
Abstract
Curcumin, a bioactive compound in Curcuma longa, exhibits various pharmacological activities, including antimalarial effects. In silico docking simulation studies suggest that curcumin possesses glycogen synthase kinase-3β (GSK3β)-inhibitory properties. The involvement of GSK3 in the antimalarial effects in vivo is yet to be demonstrated. In this study, we aimed to evaluate whether the antimalarial effects of curcumin involve phosphorylation of host GSK3β. Intraperitoneal administration of curcumin into Plasmodium berghei NK65-infected mice resulted in dose-dependent chemosuppression of parasitemia development. At the highest dose tested (30 mg/kg body weight), both therapeutic and prophylactic administrations of curcumin resulted in suppression exceeding 50% and improved median survival time of infected mice compared to control. Western analysis revealed a 5.5-fold (therapeutic group) and 1.8-fold (prophylactic group) increase in phosphorylation of Ser 9 GSK3β and 1.6-fold (therapeutic group) and 1.7-fold (prophylactic group) increase in Ser 473 Akt in liver of curcumin-treated infected animals. Following P. berghei infection, levels of pro- and anti-inflammatory cytokines, tumor necrosis factor (TNF)-α, interferon (IFN)-γ, interleukin (IL)-10, and IL-4 were elevated by 7.5-, 35.0-, 33.0-, and 2.2-fold, respectively. Curcumin treatment (therapeutic) caused a significant decrease (by 6.0- and 2.0-fold, respectively) in serum TNF-α and IFN-γ level, while IL-10 and IL-4 were elevated (by 1.4- and 1.8-fold). Findings from the present study demonstrate for the first time that the antimalarial action of curcumin involved inhibition of GSK3β.
Introduction
R
Curcumin, a major bioactive compound, present in Curcuma longa exhibits a wide array of bioactivities, including anti-inflammatory effects in many diseases, including malaria. 6,7 It has been suggested that curcumin may be developed as adjunctive therapy for cerebral malaria (CM). 8,9 In addition, combination treatment using curcumin and CQ or piperine has been reported to lower parasitemia development and improve Plasmodium berghei-infected animal survivability. 10
It is well-known that the anti-inflammatory activity of curcumin is mediated through inhibition of nuclear factor-kappa B (NF-κB) activation. 11 NF-κB is a transcription factor that is a downstream target of several cell signaling systems, including that mediated by glycogen synthase kinase-3β (GSK3β), a serine/threonine kinase, initially identified by Embi et al. for its role in the regulation of glycogen metabolism. 12 The kinase is now known to be involved in the regulation of many crucial cellular processes, including modulation of cytokine levels during inflammation. 13 –15 As an upstream signaling component, active GSK3β is able to phosphorylate directly (and activate) NF-κB p65. 16 Inhibition of GSK3 was shown to affect production of inflammatory cytokines by inhibition of the transcriptional activity of NF-κB. 15
GSK3β is one of the most studied drug targets for the development of therapeutics. 14 Inhibitors of GSK3β have been extensively evaluated for the treatment of diseases such as cancer, Alzheimer's disease, and other inflammation-related conditions. 13,14 GSK3 is also one of the protein kinases investigated as drug targets for malaria. 17 –19 In our laboratory, we have previously shown that lithium chloride (LiCl), a GSK3 inhibitor, suppressed parasitemia development in a murine model of malarial infection. 20 In another study, Dai et al. reported that LiCl treatment restored neurocognitive function in murine experimental CM (ECM). 21 In silico docking simulation studies suggest that curcumin possessed GSK3β-inhibitory properties. 22
The present study is aimed at evaluating whether the underlying mechanism of the antimalarial activity of curcumin involved inhibition of host GSK3β.
Materials and Methods
Experimental animals
Male Institute of Cancer Research (ICR) strain of mice (25 ± 5 g; 6–8 weeks old) obtained from the Animal House Complex, Universiti Kebangsaan Malaysia (UKM) were housed at the Malaria Infection Laboratory. Permission and approval for animal studies were obtained from the Universiti Kebangsaan Malaysia Animal Ethics Committee (UKMAEC) (reference number: FST/2014/HASIDAH/20-MAR./581-MAR.-2014-NOV.-2015).
Therapeutic and prophylactic tests
CQ-sensitive P. berghei NK65 strain originally purchased from MR4, United States was maintained in ICR mice. Both tests were conducted based on the method described by Peters.
23
Starting at 3 h postinfection (day 0), the experimental animals were injected (ip) for four consecutive days with either the antimalarial reference drug (10 mg/kg body weight [bw] CQ) or the GSK3 inhibitor reference (100 mg/kg bw LiCl), 0.85% saline solution (control group A), or different dosages of curcumin (3, 10, or 30 mg/kg bw) (test group B) in 0.2 mL volumes, respectively. For prophylactic evaluation, similar dosages of curcumin, reference drugs, and controls were given for 4 days consecutively before infection (last treatment at 3 h before infection). Thin smears were prepared from tail blood of each animal to determine the percentage of parasitemia on day-4 postinfection. The average percentage of chemosuppression (PC) was calculated by comparing the percentage of blood parasitemia levels in control group A and test group B:
Liver protein extraction
Protein extraction from liver was carried out as described by Lee. 24 Organs obtained from in vivo antimalarial experiments described above were each homogenized in 1 g/mL extraction buffer containing protease inhibitors (Sigma, USA) and phosphatase inhibitors (Sigma). Protein content of liver samples was determined 25 using bovine serum albumin (BSA) as a standard.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis and western analysis
Equal amounts of 40 μg (liver) protein samples were loaded into each well of sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels (30% w/v acrylamide). Separated proteins were transferred onto nitrocellulose membranes then blocked using 3% BSA in Tris-buffered saline-Tween 20 (TBST) (0.1% w/v Tween-20 in TBS). 26 Membranes were further probed and incubated overnight at 4°C with primary monoclonal antibodies, anti-GSK3β, anti-phosphoSer9-GSK3β, anti-Akt, anti-phosphoSer473-Akt, anti-NF-κB, or anti-phosphoSer536-NF-κB (Cell Signaling, USA), before a 2-h incubation with corresponding secondary antibody, anti-rabbit IgG, HRP-linked antibody (Cell Signaling) at room temperature. Membranes were stripped with 0.2 M NaOH and then reprobed with anti-β-actin (Santa Cruz Biotechnology, USA) to ensure equal protein loading. Immunoreactive bands were detected using SuperSignal West Pico Chemiluminescent Substrate Kit (Thermo Scientific, USA). Band area intensity was quantified using a densitometer (Vilbert Lourmat 302, France).
Cytokine analysis
At day-4 postinfection during therapeutic and prophylactic treatments, blood was collected from euthanized mice (n = 6) and processed. 27 The levels of pro- and anti-inflammatory cytokines (tumor necrosis factor [TNF]-α, interferon [IFN]-γ, interleukin [IL]-10, and IL-4) were determined by an enzyme-linked immunosorbent assay kit (Qiagen, Germany).
Statistical analysis
Statistical significance of data between groups was evaluated using analysis of variance and log rank test (for Kaplan–Meier survival analysis). Data obtained are expressed as mean ± SD. A P value of <.05 between groups was considered to be significant.
Results
Curcumin inhibited parasitemia development in P. berghei-infected mice during both therapeutic and prophylactic antimalarial tests
Mice administered (ip) with curcumin for four consecutive days following injections with P. berghei-infected erythrocytes showed dosage-dependent suppression of parasitemia development (Table 1). At the highest dosage tested (30 mg/kg bw), curcumin inhibited P. berghei parasitemia development in mice by 67.6% ± 1.7%. Survivability of infected animals treated with the compound (30 mg/kg bw) was prolonged compared to nontreated mice (negative control), which did not survive beyond day 25 after infection. Median survival time for nontreated infected mice was 19 days. Control experimental animals administered CQ (10 mg/kg bw) showed nearly 100% suppression of parasitemia development on day-4 and survived throughout the observation period of 30 days (Table 1) (Fig. 1).
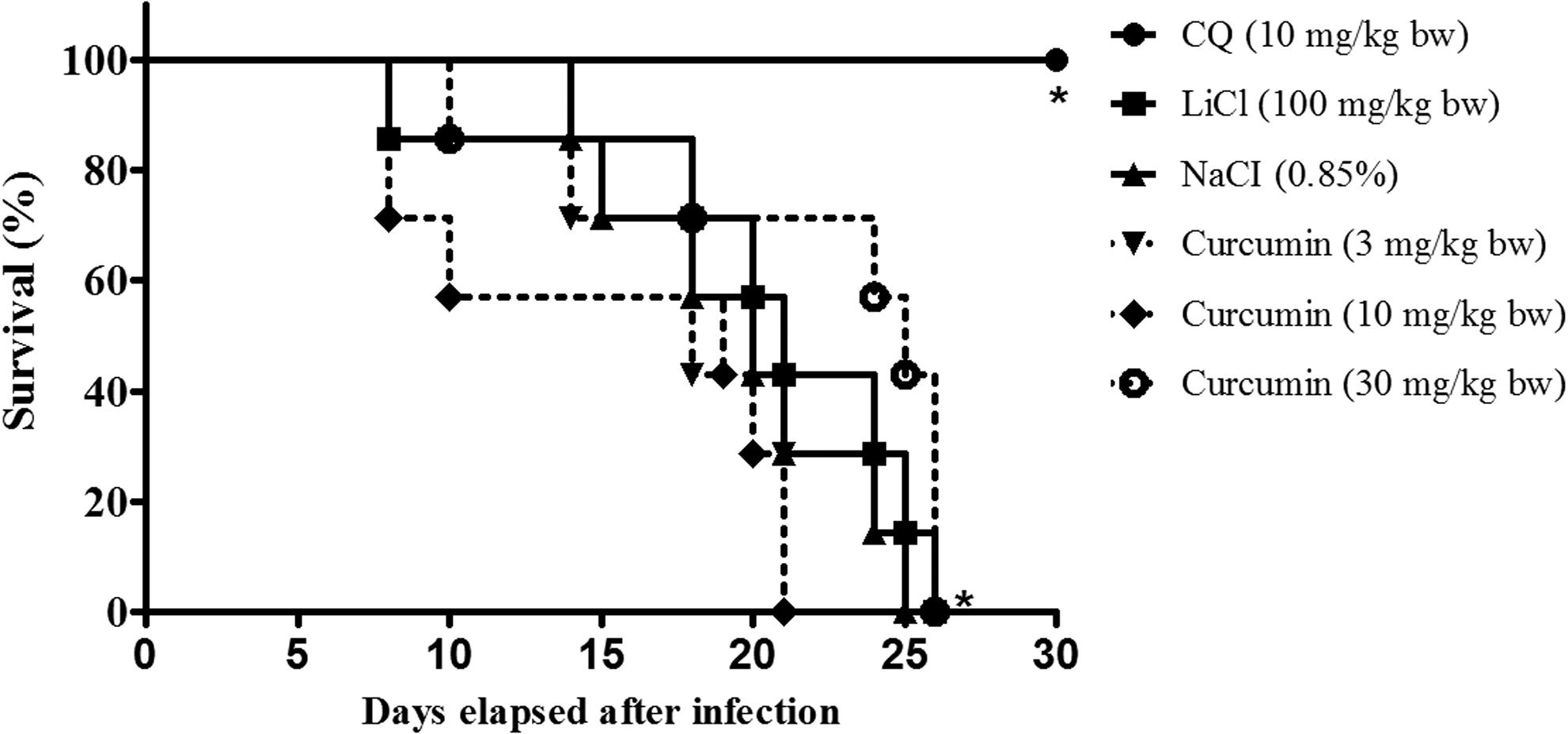
Representative Kaplan–Meier survival curve of mice infected with Plasmodium berghei NK65 infection with and without curcumin treatment at 3 h postinfection during therapeutic test. Data represent survival of negative control (nontreated P. berghei NK65-infected mice) (n = 7), curcumin-treated (n = 7), CQ-treated (n = 7), and LiCl-treated (n = 7) groups of experimental mice. Significant difference between tested and control groups was evaluated at P < .05 (*).
Parasitemia suppression was calculated on day-4 postinfection and survivability of mice recorded throughout the experimental period (30 days). Data represent mean ± SD for parasitemia suppression and median survival time (n = 7).
Significantly different from negative control at P < .05.
Significantly different from drug control (chloroquine) at P < .05.
bw, body weight; GSK3, glycogen synthase kinase-3.
Administration of curcumin for four consecutive days before injections with P. berghei-infected erythrocytes (prophylactic test) also showed dosage-dependent suppression of parasitemia development. However, at similar dosage tested during the suppressive test (30 mg/kg bw), curcumin moderately suppressed P. berghei parasitemia development in mice (chemosuppressive activity = 57.1% ± 1.3%). Median survival times of infected animals treated with curcumin were significantly higher from nontreated infected mice (Fig. 2). Control experimental animals given CQ showed 82.3% ± 0.8% suppression of parasitemia development on day-4 and survived throughout the observation period of 30 days (Table 2).
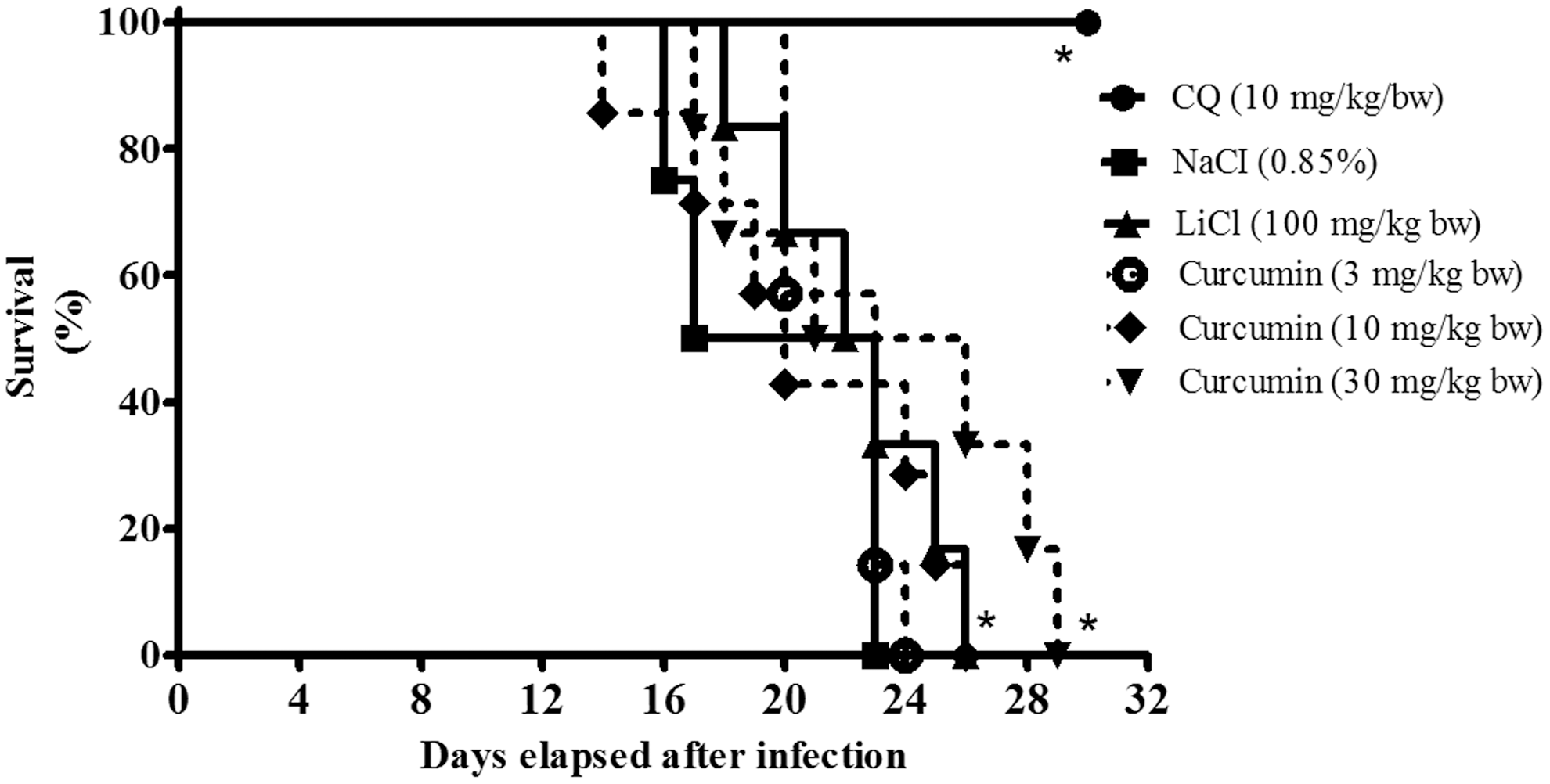
Representative Kaplan–Meier survival curve of mice infected with P. berghei infection with and without curcumin at 3 h preinfection during prophylactic test. Data represent survival of negative control (nontreated P. berghei NK65-infected mice) (n = 7), curcumin-treated (n = 7), CQ-treated mice (n = 7), and LiCl-treated mice (n = 7) groups. Significant difference between tested and control groups was evaluated at P < .05 (*).
Parasitemia suppression was calculated on day-4 postinfection and survivability of mice recorded throughout the experimental period (30 days). Data represent mean ± SD for parasitemia suppression and median survival time (n = 7).
Significantly different from negative control at P < .05.
Significantly different from drug control (chloroquine) at P < .05.
Both therapeutic and prophylactic treatments with curcumin resulted in increased phosphorylation of Akt (Ser473) and GSK3β (Ser9) in liver of P. berghei-infected mice
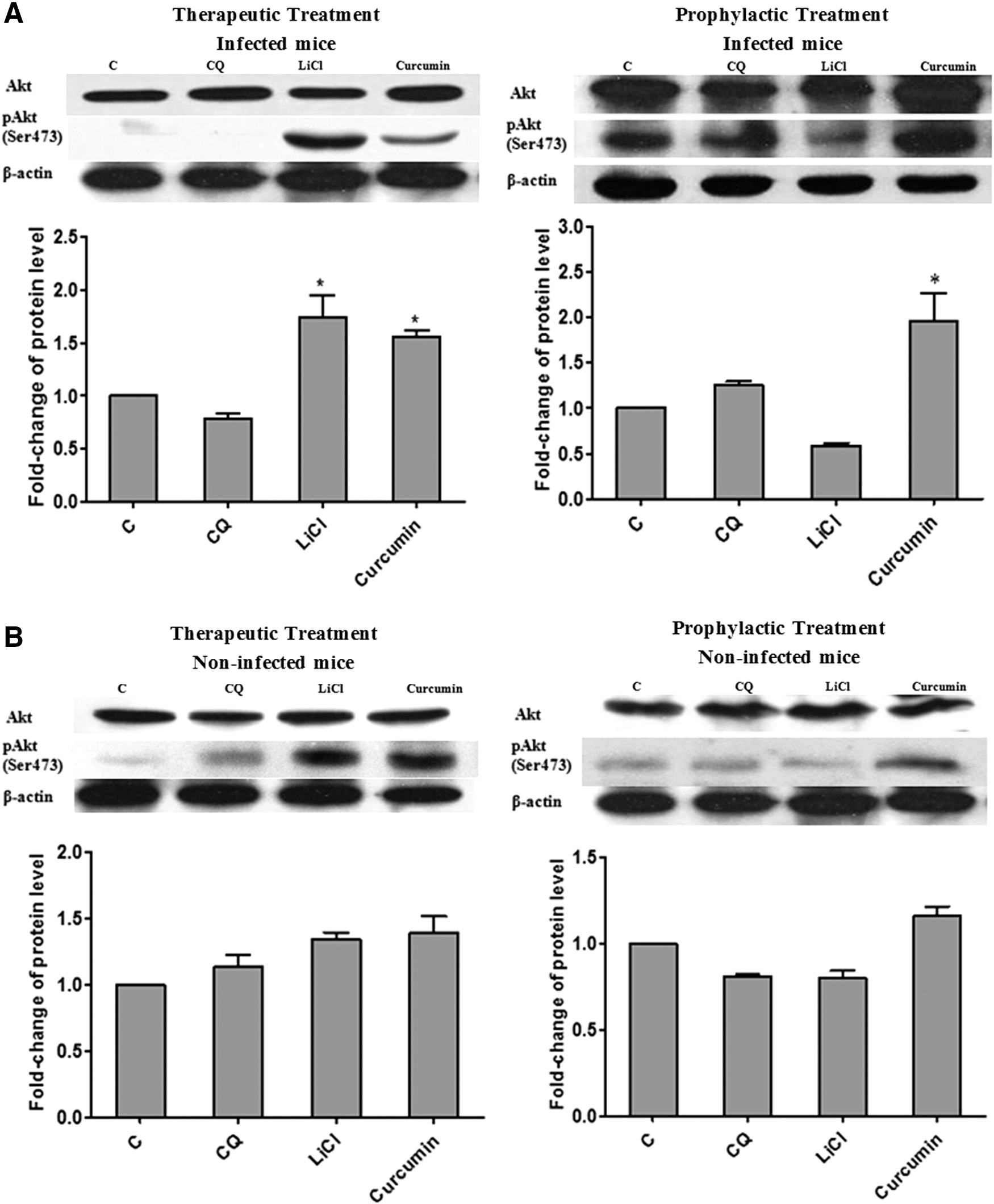
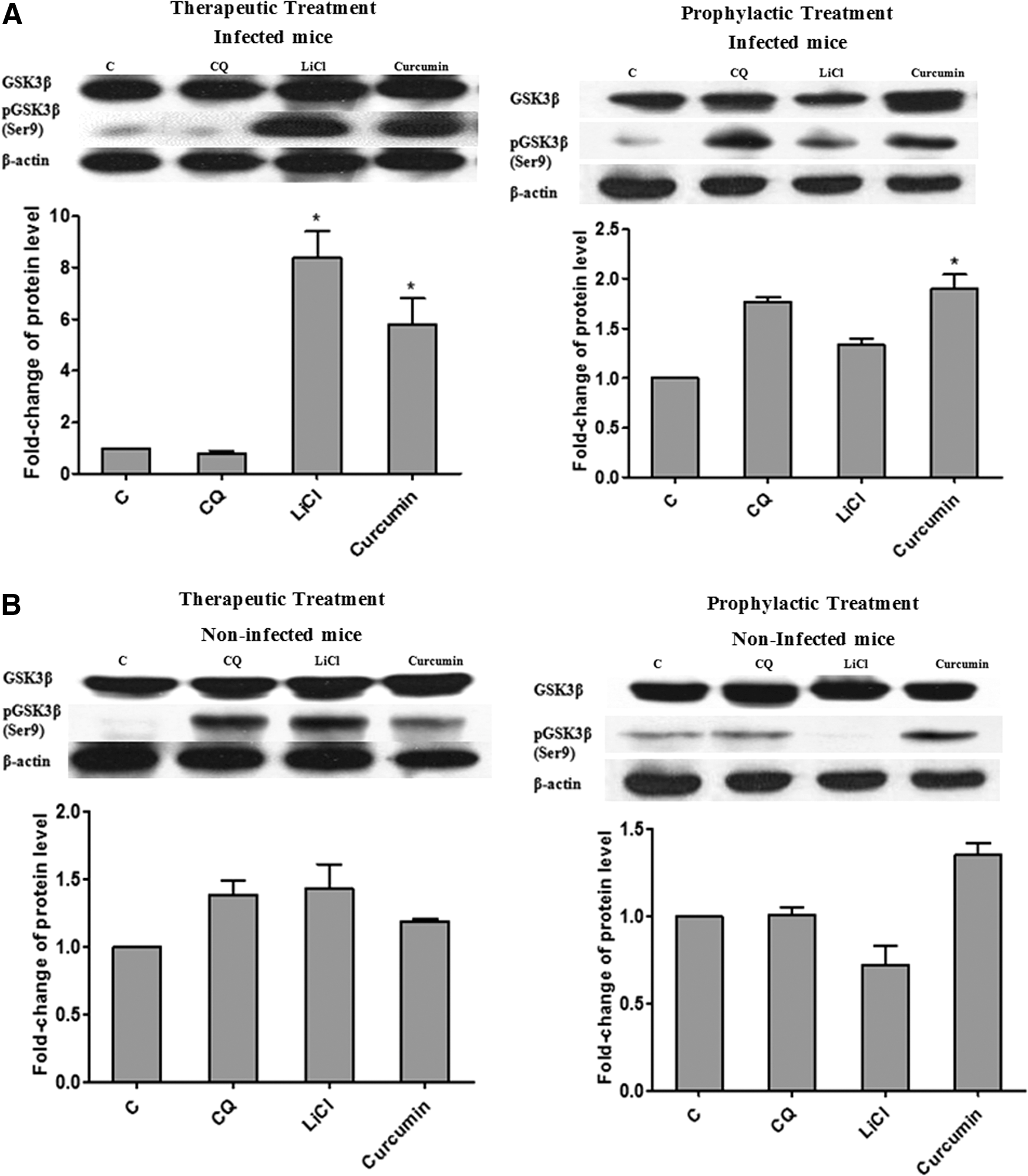
Akt plays a critical role in regulating and controlling glycogen synthesis, cell growth, and survival, whereas GSK3β is a critical downstream element of Akt whose activity can be inhibited by Akt-mediated phosphorylation at Ser9. 28 To further elucidate the potential mechanism of antimalarial effects for curcumin, we investigated the effects of curcumin on the Akt/GSK3 signaling pathway. Our results revealed that 30 mg/kg bw curcumin induced a significant (P < .05) increase in phosphorylation of Akt in liver by 1.6- and 1.7-fold for therapeutic and prophylactic tests, respectively (Fig. 3). Curcumin administration led to significantly increased (P < .05) levels of phosphorylated GSK3β (Ser9) in liver of P. berghei-infected mice compared to control group during therapeutic (5.5-fold) and prophylactic (1.8-fold) tests (Fig. 4). The fold-change observed in the curcumin-treated group was comparable to LiCl-treated group in the previous therapeutic test (8.0-fold). However for prophylactic test, the fold increase observed in curcumin-treated group was higher (1.8-fold) compared to LiCl-treated group (1.3-fold). There was no significant difference for either total Akt or GSK3β levels among all groups. These findings demonstrate that the activation of Akt led to the inhibition of host (liver) GSK3β upon treatment with curcumin for both therapeutic and prophylactic studies during malarial infection. The antimalarial effects of curcumin observed in this study suggest the involvement of GSK3β inhibition mediated through activation of Akt.
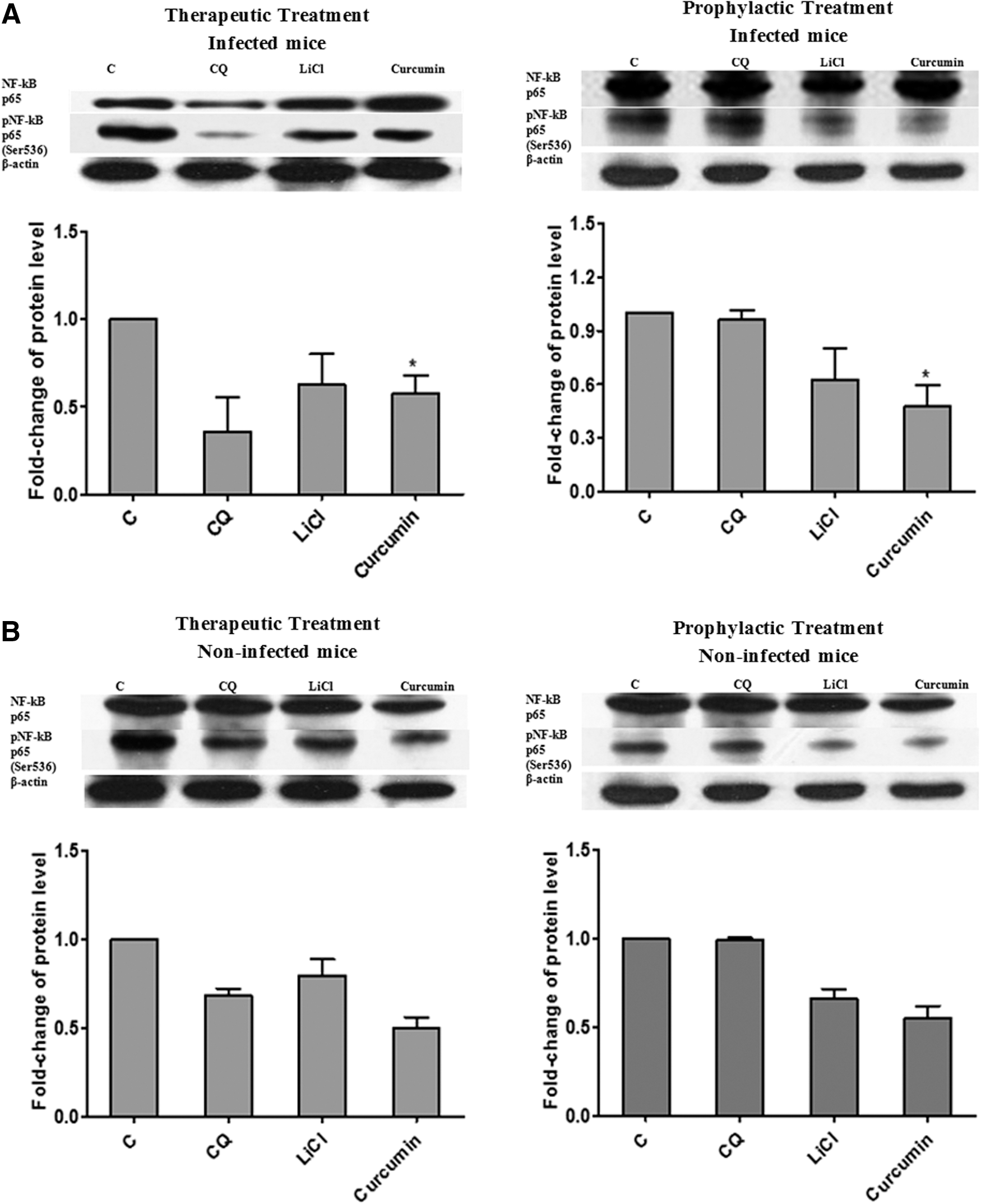
Both therapeutic and prophylactic treatments with curcumin inhibited the activation of NF-κB in liver of P. berghei-infected mice
As described above, curcumin was able to activate Akt by inducing Akt phosphorylation, which then led to inhibition of GSK3β. In this study, we investigated how GSK3β influenced the activation of NF-κB in malaria-infected mice treated with curcumin. Our results (Fig. 5) indicated that curcumin treatment decreased phosphorylation of pNF-κB p65 (Ser536) in liver of P. berghei-infected mice implying that curcumin caused inhibition of NF-κB activation. The level of pNF-κB p65 (Ser536) in liver of the curcumin-treated group was significantly (P < .05) decreased by 0.5-fold (for both therapeutic and prophylactic groups) compared to control. Similar inhibition effect was observed in mice treated with LiCl with 0.7-fold (therapeutic group) and 0.6-fold (prophylactic group) compared to control.
Administration of curcumin modulated pro- and anti-inflammatory cytokine levels through GSK3β inhibition in mice during therapeutic and prophylactic treatments
We next examined whether GSK3β inhibitory properties modulated serum levels of the cytokines during the course of curcumin treatments. The results presented in Figure 6 indicate that there was an increase in both pro- and anti-inflammatory cytokine levels on day-4 after P. berghei infection. During the therapeutic test, parasite infection increased the levels of pro- and anti-inflammatory cytokines TNF-α, IFN-γ, IL-10, and IL-4 by 7.5-, 35.0-, 33.0-, and 2.2-fold, respectively, in serum upon infection (Fig. 6). Treatment of mice with curcumin decreased the levels of pro-inflammatory cytokines TNF-α and IFN-γ (by 6.0- and 2.0-fold, respectively) and increased the levels of anti-inflammatory cytokine IL-10 and IL-4 (by 1.4-and 1.8-fold) in infected mice (Fig. 6). For prophylactic test, parasite infection increased the levels of pro- and anti-inflammatory cytokines TNF-α, IFN-γ, IL-10, and IL-4 by 3.7-, 30.0-, 18.0-, and 2.0-fold, respectively, in mice upon infection (Fig. 7). Treatment of mice with curcumin decreased the levels of pro-inflammatory cytokines TNF-α and IFN-γ (by 2.1- and 4.6- fold, respectively) and slightly increased the levels of anti-inflammatory cytokine IL-10 (by 1.1-fold) in infected mice (Fig. 7). However, treatment of mice with curcumin did not increase, but decreased the level of IL-4 (1.6-fold) in the prophylactic test. This shows that inhibition of GSK3β by curcumin modulated levels of pro- and anti-inflammatory cytokines in infected mice. As for mice treated with LiCl in the therapeutic test, levels of both TNF-α and IFN-γ decreased (by 5.0-fold) and level of IL-10 and IL-4 increased (by 1.6- and 5.6-fold) compared to infected controls. For mice-treated with LiCl in the prophylactic test, levels of TNF-α, IFN-γ, and IL-4 decreased (by 1.1-, 3.0-, and 2.5-fold) and level of IL-10 increased (by 1.2-fold) compared to infected controls. Levels of pro- and anti-inflammatory cytokines, TNF-α, IFN-γ, IL-10, and IL-4, in both therapeutic and prophylactic tests were decreased significantly by CQ treatment after infection. CQ may improve the evolution of the disease by decreasing the production of inflammatory cytokines compared to infected controls due to its ability to kill the parasites at the early days of infection. CQ has the ability to exert a direct effect on the immune system, similar to that occurring with quinine and mefloquine. 29,30
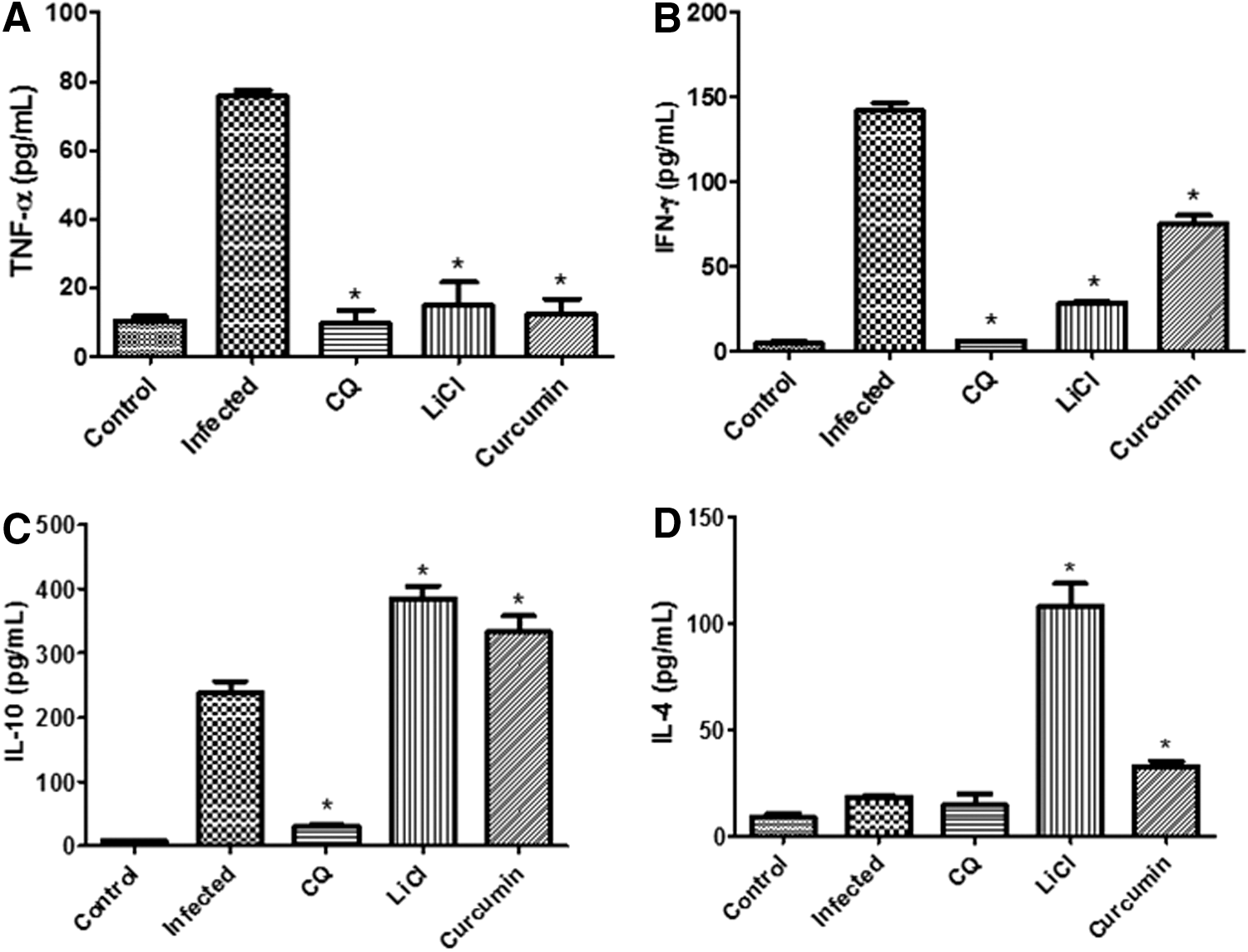
Pro-inflammatory cytokine levels of
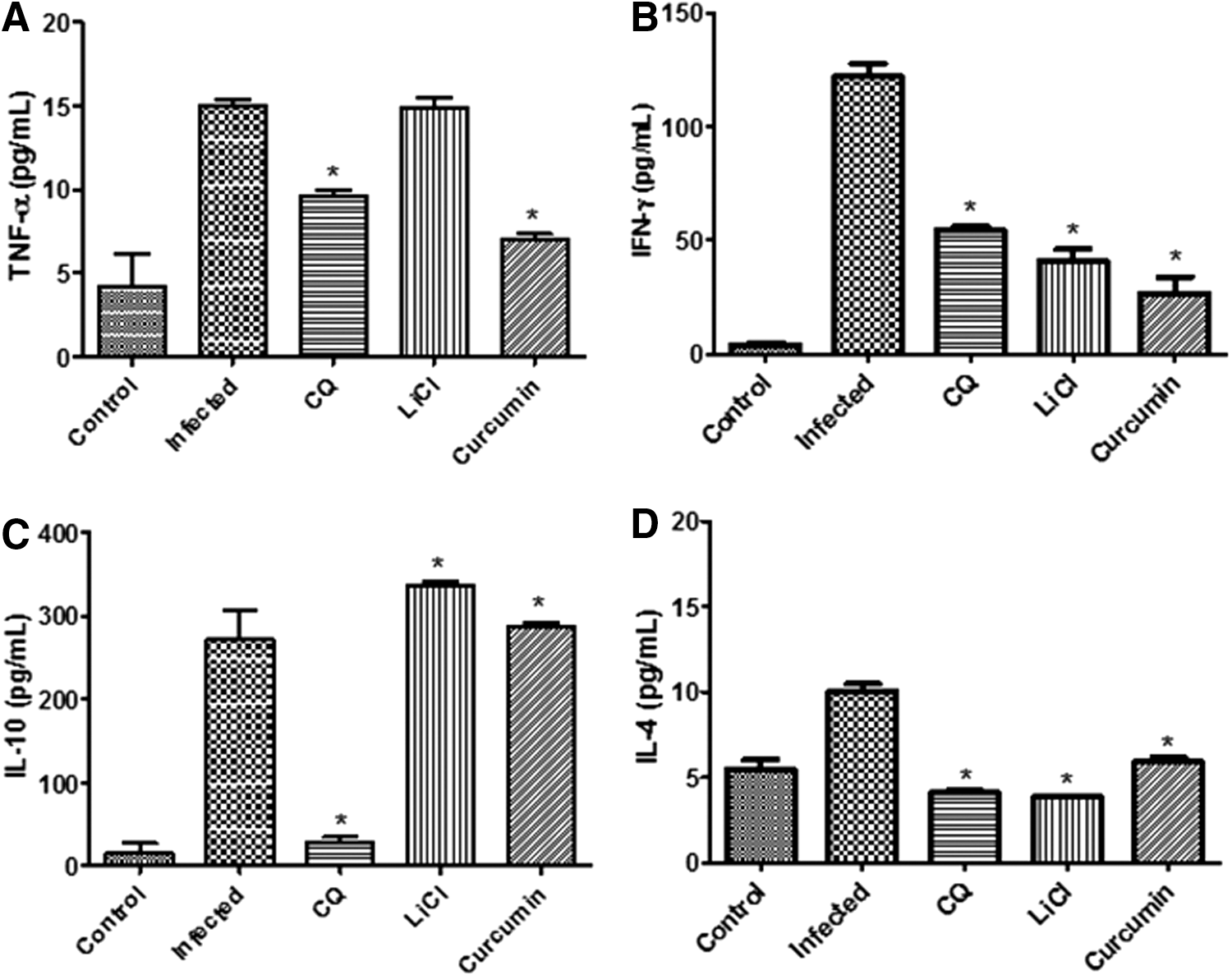
Pro-inflammatory cytokine levels of
Discussion
GSK3 is one of the pivotal modulators of cytokine levels during inflammation, among others in response to parasitic infections. 31 Severity of malaria is reputed to be partly due to overwhelming cytokine response to plasmodial infection. 8 Agents which exhibit immunomodulatory activities in the host may address the overwhelming cytokine response during infection and are potentially useful for treatment of malaria. 8
Our findings revealed that curcumin administration (therapeutic and prophylactic) into P. berghei-infected mice effectively suppressed parasitemia development in erythrocytes and improved survivability of infected animals, providing evidence that curcumin has good antimalarial properties. The present study is the first report on the antimalarial activity of the compound for uncomplicated malaria, although the antimalarial effects of curcumin has been previously reported for CM. 8 Curcumin treatment also resulted in significant decrease in levels of serum TNF-α and IFN-γ (pro-inflammatory cytokines) and increased levels of IL-10 and IL-4 (anti-inflammatory cytokines). At the same time, curcumin administration into malaria-infected mice resulted in Ser9 phosphorylation of liver GSK3 suggesting that the chemosuppressive effects of curcumin in vivo and the modulation of inflammatory cytokines described above are at least, in part, mediated by inhibition of GSK3β. It is noteworthy that our previous study showed LiCl treatment in malaria-infected mice, which also resulted in chemosuppression, modulation of cytokines, and Ser9 phosphorylation of liver GSK3, thus substantiating our previous report on the suppression of P. berghei parasitemia development by LiCl in an animal infection model. 20 Dai et al. also reported that LiCl treatment restored long-term neurocognitive function in ECM and suggested that this effect is attributed to dysregulation of GSK3β. 21
Modulatory effects of curcumin on cytokine levels have been widely documented for various inflammation-related conditions. 32 As examples, curcumin was reported to decrease lipolysaccharide (LPS)-induced TNF-α expression in microglial cells 33 and modulate TNF-α levels in a rat model of subchronic inflammation. 34 Another study revealed that curcumin protected rat liver from CCl4-induced injury and fibrogenesis by reducing levels of IFN-γ, TNF-α, and IL-6 in liver and serum. 35 It has also been demonstrated that curcumin can downmodulate the activation of NF-κB in macrophages and monocytes induced by various agents, including LPS during inflammation, 36,37 thereby suppressing the expression of TNF-α. Our findings indicate that the level of pNF-κB (Ser536) was lowered in liver of infected mice as a result of curcumin treatment. This implies that the inhibition of activation of NF-κB in liver of curcumin-treated infected mice is likely to be attributed to inhibition of GSK3β. Similar findings were obtained from previous studies on Francisella tularensis-infected mouse macrophages 38 and Streptococcus-infected mouse macrophage cell line 39 in which treatment with GSK3 inhibitors (including lithium) resulted in a decrease in the activation of NF-κB p65. NF-κB is one of the numerous transcription factors identified as molecular targets of curcumin. 40 In its regulatory role on inflammation, GSK3β is crucial in either promoting 41 or repressing 42 the activity of NF-κB. Overexpression of GSK3 during inflammation results in dysregulation of downstream processes; for example, in bacteria-driven inflammation dysregulation of NF-κB-mediated overproduction of pro-inflammatory cytokines has been associated with decreased GSK3β phosphorylation. 13
Our western analysis revealed that an upstream mediator of GSK3β, Akt, was also phosphorylated (at Ser473) in liver of infected mice for both therapeutic and prophylactic curcumin treatments. Akt is one of the protein kinases identified as a target of curcumin action. 40 Curcumin-associated activation (or phosphorylation) of Akt is not an uncommon phenomenon; curcumin administration resulted in Akt activation in heart muscle of diabetic rats, 43 in Michigan Cancer Foundation cells, 44 in rat aortic cells, 45 and in mouse β-cells. 46 Thus, our findings on the effect of curcumin administration on Akt are evidence that the Ser9 phosphorylation of liver GSK3β occurred as a consequence of Akt activation. However, this does not exclude the possibility that phosphorylation of GSK3β as a result of curcumin treatment may be due to the direct inhibitory effect of curcumin, which was suggested by an in silico modeling experiment. 22 Overall, our findings on the effect of curcumin on the activation of Akt, inhibition of GSK3, and inhibition of NF-κB activation suggest a crucial role of GSK3β in the modulation of inflammatory cytokines in malaria-infected mice.
Taken together, findings described above on host response to curcumin treatment demonstrate that inhibition of host GSK3 is, in part, involved in the underlying mechanism of the antimalarial effect of curcumin. However, this may not fully explain the antimalarial effects of curcumin observed in the in vivo infection study since the effects encompassed both host factors and intrinsic antiparasitic (chemosuppressive) activity. Our in vitro studies also revealed that curcumin displayed potent and selective antiplasmodial activity and, therefore, may contribute to the overall antimalarial activity observed in vivo.
We have previously shown that the antimalarial activity of kaempferol, an important anti-inflammatory flavonoid of plant origin, is also mediated by inhibition of GSK3β. 47 Thus, the findings of the present study represent another example of an antimalarial compound of plant origin acting through inhibition of GSK3, which has not been reported before this.
Footnotes
Acknowledgment
This research was supported by a research university grant from Universiti Kebangsaan Malaysia (UKM-ETP-2013-052).
Author Disclosure Statement
No competing financial interests exist.