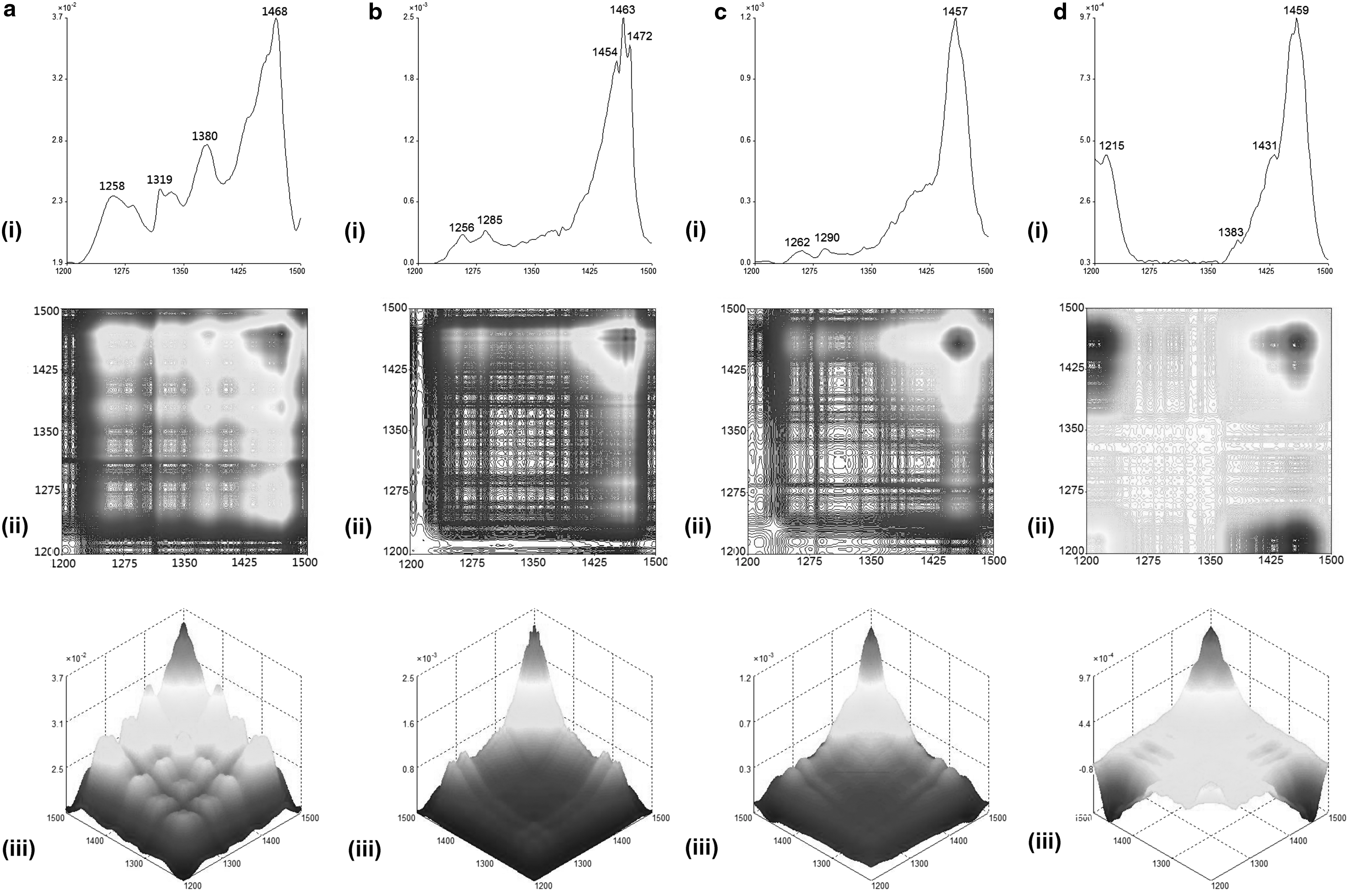
Representative (i) autopeak, (ii) synchronous 2D-IR, and their (iii) 3D-IR spectra in range of 1200–1500 cm−1 for
Effects of U. rhynchophylla extracts on PE-precontracted tone
The vasodilatory effects of three different extracts (GT100, GT50, and GTW) were tested on the isolated endothelium-intact rat aortic rings that had been primed with PE for sustained precontraction at a resting tension of 1.0 g. The primary raw data obtained from powerLab 26T are shown in Figure 4a. The GT100 exerted the highest potential of concentration-dependent vasodilatory effects compared with the other two extracts (Fig. 4b), with a maximum relaxation (R max) value of 101.3% ± 2.82% (P < .001) and half of the maximum effective concentration (EC50) at 0.028 ± 0.002 mg/mL, as shown in Table 2. Therefore, GT100 was selected for further signaling mechanism studies.
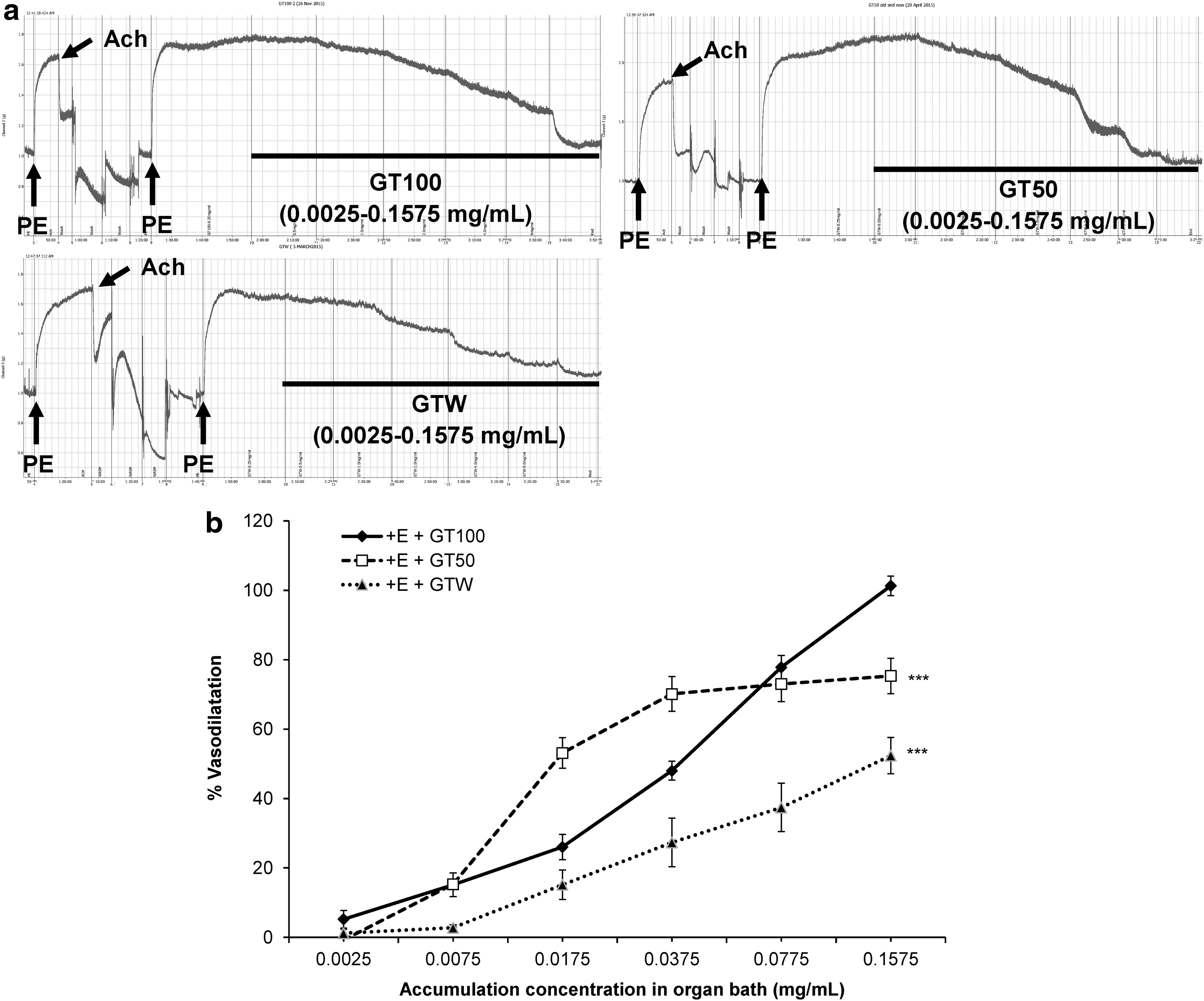
Effect of
ANOVA, analysis of variance; 4-AP, 4-aminopyridine; EC50, half of effective concentration; L-NAME, Nω-nitro-
Primary screening for endothelium-dependent and channel-linked mechanisms
Studies on the mechanism of action often start with determining the endothelium dependency as well as the channel-linked signaling mechanism pathways that mediate GT100's vasodilatory effects. The removal of the endothelium had significantly decreased the vasodilatory effects of GT100 as shown in Figure 5b, reducing about 67% of relaxation of the isolated aortic rings. Hence, the EC50 value was 2.71 ± 0.762 mg/mL whereas the R max value was 34.15% ± 2.37% (P < .001) (Table 2). Unlike its impact on endothelium-denuded rings, 80 mM of KCl precontracted aortic rings showed a similar trend of vasodilatory effects as with the control group, but to a lesser extent. The vasodilatory effect exerted by GT100 in KCl-primed aortic rings was significantly reduced compared with the control group, with an EC50 value of 0.057 ± 0.012 mg/mL and an R max value of 88.57% ± 4.64% (P < .01), as shown in Table 2. The raw data are shown in Figure 5a.
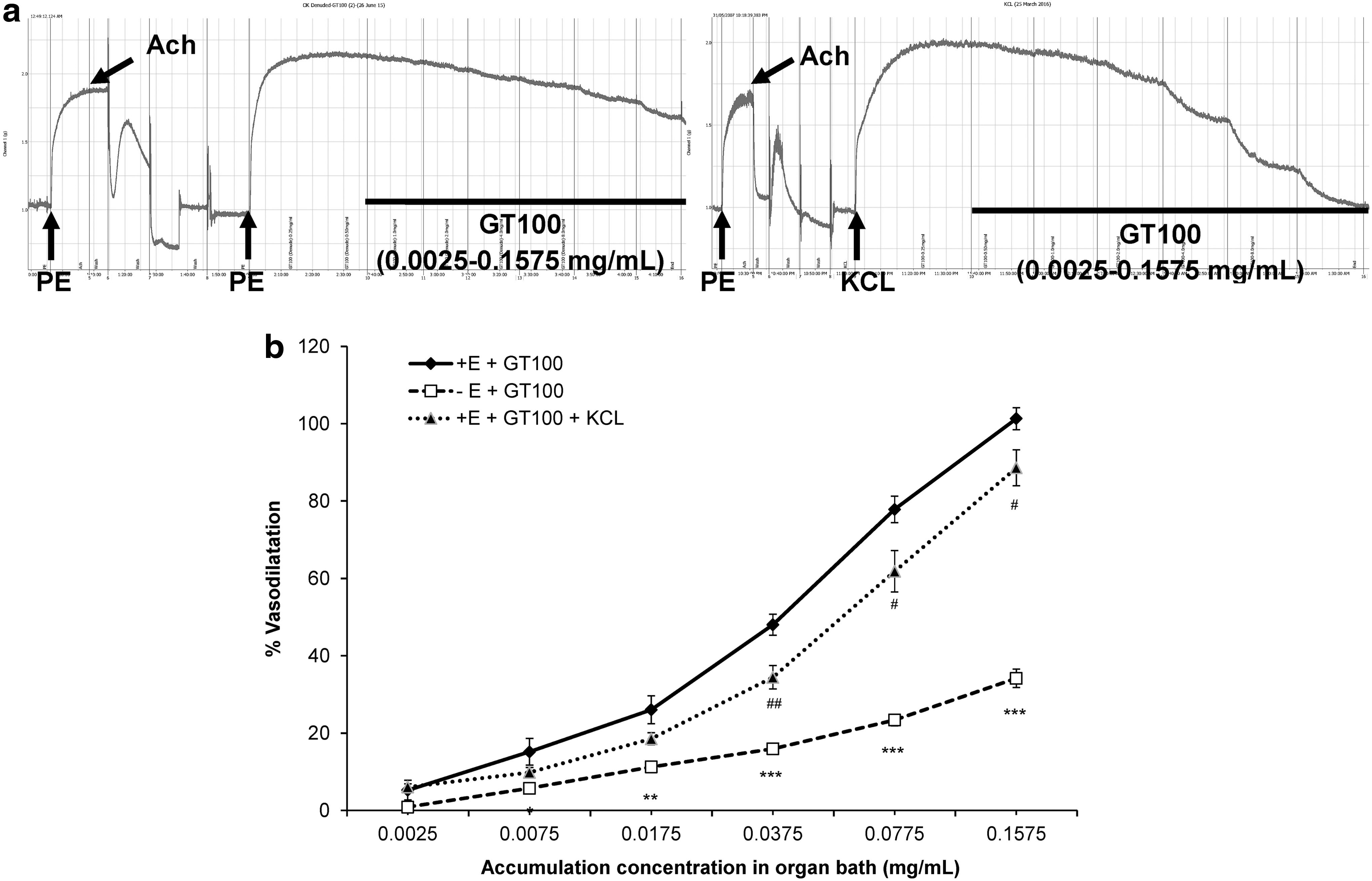
Effect of
Studies on EDRF's mechanisms
Within the endothelium, two well-known EDRFs are present, NO and PGI2. As shown in Figure 6a, the aortic rings were preincubated with antagonists, L-NAME (10 μM) and indomethacin (10 μM), respectively, in EDRF mechanism studies. Table 2 shows that the R max value was decreased in the presence of L-NAME to 49.61% ± 2.76% (P < .001) compared with the control and the EC50 value dropped to 0.345 ± 0.062 mg/mL. On the other hand, indomethacin (10 μM) also reduced the vasodilatory effects of GT100, but to a lesser extent with an R max value of 92% ± 2.47% (P < .05) compared with the control, and it increased the EC50 value to 0.038 ± 0.006 mg/mL (Table 2). To further determine the NO-linked vasodilatory cascade, ODQ (1 μM) and MB (10 μM) were applied in the test on sGC- and cGMP-dependent pathways, as shown in Figure 6a. ODQ abundantly suppressed the vasodilatory effects of GT100 to an R max value of 62.41% ± 4.24% (P < .001) compared with the control group and to an EC50 value of 0.210 ± 0.084 mg/mL. MB contributed to a blockage in the GT100's vasodilatory effects as well with an EC50 value of 0.023 ± 0.002 mg/mL and an R max value of 91.33% ± 1.99% (P < .01) compared with controls (Table 2). Figure 6b showed the extent of different inhibitory effects contributed by various antagonists.

Effects of
Studies on G protein-coupled receptor mechanisms
Other than EDRFs and enzyme-linked vasodilatory receptors, the G protein-coupled muscarinic receptors (M3) that predominate the endothelium and G protein-coupled β 2-adrenergic receptors (β 2) located on the vascular smooth muscle cells (VSMCs) could mediate the vasodilatory effects of GT100. The nonselective M3 receptor antagonist, atropine (1 μM) and the nonselective β 2-adrenoreceptor antagonist, propranolol (1 μM) were used in these studies, as shown in Figure 7a. From Table 2, it could be observed that the EC50 values in the presence of either atropine or propranolol were increased to 0.06 ± 0.017 and 0.038 ± 0.005 mg/mL, respectively, and significantly attenuated the R max values to 82.95% ± 3.97% (P < .01) and 89.19% ± 3.98% (P < .05), respectively, when compared with the control group. Figure 7b shows the different extent of inhibition in GT100's vasodilatory effects in the presence of these antagonists.
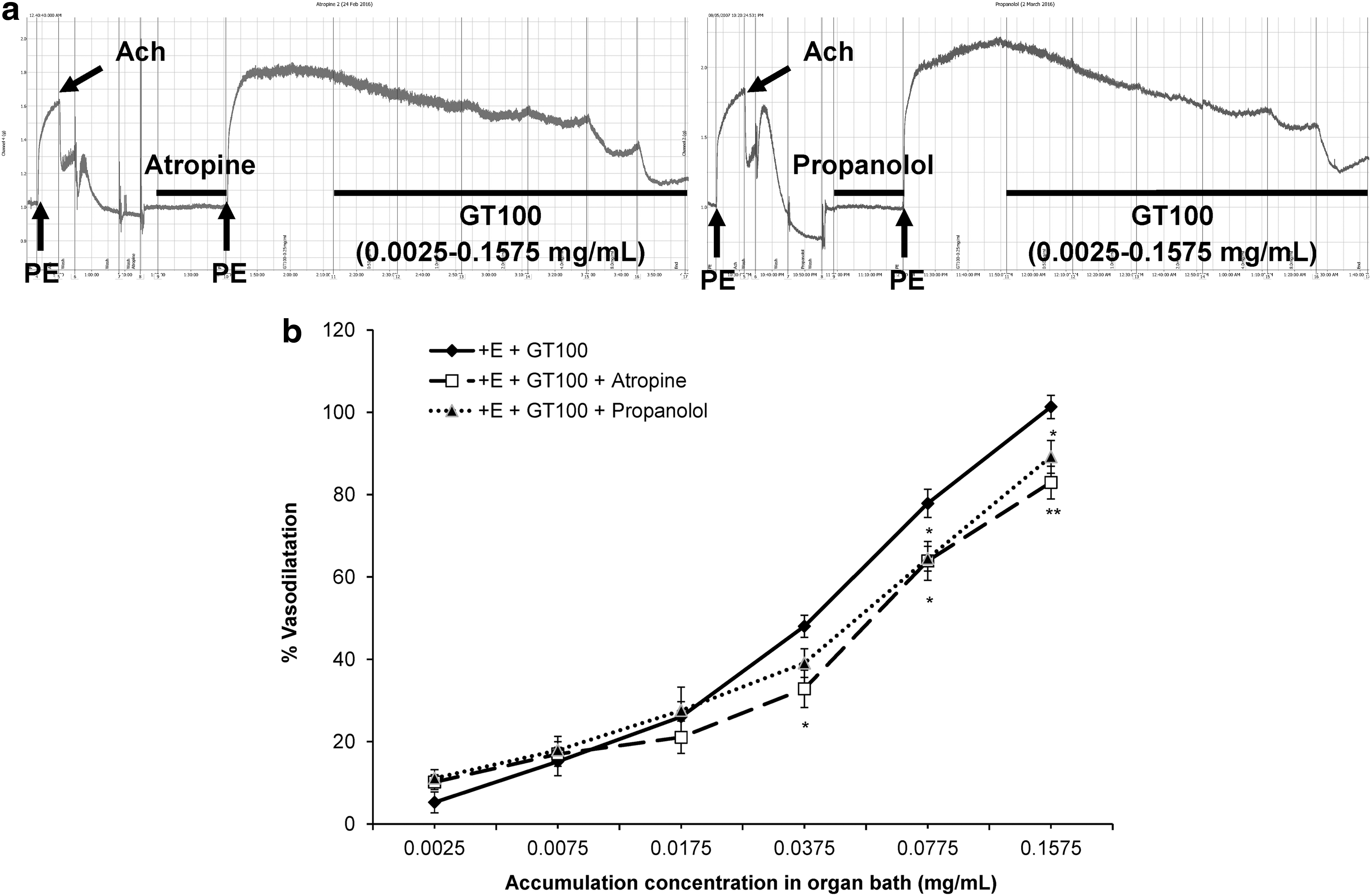
Effects of
Effects of K+ channels
There are four types of potassium channels that are frequently studied, which include Kca, KATP, Kv, and Kir. Hence, these potassium channels were assessed by using TEA (1 mM), glibenclamide (10 μM), 4-AP (1 mM), and BaCl2 (10 μM), respectively, as shown in Figure 8a. The presence of the nonselective Kca channel antagonist, TEA exerted no effect (P > .05) in vasodilation of GT100, which excluded the involvement of the Kca channel in GT100-mediated vasodilatory effects. However, the KATP channel was shown to be significantly involved in GT100-mediated vasodilatory effects since the presence of its selective blocker, glibenclamide significantly reduced the R max value to 49.67% ± 3.13% (P < .001) and increased the EC50 value to 0.446 ± 0.126 mg/mL compared with the control, as shown in Table 2. Followed by the involvement of the Kv channel, about one-quarter of the R max value of GT100's vasodilatory effects was suppressed in the presence of the nonselective Kv channel antagonist, 4-AP with a slight increase in the EC50 value to 0.092 ± 0.019 mg/mL and in the R max value of 73.04% ± 2.54% (P < .001), as shown in Table 2. Figure 8b shows that the presence of barium chloride markedly increased the vasodilatory effects of GT100 at the concentration of 0.0025–0.0375 mg/mL. However, the effect exerted by BaCl2 started to decline after the cumulative concentration of GT100 applied at 0.0775 mg/mL, indicating the competition for the site of action between the antagonist and active components of GT100. Hence, there were slight changes in the EC50 value and the R max value to 0.017 ± 0.002 mg/mL and 99.46% ± 2.16% (P < .01), respectively. The different degrees of inhibitory effects by antagonists on GT100's vasodilatory effects can be observed in Figure 8b.
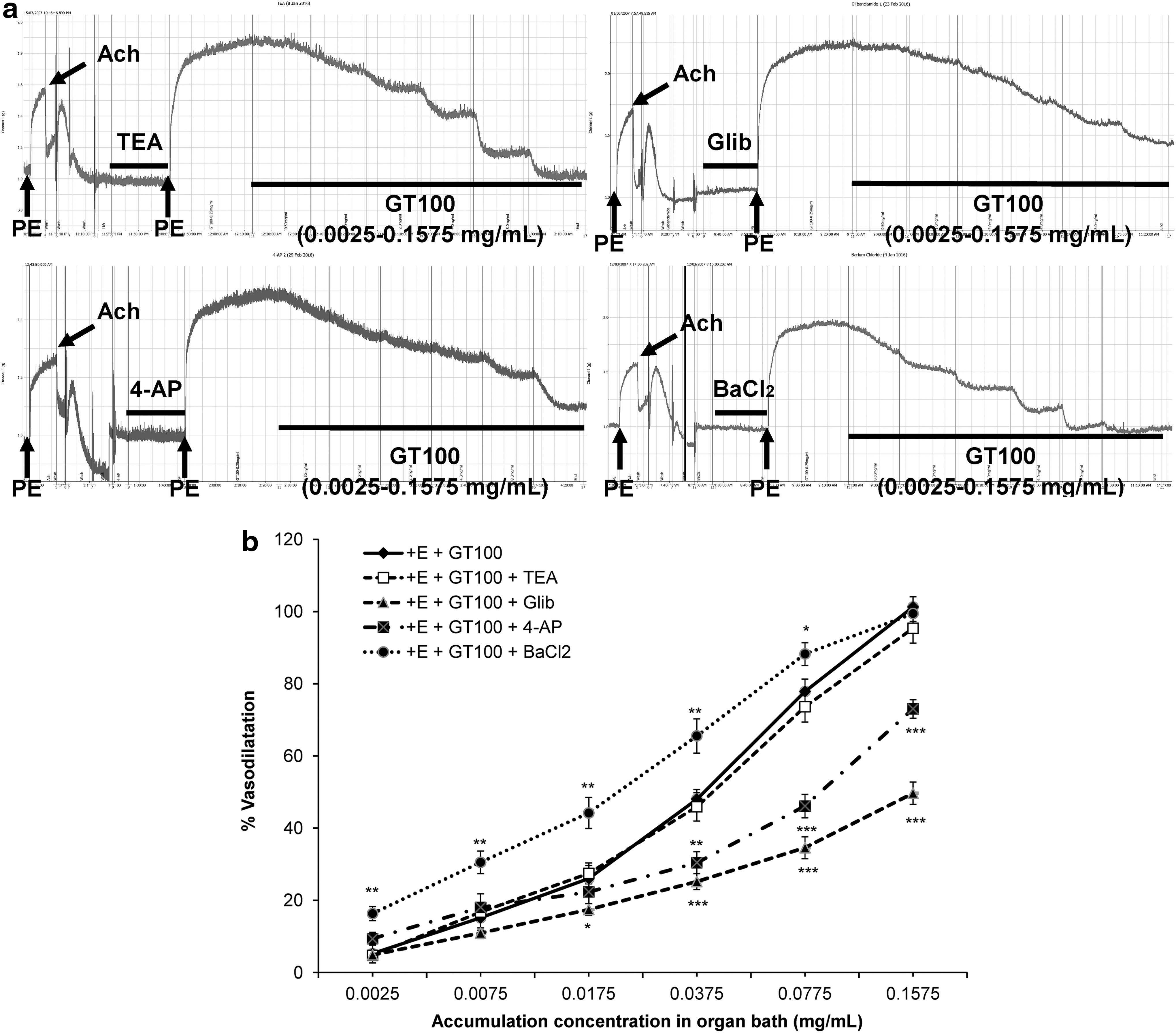
Effects of
Effect on contraction induced by extracellular Ca2+ influx
The entrance of calcium into the cytosol can occur in two ways, by extracellular influx through the VOCC and calcium efflux via IP3R 22 from SR stores. To determine whether these pathways were involved in the vasodilatory effects of GT100, three concentrations of GT100 (0.005, 0.02, 0.08 mg/mL) were applied individually in their respective tissue baths for 20 min, followed by the addition of calcium chloride (0.01–10 mM) to determine the stepwise increase of Ca2+-induced vasoconstriction. Before the commencement of the experiment, the tissue baths containing Ca2+-free Krebs' solution with endothelium-intact aortic rings were prewashed with EGTA for 10 min to eliminate any undesired residual effects that could be caused by the remaining calcium in the buffer solution. In the study of the VOCC mechanism pathway, three sets of experiments were designed for three different treatments, which are the untreated set (control), three different concentrations of selective VOCC blocker, nifedipine (0.1, 0.3, 1.0 μM) (positive control), and three different concentrations of GT100 (experimental sets). As shown in Table 3, the influx of extracellular calcium could induce up to 0.656 ± 0.04 g contraction. In contrast, the application of nifedipine had almost completely abolished the Ca2+-induced contractile effect in the isolated aortic rings and the inhibition was gradually increased with the increasing concentration of a blocker. From Table 3, the Ca2+-induced contractions in GT100 (0.005, 0.02, 0.08 mg/mL) pretreated aortic rings were 0.524 ± 0.018, 0.473 ± 0.014, and 0.351 ± 0.019 g, respectively. However, the inhibitory effects of GT100 were less when compared with the positive control, as shown in Figure 9a.
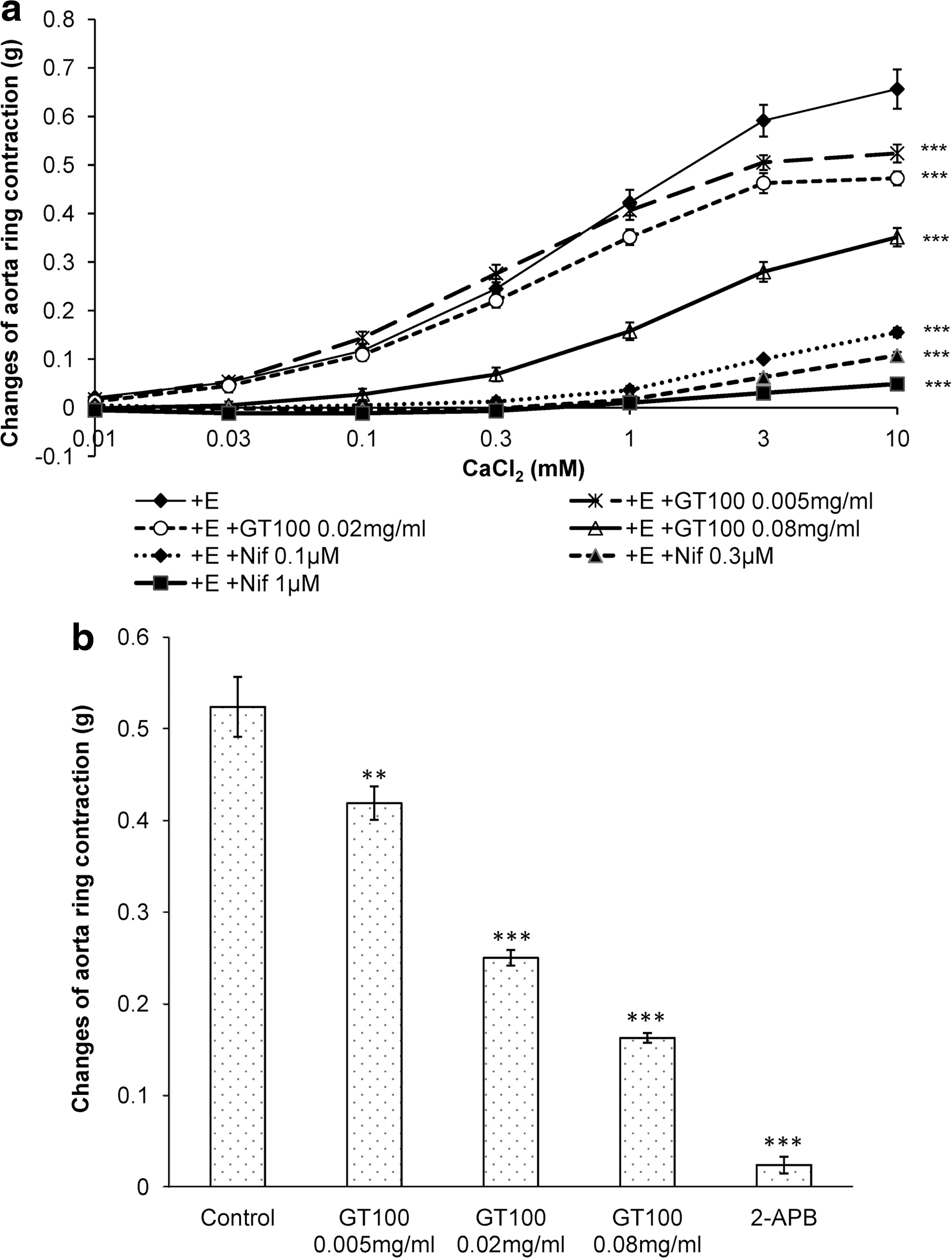
Effects of different concentrations of 95% ethanolic U. rhynchophylla extracts (GT100) (0.005, 0.02, 0.08 mg/mL) and nifedipine (0.1, 0.3, 1 μM) on CaCl2-induced contraction in endothelium-intact isolated aortic rings in Ca2+-free Krebs' solution. Figure
IP3R, inositol triphosphate receptor; n, number of determination; VOCC, voltage-operated calcium channel.
Effect on contraction induced by intracellular Ca2+ release
The same protocol as the one used to determine the VOCC mechanism pathway was utilized to study the IP3R signaling pathway. However, the application of calcium chloride was replaced with 1 μM of PE to induce the intracellular release of calcium and to compare the PE-induced contractile tone among various treatments. Three sets of experiments were carried out, which include preincubation with 2-APB (100 μM) (positive control), three different concentrations of GT100 (0.005, 0.02, 0.08 mg/mL) (experimental sets), and the untreated group (control). From Table 3, it could be observed that the selective IP3R blocker, 2-APB almost completely abolished the contractile effects induced from the intracellular release of calcium via IP3R, with maximal contraction tension of 0.0238 ± 0.009 g (P < .001). In contrast, the low concentration of GT100 (0.005 mg/mL) already started showing inhibitory effects on vasoconstriction, with tension of 0.419 ± 0.018 g (P < .01). Higher concentrations of GT100 at 0.02 and 0.08 mg/mL decreased the intracellular Ca2+ release-induced vasoconstriction with a significance of P < .001 compared with the control group. As shown in Figure 9b, the different extent of inhibition on intracellular Ca2+ release-induced vasoconstriction exerted by different concentrations of GT100 indicated the employment of this signaling mechanism pathway during its effects on vasodilation.
Discussion
U. rhynchophylla (UR), also known as Gou-teng, is one of the essential ingredients in TCM prescriptions for treating hypertension 4,5 and cardiovascular complication symptoms such as convulsions, dizziness, and numbness. 3,4,31 By using tri-step FTIR spectroscopy, the complete chemical properties of herbs were revealed. From the peak position, peak intensity, and peak shape in the UR's conventional FTIR spectrum, the components contained within the sample could be broadly inferred and compared with the atlas of UR, 30 and, thus, the authenticity of the UR used in this experiment was confirmed. In the 1D FTIR spectrum of UR [Figure 1a(i)], the characteristic peak that appeared at 1736 cm−1 represents the C = O bond of the ester group, whereas the absorption peaks at 1623 and 1319 cm−1 represent the characteristic peaks of calcium oxalate. The peaks at the sugar zone (1200–950 cm−1) formed a ladder shape, which indicated the peaks of starch. These show that UR contains ester group compounds, calcium oxalate, and starch. However, the traces of calcium oxalate and starch were not found in the spectra of GT100, GT50, and GTW, which means that neither 95% nor 50% ethanol successfully extracted these components. According to the analysis, GT100 showed a concentration-dependent vasodilatory effect with the highest percentage of R max on the aortic rings and the lowest EC50 value compared with GT50 and GTW; thus, it is used for further signaling mechanism studies. These results also proved that the vasodilatory effect of GT100 was not contributed by calcium oxalate and starch and is probably attributable to other ethanol-extractable active components. 32
The SD-IR spectra shown in Figure 1b indicated a strong absorption peak at 1687 cm−1 (C-N-H bending) in GT100, which did not appear in the spectra of GT50 and GTW. By combining the absorption peaks at 1638 cm−1 (C = O stretching), 1561, 1515, 1463 cm−1 (aromatic ring skeleton vibration), 1385 cm−1 (C-H bending), and 1285 cm−1 (C-O stretching) in the spectrum of GT100 [Fig. 1b(ii)], GT100 was confirmed to contain large amounts of alkaloids. Further, the absorption peaks of these three extracts in the sugar zone (1200–950 cm−1) are very similar, as shown in the SD-IR spectra (Fig. 1b). In Figure 2, the 2D-correlation IR proved that it has a higher resolution than SD-IR. The 2D-correlation IR spectra of these three extracts in the range of 850–1100 cm−1 are different from each other under heat perturbation of 50–120°C, which indicates different thermal stability as well as different kinds of sugar or glycoside components among them. Moreover, in Figure 3, all the extracts showed their strongest auto peaks at ∼1460 cm−1 (aromatic ring skeleton vibration). Other than this auto peak, GT100 also had two strong auto peaks at 1454 and 1472 cm−1. This indicated that GT100 contains various vasoactive components.
In the preliminary screening, endothelium-impaired aortic rings were used to investigate the vasodilatory effects of GT100. The significantly increased EC50 value suggested the endothelium-dependent vasodilatory behavior. However, the relaxation response was not completely abolished by the impaired endothelium, which suggested that GT100-mediated vasodilatory effects were mediated by endothelium-independent relaxing factors, but to a lesser extent. The results obtained were similar to those of Kuramochi et al. 33 In addition, GT100 exerted vasodilatory effects on KCl-primed aortic rings, which indicated that the depolarizing current of Ca2+ through the L-type calcium channel as well as the hyperpolarizing current through the potassium channels were involved in regulating the vascular tone.
EDRFs such as NO and PGI2 are well-known factors that dominate endothelium-dependent vasodilatory effects. In the endothelium, NO was produced from the breakdown of
In the endothelium, arachidonic acid (AA) is broken down to prostaglandin H2 by the COX enzyme, which will be catalyzed by prostacyclin synthase into PGI2, which is one of the essential EDRFs. The nonselective COX inhibitor, indomethacin, which is a nonsteroidal anti-inflammatory drug, inhibited the vasodilatory effect of GT100 on isolated aortic rings. Indomethacin is most frequently used to conduct this mechanism study because of its rapid and reversible binding on fresh application. In a time-dependent manner, it produces a higher-affinity and slow reversible binding with the COX enzyme. 37,38 However, there was a significant suppression with increasing concentrations of GT100, thus suggesting the employment of this mechanism pathway in GT100's vasodilatory effects. On comparing both EDRFs, NO was suggested as the major route employed by GT100 in exerting its vasodilatory effects, and it was only partially attributed to the production of PGI2 in an endothelium-dependent manner. The presence of the antagonists inhibited but did not block the relaxant response, which was similar to previous studies using rhynchophylline. 33,39
According to the results, the vasodilatory effects of GT100 involved endothelium-independent relaxing factors such as the β 2-adrenergic receptor. β 2-Adrenergic receptor was located in VSMC, where it was coupled with Gs α-protein. The activation of the Gs α-protein coupled to the β 2-adrenergic receptor would stimulate the production of the second messenger, cAMP through the breakdown of adenosine triphosphate (ATP), a process that is catalyzed by adenylyl cyclase (AC). The protein kinase A (PKA) would be activated by the increased production of cAMP and, thus, results in vasodilation. 35,40 The nonselective β 2-adrenergic receptor antagonist, propranolol exerted a similar amount of inhibition as indomethacin in the vasodilatory effects of GT100, both of which have an EC50 value of 0.038 mg/mL.
The nonselective M3 receptor antagonist, atropine exerted its inhibitory effect on GT100-mediated vasodilation in a similar manner. Typically, the muscarinic receptor will functionally dominate in the endothelium rather than in VSMC and is coupled with Gq α-protein. Therefore, once the M3 receptor is activated in the endothelium, the phosphatidylinositol 4,5-bisphosphate (PIP2) located on the plasma membrane would be catalyzed by phospholipase C (PLC) and cleaved into second messengers, IP3 and diacylglycerol (DAG). The IP3 will be subsequently bound to IP3R located on the SR store for the release of calcium into the cytosol, and DAG will activate the protein kinase C (PKC) and phosphorylate cytosolic proteins such as myosin light-chain kinase (MLCK). 41 The calcium released into the cytosol tends to activate eNOS by forming complexes with calmodulin, hence inducing vasodilation. Atropine significantly reduced the R max value as well as increased the EC50 values, implying its involvement in GT100 vasodilatory effects. This finding is in contrast to the previous findings due to the fact that a different species of rats had been used, and the data were analyzed by using the Aspin–Welch's t-test instead of one-way ANOVA with post hoc Dunnett's test. 33
Apart from EDRFs, G protein-coupled receptors, and enzyme-linked signaling pathways, vascular tone strictly depends on channel-linked receptors such as potassium and calcium channels to regulate the membrane potential in VSMC. 41,42 Four major types of potassium channels are evaluated in vascular tone mechanism studies: Kca, KATP, Kv, and Kir. Once these potassium channels are activated, there would be an outward movement of K+ from the cytosol across the plasma membrane, hence creating a hyperpolarizing current that results in vasodilation. However, the efflux of K+ from the cytosol will cause an imbalanced electrochemical gradient in the cell and, therefore, activate the action potential. Ca2+ would typically enter the cytosol by two routes: the extracellular influx into the cytosol via VOCC or the intracellular release of Ca2+ from the SR stores via IP3R. 23 The entrance of Ca2+ into the cytosol will depolarize the cell, and its binding with calmodulin will activate the clockwise movement of the MLC cross-bridge and result in vasoconstriction. 43 From the results obtained, it may be concluded that there was no involvement of the Kca channel due to the insignificant changes of the vasodilatory effect of GT100 in the presence of the nonselective Kca channel blocker, TEA. However, the presence of a selective KATP channel blocker exerted considerable inhibitory effects on GT100-mediated vasodilation with a similar R max value as in the presence of L-NAME. Further, 4-AP had produced at least 25% of suppression of the vasodilatory effect. Generally, the Kv channel tends to functionally correlate with the VOCC in vascular tone regulation by maintaining the membrane potential of the cell. Therefore, the results shown in Figure 9a indicated that the inhibiting effects increased gradually with preincubation of GT100 from low to high concentrations, but the effect was not as potent as the selective VOCC blocker, nifedipine. According to the results shown in the VOCC mechanism study, the depolarizing current induced by the extracellular influx of Ca2+ on the addition of calcium chloride began to decline at the concentration of 3 mM. This phenomenon could be due to the entrance of Ca2+ into the cytosol, which reached a maximal point of the depolarizing current. Hence, the VOCC started to close, allowing the efflux of cytosolic potassium and creating a hyperpolarizing current that induces vasodilatory effects. Moreover, Kir was functionally different from the other three potassium channels because it better facilitates the entrance of K+ into the cytosol to hasten the recovery of membrane potential. From the results shown, similar trends of inhibitory effects were obtained in the presence of the MB blocker. Hence, it can be concluded that the vasoactive compounds in GT100 that produce vasodilatory effects employ multiple pathways. In addition, the activation of the potassium channels within the endothelium would create an endothelium-derived hyperpolarizing factor (EDHF) that passes through the myoendothelial gap junctions, in which the EDHF would be electrically coupled with the endothelium to reach the VSMC. Since action potential only takes place in VSMC, the membrane potential was hyperpolarized and caused vasodilatory effects. 22,44 On the other hand, intracellular release of calcium into the cytosol via IP3R was markedly inhibited by GT100. Thus, the channel-linked signaling pathways employed by GT100 in eliciting vasodilatory effects include the VOCC, IP3R, and all the potassium channels except the Kca channel.
In conclusion, the 95% ethanol extract contained more vasoactive compounds from the U. rhynchophylla that would contribute to the maximum vasodilatory effects compared with other solvents. A series of signaling mechanism studies proved that GT100 has the ability to induce the production of both EDRFs such as NO and PGI2 followed by the activation of their second messengers, cGMP and cAMP. Both G protein-coupled M3 and β 2 receptors as well as the VOCC, IP3R, and all the potassium channels except the Kca channel were involved in the GT100-mediated vasodilation.
Footnotes
Acknowledgments
The author would like to express deepest appreciation to all the authors who made contributions to this article.
AUTHORS' CONTRIBUTIONS
This article and the experiments pertaining to it were conducted by Y.C. Loh; part of the article was prepared by C.S. Tan and Y.S. Ch'ng; whereas the whole experimental design and ways to prepare the article were coordinated by M.A., M.Z.A., and M.F.Y.
Author Disclosure Statement
No competing financial interests exist.