
Abstract
The nonstructural protein NS1 of human parvovirus B19 (B19) is known to exacerbate disease activity in systemic lupus erythematosus (SLE). However, no specific medicine for B19 infection is available. The roots of Gentiana macrophylla Pall. (GM), the traditional Chinese medicine “Qinjiao,” have been used for centuries to treat rheumatic disease, including SLE. Herein, we aimed to investigate the effects of GM root extract (100 and 300 mg/kg body weight) on B19-NS1-exacerbated liver injury in NZB/W F1 mice; liver tissues were assessed by hematoxylin–eosin staining and immunoblotting. The GM root extract significantly decreased B19-NS1-exacerbated liver inflammation by suppressing the expressions of hepatic inducible nitric oxide synthase, cyclooxygenase type 2 (COX-2), interleukin (IL)-1β proteins, values of serum asparate transaminase (AST) and alanine transaminase (ALT), and lymphocyte infiltration (P < .05). It also significantly reduced the B19-NS1-exacerbated hepatic matrix metalloproteinase-9 (MMP-9) and urokinase plasminogen activator (uPA) expressions by downregulating tumor necrosis factor (TNF)-α/NF-κB (p65) signaling. These findings suggest a therapeutic potential of GM root extract against B19-NS1-exacerbated liver inflammation in SLE.
Introduction
S
Complementary and alternative medicine (CAM) is a combination of traditional practices and beliefs. Tibetan medicine has been in Chinese traditional medical science for thousands of years. 12 Gentiana and Swertia are two major species of the 22 species of Gentianaceae, which only grow in the high alpine areas of the Tibetan Plateau. 13 Gentiana macrophylla Pall. (GM) is a perennial herb that belongs to the Radix Gentianaceae and has been utilized to treat a wide range of diseases, including diabetes, apoplexy, paralysis, and stomachache. 14 The roots of GM are known as the traditional Chinese medicine “Qinjiao” and are commonly used to treat pain and inflammation. 13,15 GM has been demonstrated to alleviate both CCl4-induced and lipopolysaccharide (LPS)-induced liver injuries by inhibiting the production of tumor necrosis factor (TNF)-α and is known to improve blood circulation and remove blood stasis in patients with reflex sympathetic dystrophy. 16,17 GM also reportedly alleviates the inflammatory activity in a carrageenan-induced paw edema model 18 and is effective for treating patients with rheumatoid arthritis or SLE. 19,20
In recent decades, the use of CAM appears to have been increasing. 19 However, the therapeutic potential of GM root extract against B19-NS1-exacerbated disease activity in SLE remains unclear. This study investigates whether GM root extract alleviates B19-NS1-exacerbated liver injuries in lupus-prone mice and, if so, elucidates the underlying mechanisms.
Materials and Methods
Preparation of GM root extract
Root extract of GM was prepared as described in our previous report. 20 Briefly, the roots of GM were purchased from a domestic traditional herb store (Chen-Oriental-Lin Ginseng Medicine Store, Taishan District, New Taipei City, Taiwan), ground into a powder and soaked in 80% ethanol at room temperature for 1 week. The solution was then centrifuged at 20,000 g for 30 min at 4°C to remove the precipitate and filtered through two layers of gauze (0.45 mm). The extract solution was then lyophilized in a freeze-dry system and stored at −80°C until use.
Chemicals and standards
Standards of phenolic acid, including gallic, gentisic, chlorogenic, p-hydroxybenzoic, vanillic, caffeic, p-coumaric, ferulic, sinapic, syringic, p-anisic and rosmarinic, and flavonoid, including catechin, epicatechin, rutin, naringin, myricetin, hesperidin, quercitrin, neohesperidin, eriodictyol, diosmin, morin, daidzein, quercetin, glycitein, naringenin, luteolin, genistein, hesperetin, kaempferol, apigenin, and isorhamnetin, were purchased from Sigma Co. (St. Louis, MO, USA). Methanol was obtained from Merck Co. (Darmstadt, Germany). Distilled deionized water (ddH2O) was prepared using an Ultrapure™ water purification system (Lotun Co., Ltd. Taipei, Taiwan). The total phenol contents were determined using a method that involved Folin–Ciocalteu's phenol reagent with gallic acid as a standard and expressed as mg gallic acid equivalent (GAE)/100 mL extract. 21 Total flavonoid contents were measured by a method that involved 10% AlCl3·H2O solution with (+)-catechin as a standard and expressed as mg catechin equivalent (CE)/100 mL extract. 21
Compositions and contents of flavonoids and phenolic acids in GM root extract
The compositions and contents of flavonoids and phenolic acids in GM were analyzed as described elsewhere, 22 yielding the results in Figure 1 and Table 1. Briefly, a high-performance liquid chromatography (HPLC) system that comprised a PrimeLine™ Gradient Model 500G HPLC pump system (Analytical Scientific Instruments, Inc., El Sobrante, CA, USA) and an S-3210 photodiode-array detector (Schambeck SFD GmbH, Bad Honnef, Germany) was used. A Hypersil GOLD C18 column (250 × 4.6 mm, 5 μm; Thermo Fisher Scientific, Inc.) and a gradient solvent system that consisted MeOH (solvent A) and ddH2O with 9% glacial acetic acid (solvent B) (conditions: 5–17% A from 0 to 5 min, and 17% A maintained from 5 to 25 min; 17–31% A from 25 to 40 min, and 31% A maintained from 40 to 76 min; 31–40% A from 76 to 80 min, and 40% A maintained from 80 to 120 min; flow rate = 0.8 mL/min) were used to separate components, whose UV spectra were recorded from 220 to 450 nm.
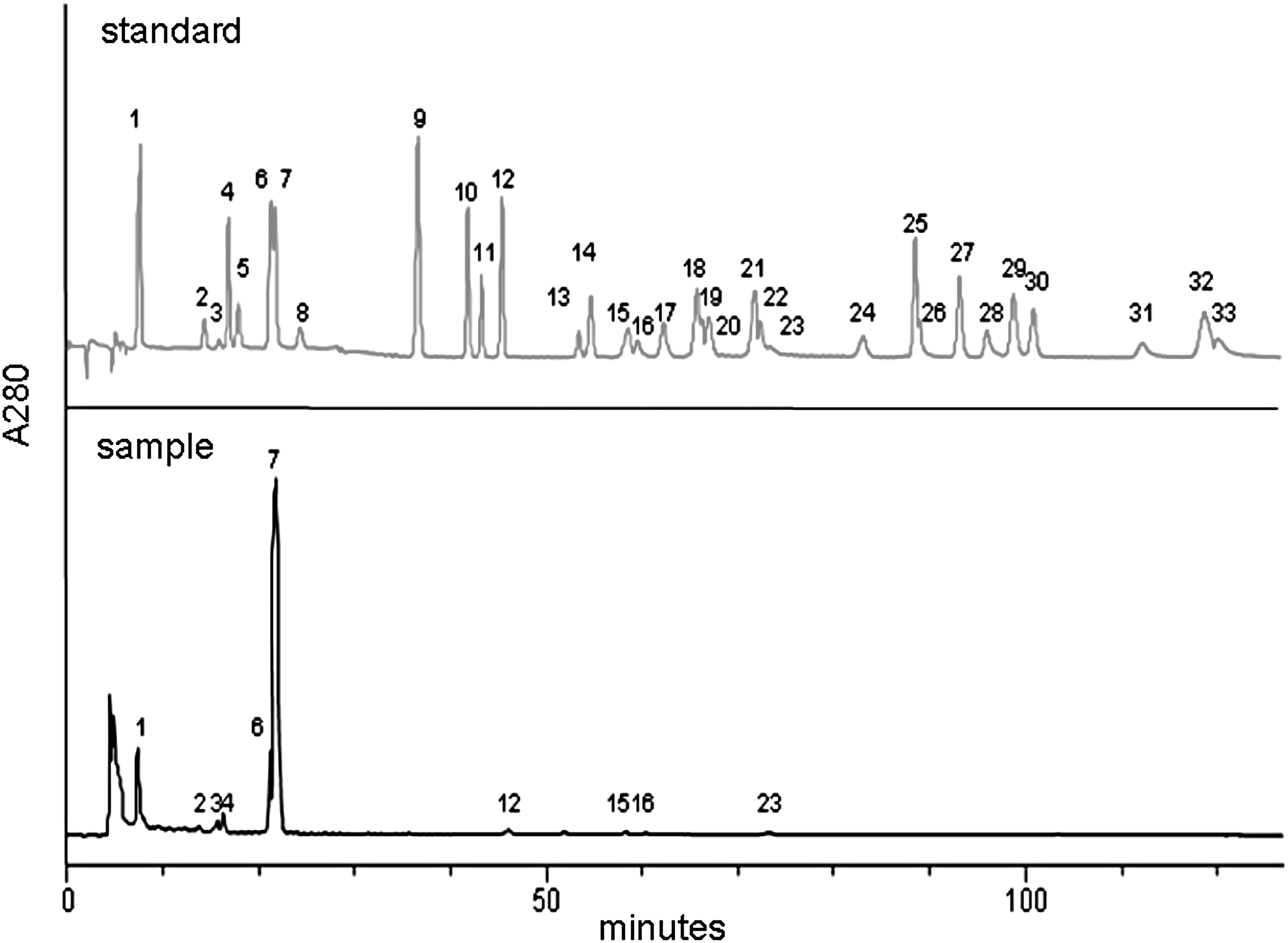
Chromatograms of flavonoids and phenolic acids in Gentiana macrophylla Pall. ethanol root extract solution. 1 = gallic acid, 2 = catechin, 3 = gentisic acid, 4 = chlorogenic acid, 5 = p-hydroxybenzoic acid, 6 = vanillic acid, 7 = caffeic acid, 8 = epicatechin, 9 = p-coumaric acid, 10 = ferulic acid, 11 = sinapic acid, 12 = syringic acid, 13 = rutin, 14 = p-anisic acid, 15 = naringin, 16 = myricetin, 17 = hesperidin, 18 = rosmarinic acid, 19 = quercitrin, 20 = neohesperidin, 21 = eriodictyol, 22 = diosmin, 23 = morin, 24 = daidzein, 25 = quercetin, 26 = glycitein, 27 = naringenin, 28 = luteolin, 29 = genistein, 30 = hesperetin, 31 = kaempferol, 32 = apigenin, 33 = isorhamnetin.
Preparation of recombinant B19-NS1 viral proteins
The recombinant human parvovirus B19 proteins were prepared as described elsewhere. 7,10 The B19-NS1 protein was purified using Profinia denaturing IMAC purification kits and the Profinia protein purification system (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Animal and treatments
This study was approved by the Institutional Animal Care and Use Committee at Chung Shan Medical University (IACUC Approval No. 1456). Thirty-two female NZB/W F1 mice aged 5 weeks were purchased from Jackson Laboratory (Bar Harbor, ME, USA) and housed under the supervision of the Institutional Animal Care and Use Committee at Chung Shan Medical University, Taichung, Taiwan. The animals were kept in a 12-h light/12-h dark cycle, and ambient temperature was maintained at 25°C. All animals had free access to water and standard laboratory chow (Lab Diet 5001; PMI Nutrition International Inc., Brentwood, MO, USA). Animal welfare and experimental procedures were performed according to the NIH Guide for the Care and Use of Laboratory Animals. All animals at week 8 of age were randomly divided into four groups (each of eight mice) and treated as follows: (1) G1: 1 mL phosphate-buffered saline (PBS) injection+vehicle treatment (p.o.), (2) G2: B19-NS1 injection+vehicle treatment (p.o.), (3) G3: B19-NS1 injection +100 mg GM/kg BW (p.o.), and (4) G4: B19-NS1 injection +300 mg GM/kg BW (p.o.). The dosage of GM root extract used in this study was described elsewhere. 23 Subcutaneous injections were performed with PBS or 20 μg purified B19-NS1 recombinant protein. All injections were boosted twice a week. The experimental period was 8 weeks, and the mice were sacrificed by CO2 asphyxiation at the age of 16 weeks. Liver tissues were collected and stored at −80°C until use.
Hematoxylin–eosin staining
Hematoxylin–eosin staining was performed as described elsewhere. 20,24 Liver samples from the animals were excised and soaked in formalin and imbedded in wax. The waxed tissue blocks were cut into 5-μm-thick sections and prepared by deparaffinization and dehydration. The sections were passed through a series of graded alcohols (100%, 95%, and 75%) for 15 min each and then dyed with hematoxylin. Photomicrographs were obtained using Zeiss Axiophot microscopes.
Immunoblotting
The liver tissues obtained as described above were homogenized in 600 μL PRO-PREP solution (iNtRON Biotech, Gyeonggi-do, Korea) by 30 strokes using a Dounce Homogenizer (Kontes Glass, Vineland, NJ, USA). The whole tissue homogenates were centrifuged at 19,083 g for 10 min at 4°C to discard the insoluble fraction. The protein content in the supernatant was measured using the Bio-Rad Protein Assay Kit (Bio-Rad Laboratories, Inc.) according to the manufacturer's instructions. Subsequently, the protein samples were separated in 10% or 12.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electrophoretically transferred to the nitrocellulose membrane (Amersham Biosciences, Piscataway, NJ, USA), as described elsewhere. 11,20 After blocking with 5% nonfat dry milk in 1 × PBS, antibodies against inducible nitric oxide synthase (iNOS), cyclooxygenase type 2 (COX-2), interleukin (IL)-1β, TNF-α, TNF-α receptor, IKK-α, IκB, p-nuclear factor (NF)-κB p65, matrix metalloproteinase-9 (MMP-9), urokinase plasminogen activator (uPA) (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and β-actin (Chemicon; EMD Millipore, Temecula, CA, USA) were diluted in PBS that contained 2.5% bovine serum albumin and incubated for 1.5 h at room temperature with gentle agitation. The membranes were washed twice with PBS-Tween for 1 h, and a secondary antibody, conjugated with horseradish peroxidase (HRP) (Santa Cruz Biotechnology), was added. Immobilon Western HRP Chemiluminescent Substrate (EMD Millipore) was then used to detect the antigen antibody complexes. The blots were scanned and semiquantified by densitometry (Alpha-Imager 2200; ProteinSimple, San Jose, CA, USA).
Detection of AST and ALT
The mice were sacrificed as described above, and the heart sera were collected by direct puncture with a sterilized 1 mL syringe. The serum value of asparate transaminase (AST) and alanine transaminase (ALT) was detected by a nephelometric system according to the manufacturer's instruction (Cobas Mira, Roche, Swiss).
Statistical analyses
All statistical analyses were performed with SPSS 10.0 software (SPSS, Inc., Chicago, IL, USA). Three independent experiments were repeated. Statistical analyses were performed with the Student's t-test and one-way ANOVA. P < .05 was considered statistically significant.
Results
Phytochemical contents in GM root extract
The contents of total phenols and flavonoids in GM root extract were 100.49 ± 5.92 (mg GAE/g dry weight of GM root extract) and 11.71 ± 0.87 (mg CE/g dry weight of GM root extract), respectively. Flavonoids, such as catechin, naringin, myricetin, and morin, and phenolic acids, including gallic acid, gentisic acid, chlorogenic acid, vanillic acid, caffeic acid, and syringic acid, were found in the GM root extract (Fig. 1). Morin (4.19 ± 0.04 mg/mL extract) and caffeic acid (51.34 ± 2.15 mg/mL extract) were the most abundant flavonoid and phenolic acid, respectively, in the GM root extract (Table 1).
Effects of GM root extract on lymphocyte infiltration and hepatic inflammatory indicators in NZB/W F1 mice that received B19-NS1
Anti-inflammatory effects of GM root extract were evaluated in livers of NZB/W F1 mice challenged with B19-NS by measuring lymphocyte infiltration and various hepatic inflammatory markers. Considerably increased lymphocyte infiltration was observed in the livers of NZB/W F1 mice that had received B19-NS1 protein (G2), relative to those in the G1 group (Fig. 2A). Considerably less lymphocyte infiltration was detected in the livers of the NZB/W F1 mice that had received both B19-NS1 and GM root extract (G3 and G4), relative to those in the G1 and G2 groups. Significantly increased iNOS and COX-2 protein levels were detected in livers from NZB/W F1 mice that had received B19-NS1 (G2), relative to those in the G1 group (Fig. 2B). In contrast, significantly lower iNOS and COX-2 protein levels were detected in the livers of NZB/W F1 mice that had received both B19-NS1 and GM root extract (G3 and G4), relative to those in the G1 and G2 groups, respectively (Fig. 2B). Figure 2C presents quantitative results concerning iNOS and COX-2 protein levels relative to β-Actin.

Hepatic histopathological changes and protein levels of iNOS and COX-2.
Significantly greater IL-1β protein levels were detected in livers from NZB/W F1 mice that had received B19-NS1 (G2), relative to those in the G1 group (Fig. 3A). Conversely, significantly lower IL-1β protein levels were also detected in the livers of NZB/W F1 mice that had received both B19-NS1 and GM root extract (G3 and G4), relative to those in the G1 and G2 groups, respectively (Fig. 3B). In addition, significantly increased values of serum AST and ALT were also detected in NZB/W F1 mice that had received B19-NS1 (G2), relative to those in the G1 group. Conversely, significantly reduced serum AST and ALT were detected in NZB/W F1 mice that had received both B19-NS1 and GM root extract (G3 and G4), relative to those in the G1 and G2 groups, respectively (Fig. 3C).

Expressions of IL-1β and activities of serum AST and ALT.
Effects of GM root extract on signaling involved in B19-NS1-exacerbated liver injury in NZB/W F1 mice
To elucidate the effects of GM root extract on signaling involved in B19-NS1-exacerbated liver injury in NZB/W F1 mice, the expressions of TNF-α, TNF-α receptor, IKK-α, IκB, NF-κB (p65), MMP-9, and uPA proteins were examined. Significantly greater TNF-α and TNF-α receptor expressions were detected in livers from NZB/W F1 mice that had received B19-NS1 (G2), relative to those in the G1 group (Fig. 4A). Significantly lower TNF-α and TNF-α receptor expressions were observed in livers of NZB/W F1 mice that received both B19-NS1 and GM root extract (G3 and G4), relative to those in the G1 and G2 groups, respectively (Fig. 4A).
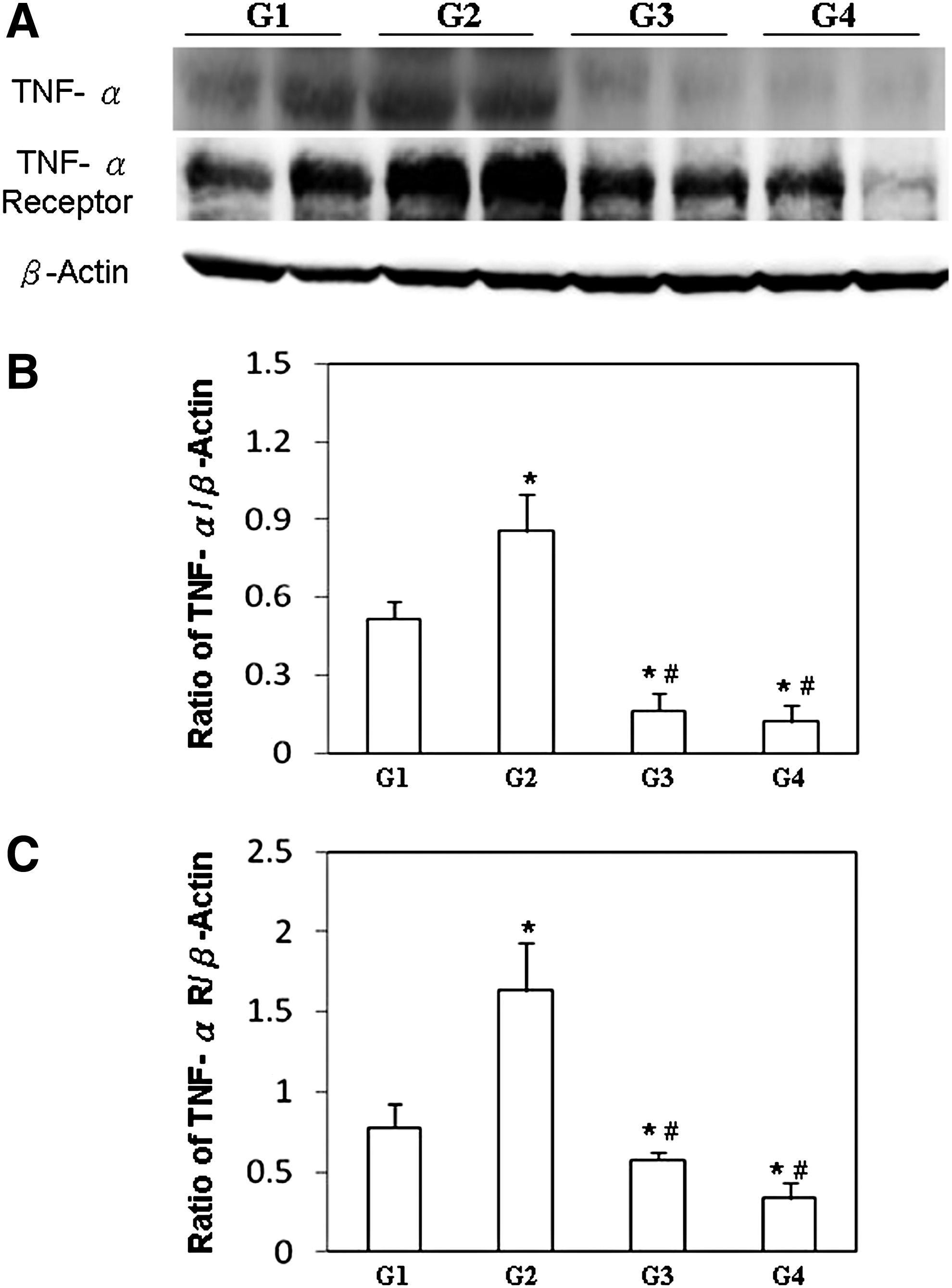
Expressions of TNF-α and TNF-α receptor.
The downstream molecules of TNF-α/TNF-α receptors, including IKK-α, IκB, and NF-κB (p65), were significantly increased in livers of NZB/W F1 mice that received B19-NS1 (G2), relative to those in the G1 group (Fig. 5A). Significantly lower IKK-α, IκB, and NF-κB (p65) levels were detected in the livers of NZB/W F1 mice that received both B19-NS1 and GM root extract (G3 and G4), relative to those in the G1 and G2 groups, respectively (Fig. 5A). Figure 5B–D display quantitative results concerning IKK-α, IκB, and NF-κB (p65) protein levels relative to β-Actin, respectively.
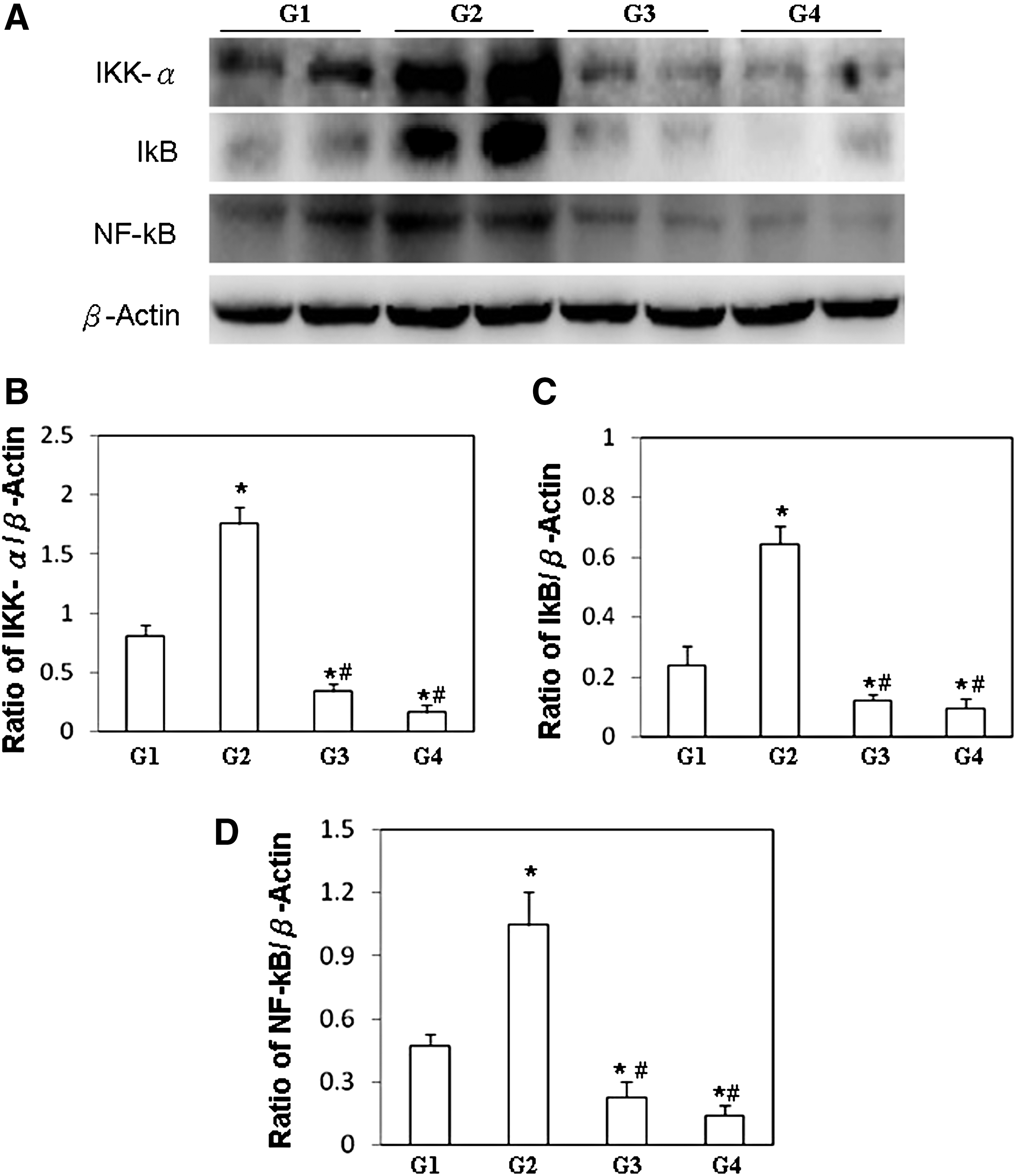
Expressions of IKK-α, IκB, and NF-κB p65.
Accordingly, significantly higher MMP-9 and uPA levels were detected in livers from NZB/W F1 mice that received B19-NS1 (G2), relative to those in the G1 group (Fig. 5). Significantly lower expressions of MMP-9 and uPA were detected in livers from NZB/W F1 mice that received both B19-NS1 and GM root extract (G3 and G4), relative to those in the G1 and G2 groups, respectively (Fig. 6A). Figure 6B and C display MMP-9 and uPA protein levels relative to β-Actin, respectively.

Expressions of MMP-9 and uPA.
Discussion
Increasing evidence suggests that human parvovirus B19 infection is strongly associated with the onset and exacerbation of SLE. 3,25 Hence, the infection of B19 presents a clinical and serological tableau such as that in an episode of SLE may include hypocomplementemia, cytopenia, and the generation of autoantibodies. 4 Indeed, B19 infection has been reported to aggravate disease activity in both SLE patients and animal models. 3,10,11 However, no specific antiviral drug is available (for the treatment of parvovirus B19 infection). 3,26 This study revealed beneficial effects of GM root extract on B19-NS1-exacerbated hepatic injury in NZB/W F1 mice, which were due to its suppression of iNOS and COX-2 protein expression and lymphocyte infiltration. GM root extract also significantly decreased B19-NS1-exacerbated hepatic MMP-9 and uPA expressions.
Oxidative and inflammatory stresses, such as elevated TNF-α, iNOS, and COX-2, have been known to be pivotal in the pathogenesis of SLE and are associated with the SLE Disease Activity Index (SLEDAI). 27 –29 In SLE, TNF-α and its downstream molecules, IKK and IκB, are known to be involved in the signaling pathway of MMP-9 induction, 30 which degrades the extracellular matrix in inflammatory responses. 31 Indeed, MMP-9 has also been associated with the pathogenesis of SLE. 24,32 Elevated MMP-9 activity has been demonstrated to cleave myelin basic protein or type II gelatins and generate remnant epitopes, which contribute to the development of autoimmunity. 33 Notably, these phenomena are exacerbated by the receiving of B19-NS1 proteins. 10,11 Although no specific treatment for B19-NS1-exacerbated disease activity in SLE is available, the present study reveals that GM root extract significantly decreases hepatic MMP-9 and uPA expressions in NZB/W F1 mice by downregulating TNF-α/NF-κB signaling, suggesting the therapeutic potential of GM root extract against B19-NS1-exacerbated liver injury in SLE.
Increasing attention has focused on the health benefits of flavonoids, which are phenolic compounds with a C6-C3-C6 backbone structure. Flavonoids are found in a wide range of plants and have numerous biochemical and pharmacological effects, including antioxidant, antiviral, antiallergic, and anti-inflammatory effects. 34 Although the cellular mechanisms that are associated with the activities of polyphenols and their metabolites have not been completely elucidated, such functions as antioxidant activity, free radical scavenging, and anti-inflammation are believed to be involved. 35 Gentiopicroside, a dominant constituent of GM, has a potent protective effect on the IL-1β-induced inflammatory response in rat articular chondrocyte and may have efficacy as a therapeutic strategy against osteoarthritis. 15 Furthermore, numerous biological activities of caffeic acid, such as antioxidant, anti-inflammatory, antibacterial, and anticarcinogenic activities, have been reported, 36,37 and these are recognized as being associated with radical scavenging, metal ion chelation, and the inhibition of enzymes. 38 Morin hydrate has an upregulating and immunoprotective effect. Indeed, the gastroprotective action of morin is primarily attributed to its potent antioxidant and anti-inflammatory functions. 39 Morin significantly increases splenocyte proliferation and phagocytic capacity and inhibits the production of proinflammatory cytokines and nitric oxide and reduces complement activities. 40 The fact that morin and caffeic acids are the flavonoid and phenolic acids that are most abundant in GM root extract, according to the results in this study, may explain the effectiveness of GM root extract for ameliorating B19-NS1-exacerbated liver inflammation in NZB/W F1 mice. However, further investigations are required to identify the other functional constituents of GM root extracts.
The current study provides evidence that the administration of GM root extract significantly ameliorates B19-NS1-exacerbated liver injury in NZB/W F1 mice by reducing hepatic lymphocyte infiltration, iNOS, COX-2, MMP-9 and uPA expressions, and TNF-α/NF-κB (p65) signaling. These findings suggest that GM root extract alleviates B19-NS1-exacerbated inflammation in lupus-prone mice and offers a possible remedy against liver injury for SLE patients, especially SLE patients with B19 infection.
Footnotes
Acknowledgments
This study was funded by the Chung Shan Medical University and Chi-Mei Medical Center cooperative project CSMU-CMMC-104-01. Consumptive materials were partially supported by the National Science Council grant NSC99-2320-B-040-007-MY3 and NSC101-2314-B-040-008. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article. Ted Knoy is appreciated for his editorial assistance.
Author Disclosure Statement
No competing financial interests exist.