
Abstract
We previously showed that Aster spathulifolius Maxim extract (ASE) reduced body weight gain and serum and liver lipid levels and significantly suppressed serum insulin and leptin concentrations in high-fat diet (HFD)-induced obese rats. Germacrone (GM) was identified as a potent bioactive constituent of ASE. In this study, we hypothesized that GM can attenuate hyperlipidemia by alleviating fatty acid (FA) synthesis/uptake and improve lipid metabolism by stimulating FA β-oxidation in HFD-induced obese C57BL/6J mice. To induce obesity, mice were fed an HFD for 6 weeks, while control mice were fed a commercial standard diet. The mice were allocated to six groups and fed either a normal diet, HFD, HFD with GM (5, 10, and 20 mg/kg), or HFD with 200 mg/kg Garcinia cambogia extract for 30 days. In the GM groups, body weight gain, visceral fat pad weight, fasting plasma glucose, serum insulin and leptin, and serum, as well as hepatic lipid, levels were attenuated. Transcriptional factors related to lipid metabolism, such as AMP-activated protein kinase α, sterol regulatory element-binding protein (SREBP) 1, SREBP 2, acetyl-CoA carboxylase, peroxisome proliferator-activated receptor (PPAR)-α, PPAR-γ, FA synthase, and carnitine palmitoyltransferase 1, showed higher expression in the GM groups. In summary, GM may help attenuate hyperlipidemia by suppressing FA synthesis and uptake by inhibiting SREBP signaling pathway activation and improve lipid metabolism by stimulating FA β-oxidation by activating the AMPKα signaling pathway in HFD-induced obesity.
Introduction
O
Obesity is characterized by the stimulated growth of adipose tissue resulting from both hyperplasia (increase in fat cell number) and hypertrophy (increase in fat-cell size). 6 The regulation of energy homeostasis is a key point in overweight and obesity, in which adipose tissues play an important role as critical integrators, through the regulation of both energy intake and expenditure. 7
Current medications for the management of obesity, including liraglutide, phentermine–topiramate, naltrexone–bupropion, lorcaserin, and orlistat, are limited by their adverse effects, including gastrointestinal side effects, dry mouth, paresthesia, and colon ischemia. 8 –10 Even European marketing authorizations for phentermine–topiramate and lorcaserin were refused by The European Medicines Agency due to safety concerns. 9 Therefore, various extracts and compounds from natural resources are being studied to identify alternative therapeutic options for treating obesity without significant side effects. 11,12 Numerous natural products, such as crude extracts and isolated compounds from medicinal plants (e.g., apigenin, genistein, quercetin, rutin, and catechin), have shown strong antiobesity activities. 13 –16
Garcinia cambogia (GC) has been used for centuries in Asian countries for food. Currently, numerous GC/hydroxycitric acid dietary supplements for weight management are being sold around the world.
17,18
In previous in vitro studies, GC extract (GCE) inhibited lipid droplet accumulation in 3T3-L1 preadipocytes and adipogenic differentiation of preadipocytes.
19,20
In an in vivo study, GCE effectively decreased the body weight gain, visceral fat accumulation, blood and hepatic lipid concentrations, and plasma leptin and insulin levels in high-fat diet (HFD)-induced obese mice.
17
In another in vivo study, a mixture of GCE, soy peptide, and
Aster spathulifolius Maxim (AS, Asteraceae family) is an edible plant that is distributed along the coasts of southeastern South Korea. 22 The aerial parts of AS have been used in South Korea to treat asthma and diuresis. 23 Pharmacological studies have demonstrated that AS possesses antiviral and antiallergic activities. 24,25
In our previous in vitro and in vivo studies, AS extract (ASE) showed inhibitory effects on adipogenesis/lipid accumulation in 3T3-L1 preadipocytes and antiobesity effects in HFD-induced obese rats. 22,26 In the in vitro study, ASE significantly suppressed lipid accumulation/adipogenesis and inhibited early and major adipogenic factors through the AMPKα signaling pathway. In the in vivo study, ASE reduced body weight gain and lipid levels in the serum and liver and significantly suppressed serum insulin and leptin concentrations. Germacrone (GM) was identified as a potent bioactive constituent of ASE. 27 GM is known for its anticancer and antiviral activities. 28 –30 However, the antiobesity effects of GM and its molecular mechanism remain unclear. In this study, we hypothesized that GM can attenuate hyperlipidemia by alleviating fatty acid (FA) synthesis/uptake and improve lipid metabolism by stimulating FA β-oxidation in HFD-induced obese C57BL/6J mice. Specifically, we investigated the effect of GM on serum and hepatic lipid metabolism and the expression of lipid metabolism-related factors in epididymal white adipose tissue, skeletal muscle, and the liver in obese mice.
Materials and Methods
Preparation of ASE, isolation of GM, and preparation of GCE
AS leaves were collected along the coasts of Ulleung Island in Korea and were identified by a manager at the Ulleung-gun Agriculture Technology Center. The leaves were washed and dried at 25°C for a day. The dried AS leaves were ground to a fine powder. The ground sample was refluxed three times with 50% aqueous edible ethanol at 60°C for 4 h. After filtering, the extract was evaporated under vacuum using a rotary evaporator (Eyela, Tokyo, Japan) and then spray-dried. The extract powder was stored at −80°C for future use. An edible ethanol extract of AS was obtained (27.1% yield). 22 The components of the thus obtained ASE had been analyzed previously. 22
ASE was suspended in chloroform and kept at room temperature overnight. The suspension was filtered, and the chloroform-insoluble portion was washed three times with a small portion of chloroform and dried to obtain the insoluble and soluble fractions. The combined filtrate was concentrated and subjected to repeated silica gel column chromatography (packed with 200 g of silica gel) using chloroform as the eluent to obtain GM (1.8 g). 27 The structure of GM is shown in Figure 1A.

Germacrone
Mp: 51–52°C; [α]25 D-24 (c = l, methanol); EI-MS m/z: 218 (M+, C15H12O); IR (KBr) Vmax (cm−1): 1672, 1443, 1385, 1289, 1178, 1134, 858, and 812; UV (MeOH) λmax nm (logɛ): 217.5 (4.03).
1 H-NMR (CDCl3) δ: 4.97 (1H, dd, J = 10.5, 10 Hz, 2-CH), 2.35 (1H, m, 3-CH2), 2.08 (1H, m, 3-CH2), 2.10 (2H, m, 4-CH2), 4.70 (1H, ddd, J = 10.5, 3.0, LOHz, 6-CH), 2.90 (2H, m, 7-CH2), 2.95 (1H, d, 10-CH2O), 3.40 (1H, d, J = 10.5 Hz, 10-CH2O), 1.76 (3H5 s, 12-CH3), 1.71 (3H, s, 13-CH3), 1.61 (3H, t-like, 14-CH3), and 1.42 (3H, t-like, 15-CH3) (Fig. 1B).
13C NMR (400 MHz, CDCl3) δ (ppm): 132.659 (1), 24.032 (2), 38.027 (3), 126.616 (4), 125.336 (5), 29.192 (6), 129.503 (7), 207.909 (8), 55.865 (9), 134.582 (10), 137.186 (11), 22.316 (12), 19.861 (13), 15.548 (14), and 16.669 (15) 31 (Fig. 1C).
GCE (Super CitriMax®) was purchased from InterHealth Nutraceuticals (Benicia, CA, USA).
Animals and diets
Male C57BL/6J mice (5 weeks of age) were obtained from Daehan BioLink (ChungBuk, Korea) and acclimated for 1 week before the experiments. The animals were individually housed in standard cages in a room with a 12-h light/12-h dark cycle at 21°C ± 2.0°C and 50% ± 5% humidity. The mice were provided a laboratory diet and water ad libitum. All of the experimental protocols involving the use of animals were approved and conducted by the Institutional Animal Care and Use Committee of Kyung Hee University (Approval No. KHP–2014–11-2), Seoul, Korea. To induce obesity, the mice were fed an HFD (Rodent diet D12091402; Research Diets, New Brunswick, NJ, USA) consisting of 60% fat, 20% protein, and 20% carbohydrates. Control mice were fed a normal diet (ND; Rodent diet D12450B; Research Diets) consisting of 10% fat, 20% protein, and 70% carbohydrates. After 6 weeks, the mice were subdivided into six dietary groups (n = 7) (Table 1): an ND group, an HFD group, HFD+GM (5, 10, or 20 mg/kg body weight) groups, and an HFD+GCE (200 mg/kg) group, and were orally administered GM or GCE (positive control) once per day for 30 days. The ND and HFD groups were administered 0.5% carboxymethylcellulose solution. Body weight and food intake were measured every 3 days during the feeding period.
CMC, carboxymethylcellulose; GCE, Garcinia cambogia extract; GM, germacrone; HFD, high-fat diet; ND, normal diet.
Blood biochemical parameter analyses
At the end of the experimental period, food was withheld for 12 h before sacrifice. Blood samples were collected from the inferior vena cava of the euthanized mouse, allowed to clot for 30 min at room temperature, and centrifuged at 900 g for 15 min at 4°C. The epididymal, retroperitoneal (RP), and perirenal, mesenteric, and brown fat pads and the livers were removed, rinsed with saline, and weighed. The livers and fat pad samples were stored in serum at −80°C until analysis. Serum and hepatic triglyceride (TG) and total cholesterol (TC), serum high-density lipoprotein cholesterol (HDL-C), alanine aminotransferase (ALT), aspartate aminotransferase (AST), and blood urea nitrogen (BUN), as well as the fasting blood glucose, were determined using commercial kits (Asan Diagnostics, Seoul, Korea). Hepatic TG and TC were measured according to the Folch method. 32 The concentration of serum adiponectin was measured using a mouse enzyme-linked immunosorbent assay kit (R&D Systems, Minneapolis, MN, USA), and leptin and insulin levels were analyzed using enzyme-linked immunosorbent assay kits (Morinaga Institute of Biological Science, Yokohama, Japan). The absorbance was measured using a microplate spectrophotometer (BioTek, Inc., Winooski, VT, USA). All biochemical analyses were carried out according to the manufacturers' protocols.
Histological analysis
Liver tissues and epididymal fat pads were fixed in a 10% neutral formalin solution and then embedded in paraffin. All of the tissues were cut to a thickness of 4 μm and stained with hematoxylin and eosin (H&E). The liver sections were additionally stained with Oil Red O (ORO) reagents. Then, the samples were viewed with an optical microscope (Olympus Optical, Tokyo, Japan) and images were captured at a magnification of 200 × .
RNA extraction and quantitative reverse transcription
Tissue samples from the liver, epididymal adipose tissue, and skeletal muscle were homogenized, and total RNA was extracted using the R&A-BLUE reagent (iNtRON, Seongnam, Republic of Korea). The mRNA was processed for cDNA synthesis using a cDNA synthesis kit (Takara, Shiga, Japan). After cDNA synthesis, quantitative reverse transcription–polymerase chain reaction (RT-qPCR) was carried out in 2 μL of SYBR premix EX Taq (Takara) using an ABI StepOnePlus™ RT-qPCR machine (Applied Biosystems, MA, USA). Gene expression was normalized to β-actin. Gene-specific primers for the mouse genes analyzed in this study are shown in Table 2.
RT-qPCR, quantitative reverse transcription–polymerase chain reaction.
Western blotting
Western blotting experiments were conducted according to formerly published methods. 33 Liver tissues (150 mg) were homogenized with an ice extraction buffer (20 mM Tris-HCl [pH 7.4], 0.32 mM sucrose, protease inhibitor, 1 mM PMSF, 0.5 M EDTA [pH 8.0], 1 mM NaF, and 1 mM Na3VO4) and centrifuged at 13,000 rpm for 15 min at 4°C to recover the supernatants. The protein concentrations in the whole cell lysates were measured using a Pierce™ BCA protein assay kit (Thermo Fisher Scientific, Rockford, IL, USA). Equal amounts of protein (30 μg) were subjected to electrophoresis on a sodium dodecyl sulfate–polyacrylamide gel (7.5%) and transferred onto a polyvinylidene fluoride membrane. The membranes were blocked in blocking solution (5% nonfat dried milk in Tris-buffered saline containing 0.1% Tween-20). Primary antibodies were used at a dilution of 1:1000 and incubated at 4°C overnight. Thereafter, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies used at 1:5000 dilutions. Antibody binding was visualized using an LAS3000® luminescent image analyzer (Fuji Film, Tokyo, Japan). Immunoblotting was performed using the following antibodies: anti-PPAR-α, anti-SREBP-1, anti-SREBP-2, and anti-β-actin from Santa Cruz Biotechnology (Santa Cruz, TX, USA); anti-FAS, anti-CPT1, anti-p-ACC (Ser79), anti-ACC, and anti-AMPKα from Cell Signaling Technology (Danvers, MA, USA); and anti-p-AMPKα (Thr172) from Millipore (Billerica, MA, USA).
Statistical analyses
All of the data are expressed as the mean ± SE, and all of the experiments were carried out in triplicate. The data were analyzed by one-way ANOVA and Tukey's honestly significant difference test. All analyses were conducted using SPSS software (Version 21.0; SPSS, Inc., Chicago, IL, USA).
Results
Effects of GM on body weight, food intake, and adipose tissue weight
After being fed an HFD for 6 weeks, the final body weights, body weight gains, and serum lipid parameters of the HFD group were significantly greater than those of the ND group (Table 3). GM supplementation suppressed the HFD-induced body weight increase in a dose-dependent manner. In contrast, GCE supplementation did not significantly reduce the body weight gain induced by HFD. During the treatment period, food intake was markedly reduced in HFD-fed groups compared with ND group, this may be due to the HFD (diet with 60% kcal% fat) containing six times as much fat as the ND (diet with 10% kcal% fat). However, food intake did not significantly differ among the HFD-fed groups. Additionally, the epididymal, RP, and perirenal, mesenteric fat pad weights were significantly higher in the HFD group than in the ND group, and these increases were lowered by GM treatment (Table 3). These results were consistent with those of histological analysis of the mouse epididymal adipose tissues. The stained adipocytes in the epididymal white adipose tissues were visibly larger in the HFD group than in the GM- or GCE-treated groups (Fig. 2A).
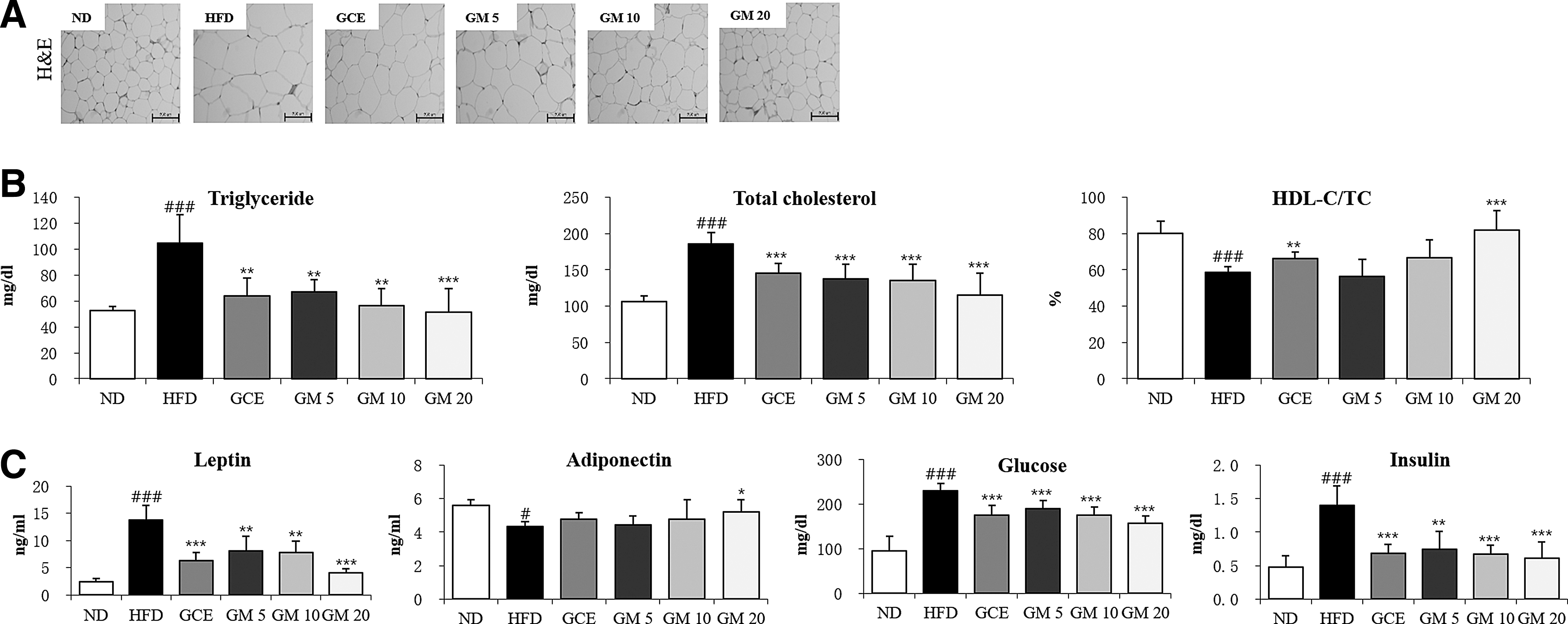
The data are presented as the means ± SEs (n = 5–7). # P < .05, ## P < .01, and ### P < .001 versus the ND group. * P < .05, ** P < .01, and *** P < .001 versus the HFD group.
Effects of GM on serum lipid parameters
The HFD group showed significantly higher TG and TC levels and lower HDL-C/TC ratios than the ND group (Fig. 2B). However, the GM- and GCE-treated groups exhibited reversals in these parameters to levels similar to those observed in the ND group. These results suggested that accumulation of body fat was inhibited by GM treatment, which resulted in improved serum lipid profiles.
Additionally, mice in the HFD group showed increased serum leptin, glucose, and insulin levels (Fig. 2C). Conversely, compared with the ND group, the HFD group showed significantly lower adiponectin levels. The GM- and GCE-treated groups showed lower serum leptin, glucose, and insulin levels and increased adiponectin levels.
To evaluate the potential toxic effects of GM on hepatic and renal functions, we measured the serum ALT, AST, and BUN levels in GM-treated obese mice (Table 4). The ALT, AST, and BUN levels were not significantly different between the GM and GCE groups compared with the HFD group. The ALT, AST, and BUN levels in all of the treatment groups fell in the normal ranges according to previously published reports. 34 These results indicated that GM supplementation for 30 days does not affect liver and kidney functions.
The data are presented as the means ± SEs (n = 5–7).
ALT, alanine aminotransferase; AST, aspartate aminotransferase; BUN, blood urea nitrogen.
Effects of GM on the expression of lipid metabolism-related factors in epididymal white adipose tissue
Histological analysis showed that the GM-treated groups had smaller adipocytes than the HFD group. To understand the action mechanism of GM, the mRNA expression levels of the lipogenic transcription factor (PPAR-γ and SREBP-1c) and lipogenesis-related genes (ACC1, FAS, and SCD1) were measured in epididymal white adipose tissue by RT-qPCR. The expression levels of PPAR-γ, SREBP-1c, ACC1, FAS, and SCD1 in the HFD group were significantly higher than those in the ND group (Fig. 3A). However, GM inhibited the increase in mRNA expression in a dose-dependent manner.

Effects of GM on the expression of lipid metabolism-related factors in skeletal muscle
To understand the effects of GM on lipid metabolism, the expression of lipid-regulating genes related to FA oxidation and lipolysis in skeletal muscle was investigated. The mRNA expression levels of both FA oxidation-related genes (ACC2 and CPT1b), ligand-induced transcription factor PPAR-α, and its downstream target genes (ACOX1, UCP2, and UCP3) were measured in skeletal muscle by RT-qPCR. Compared with the HFD group, GM decreased the expression levels of ACC2 and increased the expression levels of PPAR-α, CPT1b, ACOX1, UCP2, and UCP3 in a dose-dependent manner (Fig. 3B). GCE treatment showed an effect similar to that of GM treatment (5 or 10 mg/kg). These results suggested that the antiobesity effect of GM is likely due to increased FA oxidation and lipolysis in skeletal muscle through PPAR-α and its downstream target genes.
Effects of GM on hepatic lipid profiles
Hepatic TG and TC levels were significantly higher in the HFD group than in the ND group (Fig. 4C). In the GM-treated group (20 mg/kg), hepatic TG and TC levels were 31.7% and 26.1% lower, respectively, than those in the HFD group. The results from H&E and ORO staining of the liver tissues (Fig. 4A, B) were consistent with the above results. These results suggested that GM effectively inhibits lipid accumulation in the liver upon HFD.
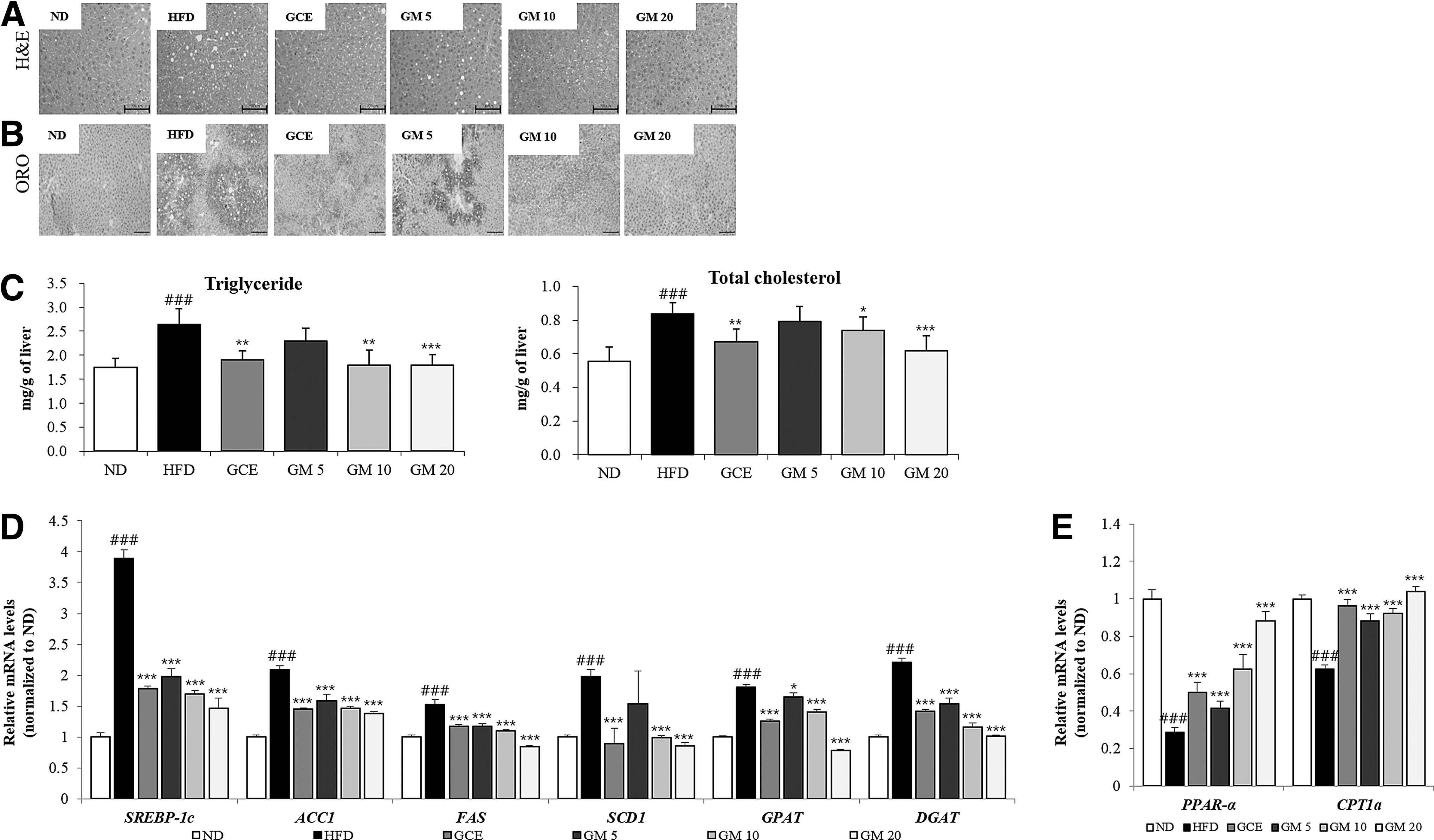
Effect of GM on lipid accumulation in liver tissue.
Effects of GM on the expression of lipid metabolism-related factors in the liver
To understand the mechanism by which GM regulates liver lipid metabolism, the mRNA expression levels of both lipogenesis-related genes (SREB-1c, ACC1, FAS, SCD1, GPAT1, and DGAT1) and FA oxidation-related genes (PPAR-α and CPT1a) in the liver were measured by RT-qPCR (Fig. 4D, E). The expression of lipogenesis-related genes in the HFD group was significantly higher than that in the ND group. However, GM supplementation suppressed this increase in mRNA expression in a dose-dependent manner. Conversely, the reduced expression of FA oxidation-related genes in the HFD group was significantly restored by GM treatment. These results suggest that the antiobesity effect of GM is likely due to an increase in FA oxidation and a decrease in lipogenesis in the liver through PPAR-α and its downstream target genes.
Effects of GM on AMPKα, ACC, PPAR-α, CPT1, and FAS protein expression and the inhibition of SREBP signaling pathway in hepatic tissue
To investigate the molecular mechanisms further, we examined the effect of GM on the AMPKα signaling pathway, which plays a key role in lipid metabolism, 35 and on the sterol regulatory element-binding protein (SREBP) signaling pathway and its downstream target proteins by using western blotting. HFD induced elevated levels of hepatic mature SREBP-1, mature SREBP-2, and FAS. However, GM treatment restituted the levels of all three proteins to near-normal levels (Fig. 5A, B).
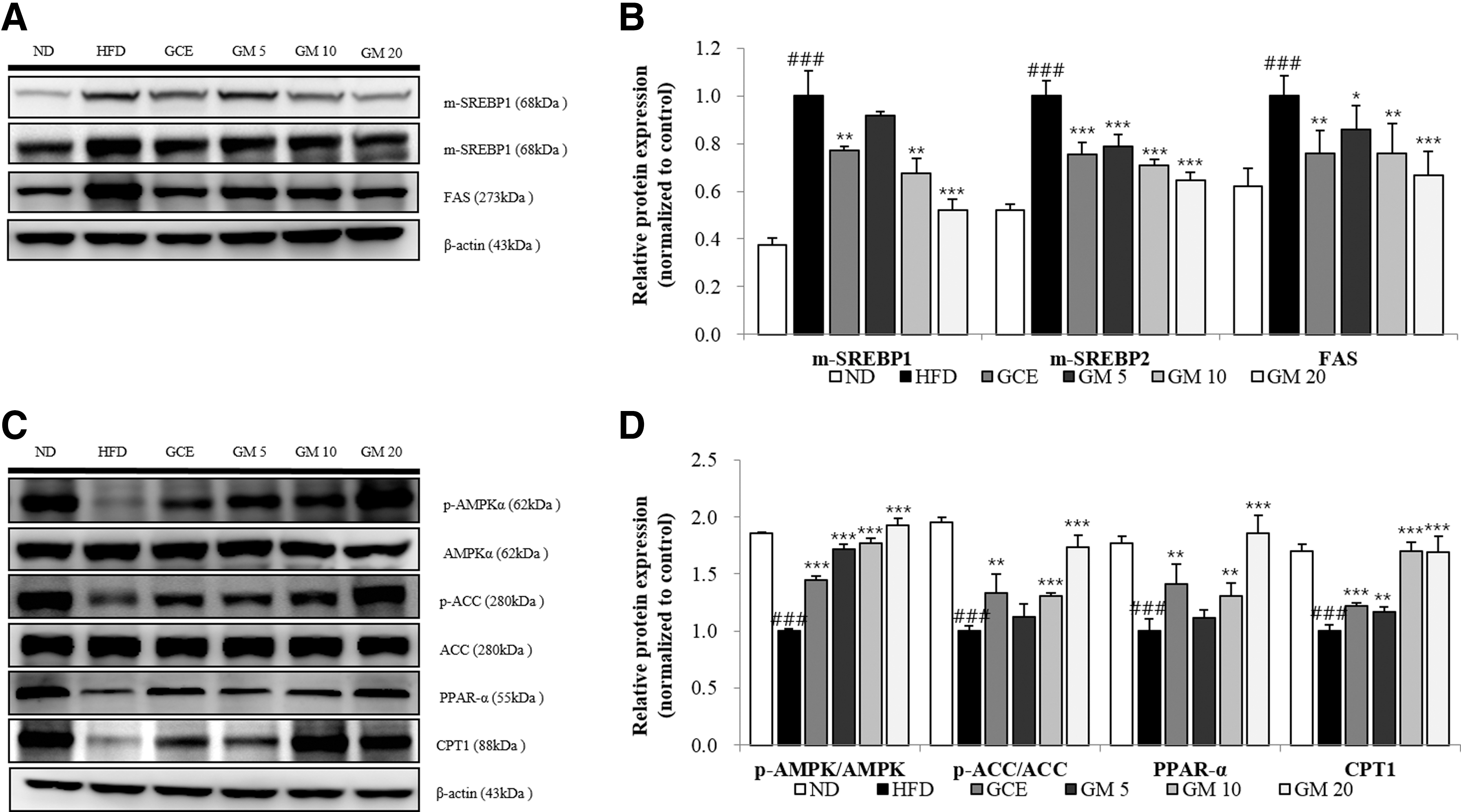
Effects of GM on m-SREBP1, m-SREBP2, FAS, CPT1, and PPAR-α protein expression levels and ACC and AMPKα phosphorylation levels in hepatic tissue.
Moreover, it increased phosphorylated AMPKα, phosphorylated ACC, PPAR-α, and CPT1 protein levels in a dose-dependent manner (Fig. 5C, D). These results suggested that GM ameliorates dyslipidemia by inhibiting SREBP signaling pathway activation and stimulating FA β-oxidation by activating the AMPKα signaling pathway.
Discussion
The key finding of our study is that GM decreased the adipocyte size by reducing lipid accumulation in the white adipose tissue, which resulted in decreased adipose tissue weight and body weight in HFD-induced obese C57BL/6J mice.
Treatment with GM or GCE ameliorated the elevated serum and liver levels of TG and TC and the lowered serum HDL-C/TC ratio induced by HFD, without any renal or hepatic toxicity.
White adipocytes secrete adipocytokines such as TNF-α, interleukin-6, leptin, and adiponectin. 36 In rodents and obese humans, plasma leptin concentrations are highly correlated with adiposity and body–mass index. 37 Circulating adiponectin is inversely correlated with body mass and profoundly influences insulin sensitivity and FA oxidation by stimulating AMP kinase activity in obese diabetic mice. 7,38 The present study showed that serum leptin levels were lower in the GM-treated group than in the HFD group. Moreover, serum adiponectin levels were higher only in the high-dosage GM-treated group when compared with the HFD group. These results suggest that GM decreases leptin levels in the serum after decreasing lipid accumulation in the adipose tissue and weight loss. Furthermore, several studies have indicated that HFD increases FA synthesis and the delivery of free FAs in the liver and decreases free FA β-oxidation, leading to accumulation of fat in the liver. 39,40 As shown in Figure 4A, B, the HFD group showed macrovesicular hepatic steatosis. However, GM remarkably ameliorated the extent of steatosis, suggesting that GM may regulate lipid mobilization and storage in adipocytes through the adjustment of leptin levels.
Both leptin and adiponectin stimulate FA oxidation by activating AMPK and ACC. 41,42 AMPK plays a key role as an energy sensor at the cellular and whole-organism levels. 43 Once activated, AMPK phosphorylates and inhibits ACC and diminishes the level of malonyl-CoA, which is an intermediate in de novo FA synthesis, and inhibits CPT1, an enzyme that regulates mitochondrial FA oxidation. 44,45 In the present study, the protein levels of phospho-AMPK and phospho-ACC, which were decreased in the livers of mice fed the HFD, were upregulated when they were given GM. Therefore, it appears that GM stimulates FA β-oxidation by activating the AMPK pathway.
In contrast, PPAR-α has been shown to regulate lipid metabolism by regulating the transcription of target genes as a ligand-induced transcription factor. 46 Increasing the expression of PPAR-α improved the expression of CPT1, ACOX1, UCP2, and UCP3, genes known to elevate energy expenditure, eventually leading to antiobesity effects. 47,48 The expression of PPAR-α in the skeletal muscle was significantly increased in the GM-treated group compared with the HFD group. GM supplementation significantly increased the expression of PPAR-α downstream target genes, including ACOX1, CPT1b, UCP2, and UCP3. Additionally, GM inhibited the expression of ACC2, a potent inhibitor of FA oxidation, in the skeletal muscle. Downregulation of ACC2 decreases malonyl-CoA content in the cell, thus increasing CPT1 expression and enhancing FA transport into the mitochondria for β-oxidation. 49 Our data showed that GM might reduce body fat by activating FA oxidation in the skeletal muscle through inhibition of ACC2. These results suggest that the antiobesity effect of GM is likely due to an increase in FA oxidation and thermogenesis in the liver and skeletal muscle through PPAR-α and its downstream target genes.
SREBP-1c is an important transcription factor for FA uptake and TG synthesis. 50 It transactivates downstream genes encoding lipogenic enzymes, including FAS, ACC, and SCD1. 51 Various genes (e.g., GPAT and DGAT) are important for catalyzing several steps in TG synthesis. 52 Our results showed that GM decreased the SREBP-1c, FAS, ACC1, SCD1, GPAT1, and DGAT1 mRNA levels in the liver. Mature SREBPs (68 kDa) were also lower upon GM supplementation in the HFD in the liver. These results suggest that GM inhibits the SREBP signaling pathway that is activated by the HFD.
PPAR-γ is a transcription factor that is mainly expressed in adipose tissue and that plays an important role in adipogenesis, lipid accumulation, and glucose homeostasis. 50,53 PPAR-γ regulates the transcription of target genes, including FAS, aP2, and lipoprotein lipase (LPL). 54 FAS catalyzes the biosynthesis of long-chain FAs from acetyl-CoA precursors as a key lipogenic enzyme. 55 SREBP-1c activates FAS and PPAR-γ expression. 56 In this study, GM significantly decreased the expression of genes involved in adipose tissue lipogenesis, such as PPAR-γ, SREBP-1c, and FAS, in white adipose tissue. These results suggest that its antiobesity effects are likely also due to the inhibition of FA and TG synthesis-related gene expression.
In conclusion, GM administration has beneficial effects, including a decrease in body weight gain, adipose tissue weight, adipocyte size, and lipid levels in the serum and the liver of HFD-induced obese mice. These results suggest that GM exerts its antiobesity effects in HFD-induced obese mice by regulating lipid metabolism, by inhibiting lipogenesis, and by stimulating FA oxidation through activation of the AMPK pathway. In this perspective, GM may be a potent bioactive constituent of ASE and an effective agent for treating obesity and obesity-related comorbidities.
Footnotes
Acknowledgments
This study was supported by a grant from the Kyung Hee University in 2005 (KHU-20050434) and through the Industrial Strategic Technology Development Program (10045257, Development of functional food product for improving metabolic syndrome using natural resources and extending global market) funded by the Ministry of Trade, Industry and Energy (MOTIE, Korea).
Author Disclosure Statement
No competing financial interests exist.