
Abstract
To increase the functionality of Opuntia ficus-indica var. saboten cladodes, it was fermented by Lactobacillus plantarum and Bacillus subtilis. Eighty percent methanol extracts were investigated for their effects on nitric oxide (NO) production, cytokine secretion, nuclear factor-κB (NF-κB) activity, and mitogen-activated protein kinase (MAPK) phosphorylation in RAW 264.7 cells. Methanol extracts of L. plantarum culture medium (LPCME) and B. subtilis culture medium (BSCME) did not affect lipopolysaccharide (LPS)-induced NO production but, at 500 μg/mL, increased interferon (IFN)-γ-induced NO production by 55.2 and 66.5 μM, respectively, in RAW 264.7 cells. In RAW 264.7 cells not treated with LPS and IFN-γ, LPCME did not affect NO production, but BSCME increased NO production significantly in a dose-dependent manner. In addition, BSCME induced the expression of tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) in RAW 264.7 cells in a dose-dependent manner. BSCME at 500 μg/mL increased TNF-α and IL-1β mRNA levels by 83.8% and 82.2%, respectively. BSCME increased NF-κB-dependent luciferase activity in a dose-dependent manner; 500 μg/mL BSCME increased activity 9.1-fold compared with the control. BSCME induced the phosphorylation of p38, c-JUN NH2-terminal protein kinase (JNK), and extracellular signal-regulated kinase (ERK) in a dose-dependent manner, but did not affect total ERK levels. In conclusion, BSCME exerted immunostimulatory effects, which were mediated by MAPK phosphorylation and NF-κB activation, resulting in increased TNF-α and IL-1β gene expression in RAW 264.7 macrophages. Therefore, BSCM shows promise for use as an immunostimulatory therapeutic.
Introduction
O
Macrophage activation, the transformation to an effector cell, occurs during both natural (innate) and specific (adaptive) immune reactions to infection in immunocompetent hosts. It is induced by interferon (IFN), a cytokine released mainly by appropriately stimulated natural killer (NK) or T cells. Activation of microbicidal mechanisms in macrophages may also result from exposure to immunomodulating agents. 11 Tumor necrosis factor-α (TNF-α) is an important regulator of inflammatory and immune responses. Both of these factors are associated with the nuclear factor-κB (NF-κB) and mitogen-activated protein kinase (MAPK) signaling pathways. 12
Probiotics are live microorganisms that confer a beneficial health effect to the host. The bacteria genera most commonly used in probiotic manupulations are Lactobacillus, Streptococcus, Enterococcus, Bifidobacterium, and Bacillus. Probiotics play an important role in regulating the immune system. The clinical effects of Bacillus subtilis as an agent with immunostimulatory effect in a variety of diseases in human and animals have been documented. 13
Fermentation is a popular food-processing method used to generate biologically active substances from raw materials and/or fermentation products. Yeasts and bacteria are used to produce fermented foods, such as bread, sausages, fermented milk, and fruit beverages. Fermentation also improves the nutritional quality of food and increases its safety by reducing the levels of various toxic compounds. Fermented products have high antioxidant activity, 14 anti-inflammatory activity, 15 and immunomodulatory effects. 16 Ginseng flower buds fermented by B. subtilis had high ginsenoside contents and exerted greater immunomodulatory effects than nonfermented buds. 17 However, the immunological activity of fermented OFSCs has not been reported to date.
In this study, OFSCs were fermented by B. subtilis and Lactobacillus plantarum, and the effects of the resulting products on nitric oxide (NO) production, cytokine secretion, NF-κB activity, and MAPK phosphorylation in RAW 264.7 macrophages were investigated.
Materials and Methods
Sample preparation
OFSCs were purchased from a local company and washed, sliced, and freeze-dried. B. subtilis was isolated from Doenjang (a fermented soybean paste) purchased from Doguri Agriculture Association Corporation (Jeju, Korea), and L. plantarum (KCCM 80077) was isolated from Kimchi (a traditional Korean food) purchased from Jejuinjeju Agriculture Association Corporation (Jeju, Korea). They were identified based on the 16S ribosomal RNA gene sequencing analysis by Sol-Gent Co. (Seoul, Korea), and the sequence similarities were compared with the NCBI database (
L. plantarum and B. subtilis were initially incubated in de Man–Rogosa–Sharpe broth (Gibco, Rockville, MD, USA) and lactose broth (Gibco), respectively, in conical 50-mL tubes for 24 h, and then inoculated into 500-mL flasks containing 200 mL of basal medium. The basal medium consisted of 200 g of dried OFSCs, 20 g of yeast extract, and 2 g of anhydrous dextrose. The cultures were incubated in a rotary shaker (180 × g ; SI-600R, JeioTech, Seoul, Korea) at 35°C for 4 days.
The cultures (50 mL) were extracted three times using 80% methanol (500 mL) (HPLC grade; Fisher Scientific Co., Chicago, IL, USA) at room temperature for 24 h and filtered through No. 5A filter paper (Whatman, Maidstone, United Kingdom). The methanol extracts were concentrated in a rotary vacuum evaporator and were stored at −20°C until needed. The immunological activities of the methanol extract of L. plantarum culture medium (LPCME) and B. subtilis culture medium (BSCME) were analyzed.
Cell culture
Murine macrophage RAW 264.7 cells (ATCC#TIB-71) were provided by the American Type Culture Collection (Manassas, VA, USA). The cells were cultivated in Dulbecco's modified Eagle's medium (DMEM) supplemented with 100 μg/mL streptomycin, 100 U/mL penicillin, and 10% heat-inactivated fetal bovine serum, and maintained at 37°C in a humidified atmosphere containing 5% CO2.
Cell viability measurement
Cell viability was measured using an EZ-Cytox Cell Viability Assay Kit (Daeil Laboratory, Seoul, Korea). Cells were cultured in 96-well plates at 4 × 105 cells/well for 24 h and subsequently treated with various concentrations of the samples for a further 24 h. Then, 10 μL of the kit solution was added to each well, followed by incubation for 2 h at 37°C in 5% CO2. Cell viability was determined by measuring formazan production by enzyme-linked immunosorbent assay (ELISA) at a wavelength of 480 nm. 18
Measurement of NO production
RAW 264.7 cells were cultured in DMEM in 96-well plates, pretreated with various concentrations of the samples for 1 h, and stimulated with lipopolysaccharide (LPS) for 24 h. The cultured medium was mixed with Griess reagent (Sigma, St. Louis, MO, USA), and the absorbance was determined at 540 nm using a microplate reader. 19
Reverse transcription–polymerase chain reaction analysis
LPS-induced RAW 264.7 cells were incubated in the presence of various concentrations of the samples for 6 h, and total cellular RNA was isolated using the TRIzol reagent (Molecular Research Center, Cincinnati, OH, USA). 20 One microgram of total cellular RNA was reverse transcribed from each sample using the ImProm-II™ Reverse Transcription System (Promega, Madison, WI, USA). Polymerase chain reaction (PCR) was then performed on aliquots of the cDNA preparation to amplify TNF-α, interleukin-1β (IL-1β), interleukin-6 (IL-6), and β-actin mRNA (Table 1). 21 The reactions were conducted in a 20-μL volume containing Taq DNA polymerase, dNTPs (0.2 mM), reaction buffer, and primers (Bioneer, Seoul, Korea). Relative band density was measured by densitometry using appropriate image acquisition and analysis software (VisionWorks, UVP, Cambridge, United Kingdom). 22
IL-6, interleukin-6; IL-1β, interleukin-1β; TNF-α, tumor necrosis factor-α.
Western blot analysis
Western bolt analysis was conducted by lysing cells in radioimmunoprecipitation assay buffer (Upstate Biotechnology, Charlottesville, VA, USA) containing a protease inhibitor mixture. The levels of protein were measured using a Bio-Rad Protein Assay Reagent (Bio-Rad, Hercules, CA, USA). Proteins were subjected to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The separated proteins were transferred to polyvinylidene fluoride membranes (Millipore Corporation, Bedford, MA, USA). The membrane was blocked with 5 g/100 mL skim milk dissolved in tris-buffered saline with Tween 20 buffer (10 mM Tris-HCl [pH 7.5], 0.1% [v/v] Tween 20, 150 mM NaCl) and sequentially incubated with phosphor-c-JUN NH2-terminal protein kinase (JNK) (Thr183/Tyr185), phosphor-P38 MAPK (1:1000; Cell Signaling Technology, Danvers, MA, USA), phosphor-extracellular signal-regulated kinase (ERK) (1:1000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), and secondary antibodies (1:1000; Jackson immunoResearch, West Grove, PA, USA) followed by the WEST-ZOL Western Blot Detection System (iNtRON Biotechnology, Gyeonggi, Korea). Relative band density was measured by densitometry as described previously. 22
Cytokine assay
Cytokine (TNF-α, IL-1β, and IL-6) levels were quantified by ELISA according to the protocol recommended by the manufacturer (Quantikine, R&D Systems, Minneapolis, MN, USA). 23
Transient transfection and luciferase assay
Cells were cotransfected with 0.4 mg of the Renilla luciferase reporter plasmid pGL4.74[hRluc/TK] (Promega) and 1 mg of the NF-κB-promoted luciferase reporter gene plasmid pGL4.32[luc2P/NF-κB-RE/Hygro] (Promega) using FuGENE 6 Transfection Reagent (Roche, Indianapolis, IN, USA). The cells were incubated with LPS (100 ng/mL) for 24 h in the presence or absence of the samples, and the luciferase activity in the cell lysate was determined using a Dual-Luciferase Reporter Assay Kit (Promega). Luciferase activity was normalized to the transfection efficiency using the Renilla luciferase reporter plasmid expression vector. Induced luciferase activity was measured relative to that of nonstimulated cells. 24
Statistical analysis
Data are expressed as means ± standard deviation of three replications. The statistical differences between experimental data and those of the respective controls were evaluated using Student's t-test (SPSS, Inc., Chicago, IL, USA). The probability (P < .05) was considered statistically significant.
Results
NO production in RAW 264.7 cells
NO production in LPS-induced (Fig. 1) and IFN-γ-induced (Fig. 2) RAW 264.7 cells treated with LPCME and BSCME was investigated. LPCME and BSCME at 1000 μg/mL did not exhibit cytotoxicity in vitro. Therefore, NO production in LPS- and IFN-γ-stimulated RAW 264.7 cells was evaluated after treatment with the samples at 62.5–1000 μg/mL.
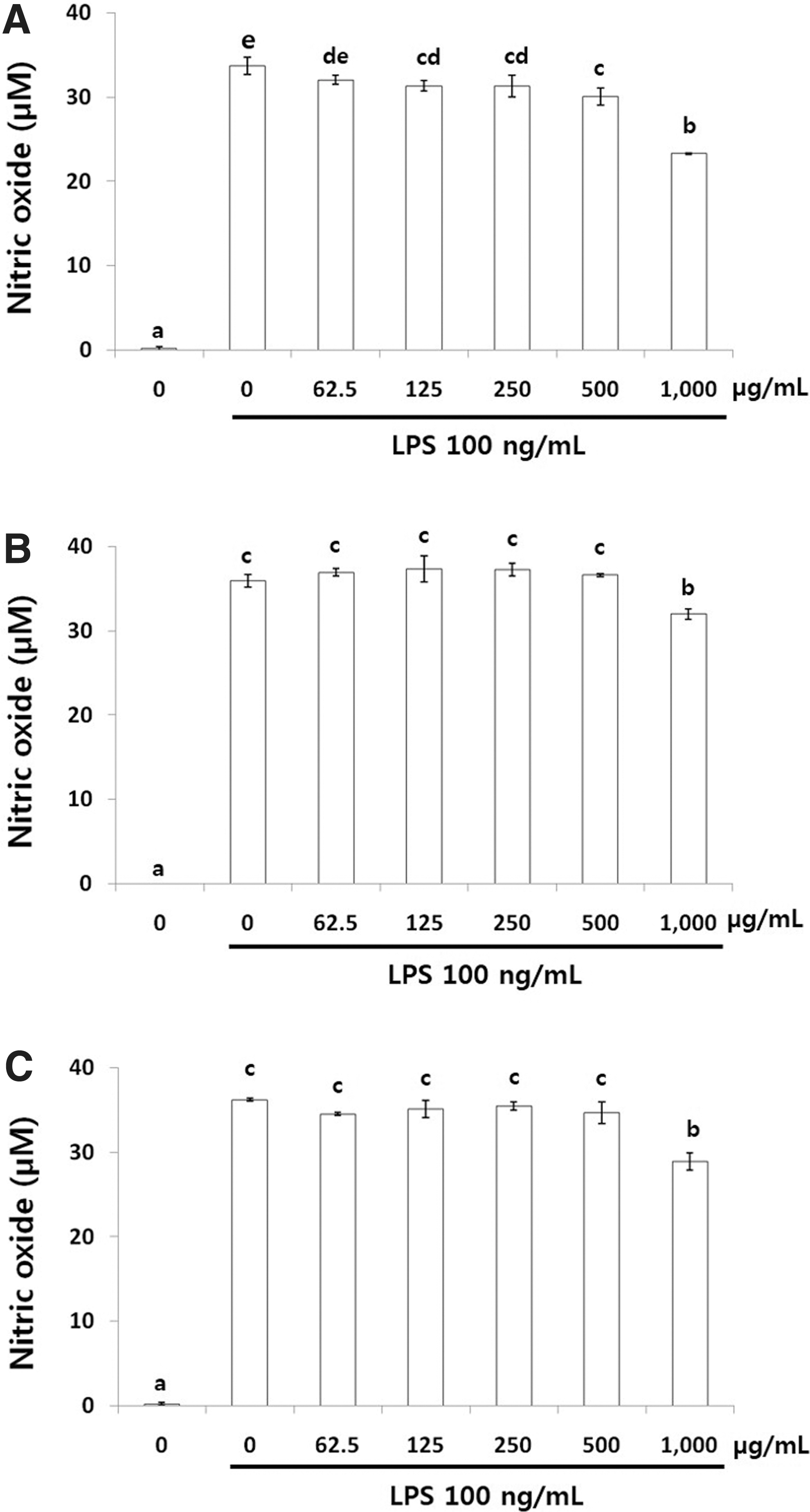
NO production in LPS (100 ng/mL)-induced RAW 264.7 cells treated with 80% methanol extracts of OFS cladodes.
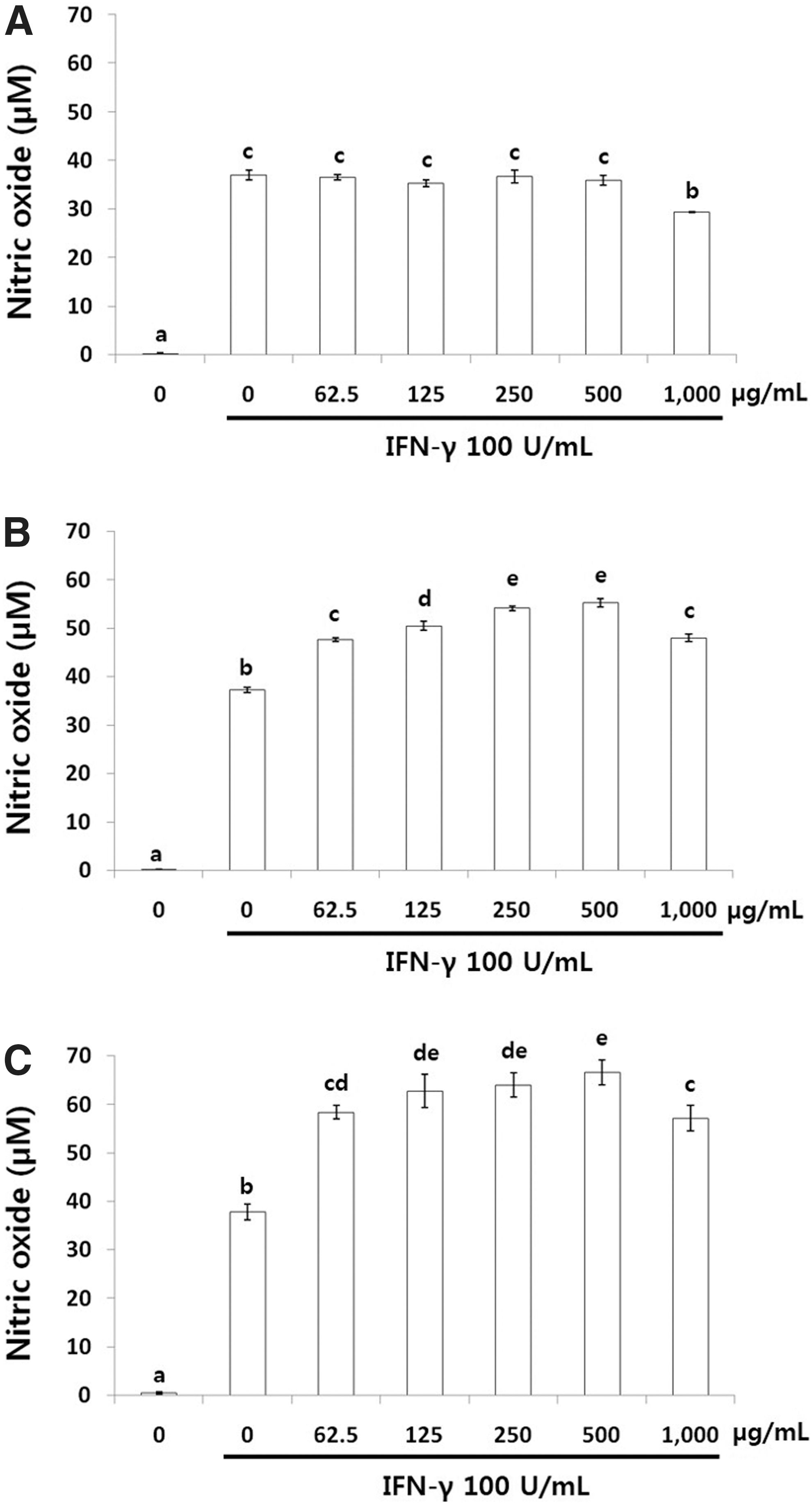
NO production in IFN-γ (100 U/mL)-induced RAW 264.7 cells treated with 80% methanol extracts of OFS cladodes.
Nonfermented OFSCs (Fig. 1A) inhibited NO production slightly, whereas LPCME (Fig. 1B) and BSCME (Fig. 1C) <1000 μg/mL did not inhibit NO production at all in LPS-stimulated RAW 264.7 cells. However, LPCME (Fig. 2B) and BSCME (Fig. 2C) increased IFN-γ-stimulated NO production dose dependently in RAW 264.7 cells. At 500 μg/mL, LPCME and BSCME increased NO production by 55.2 and 66.5 μM, respectively.
NO production by RAW 264.7 cells not treated with LPS and IFN-γ and cultured in the presence of LPCME or BSCME for 24 h was measured (Fig. 3). LPCME did not affect NO production (Fig. 3A), but BSCME at less than 500 μg/mL increased NO production (Fig. 3B) significantly in a dose-dependent manner. Therefore, BSCME was considered to have immunomodulatory activity similar to LPS, and further experiments were performed using only BSCME.
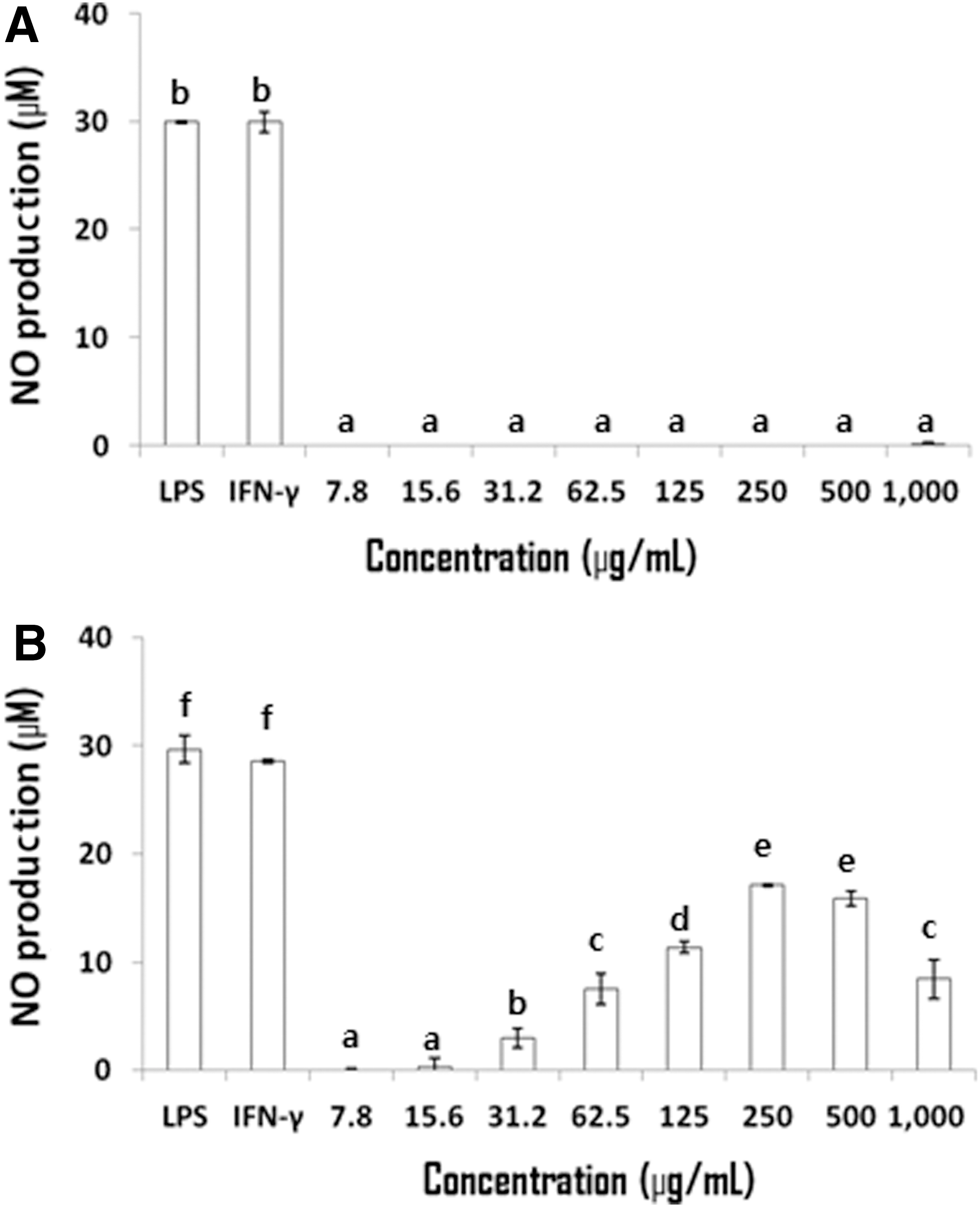
NO production in RAW 264.7 cells treated only with 80% methanol extracts of OFS cladodes. Fermented with
Effects on TNF-α, IL-1β, and IL-6 mRNA and protein levels in RAW 264.7 cells
The effects of BSCME on TNF-α, IL-1β, and IL-6 mRNA levels in RAW 264.7 cells were determined next (Fig. 4). Treatment with BSCME for 6 h increased the TNF-α, IL-1β, and IL-6 mRNA levels in RAW 264.7 cells dose dependently. When compared with β-actin at 500 μg/mL as a control, BSCME increased TNF-α and IL-1β mRNA levels by 83.8% and 82.2%, respectively.
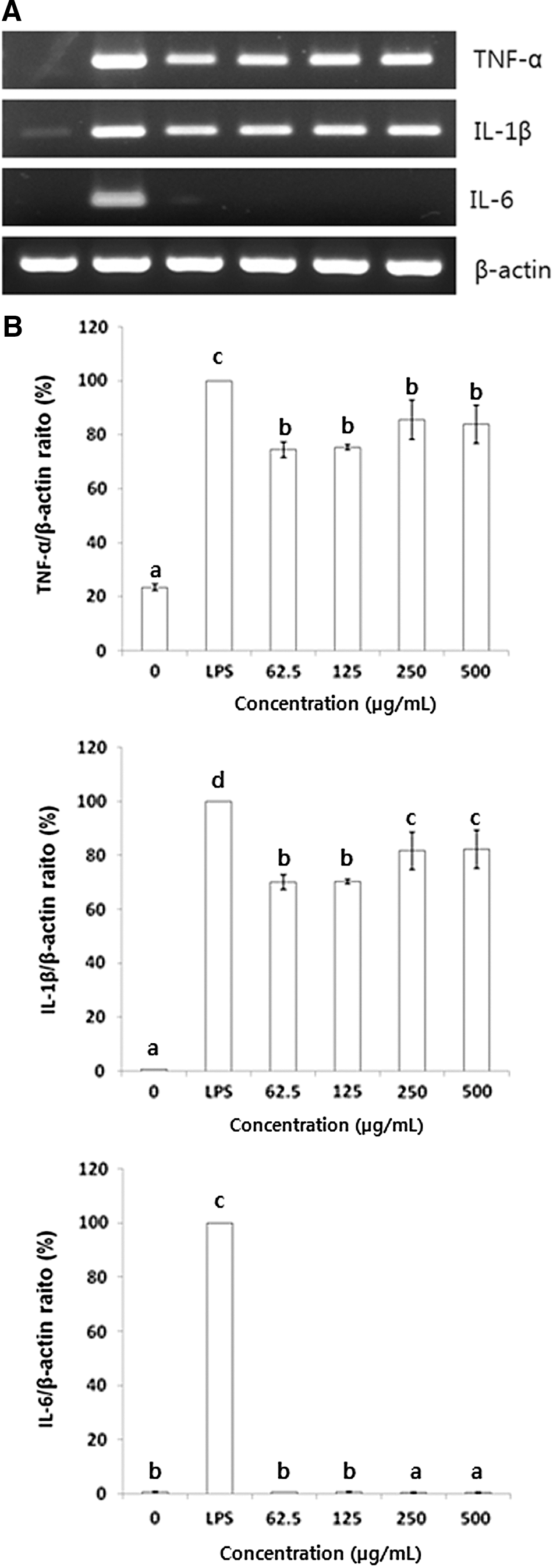
Effect of an 80% methanol extract of OFS cladodes fermented with B. subtilis on TNF-α, IL-1β, and IL-6 mRNA levels in RAW 264.7 cells.
The effects of BSCME on TNF-α, IL-1β, and IL-6 protein levels in RAW 264.7 cells after 6 and 24 h (Fig. 5) were also assessed. TNF-α and IL-1β protein levels in RAW 264.7 macrophages were increased after 24 h (Fig. 5A, B), and the IL-6 protein level was slightly increased at the same time point (Fig. 5C).
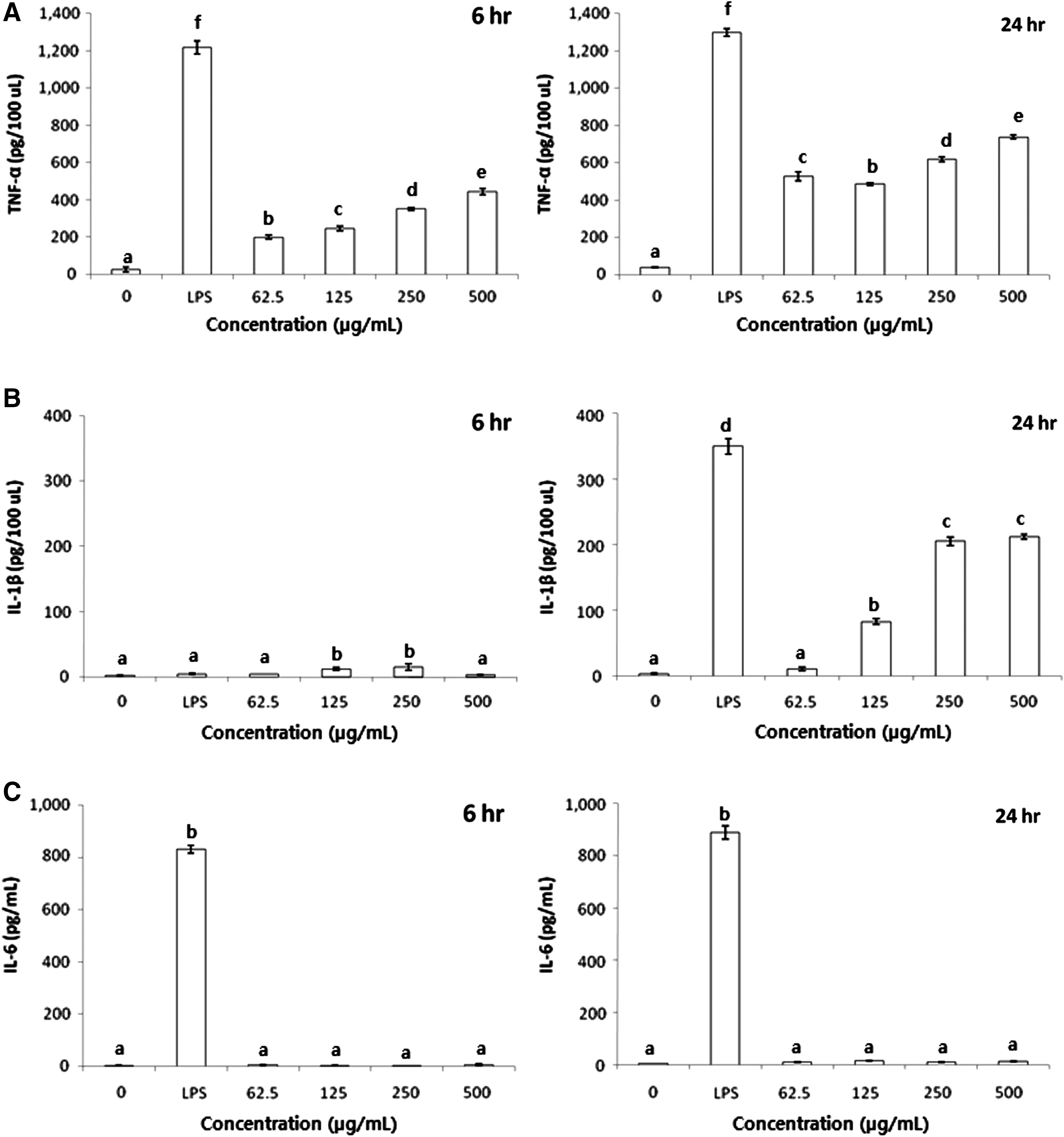
Effect of 80% methanol extracts of OFS cladodes fermented with B. subtilis on
Induction of NF-κB activity in RAW 264.7 cells
To determine whether IL-1β mRNA and protein levels were induced by NF-κB activity in RAW 264.7 cells, we investigated the effect of BSCME on NF-κB activity using a pNF-κB-Luc plasmid by inserting four NF-κB binding sites into the pLuc-promoter vector (Fig. 6). NF-κB-dependent luciferase activities were increased significantly in a dose-dependent manner by BSCME treatment; indeed, 250 μg/mL BSCME increased the activity 9.1-fold compared with that of the control.
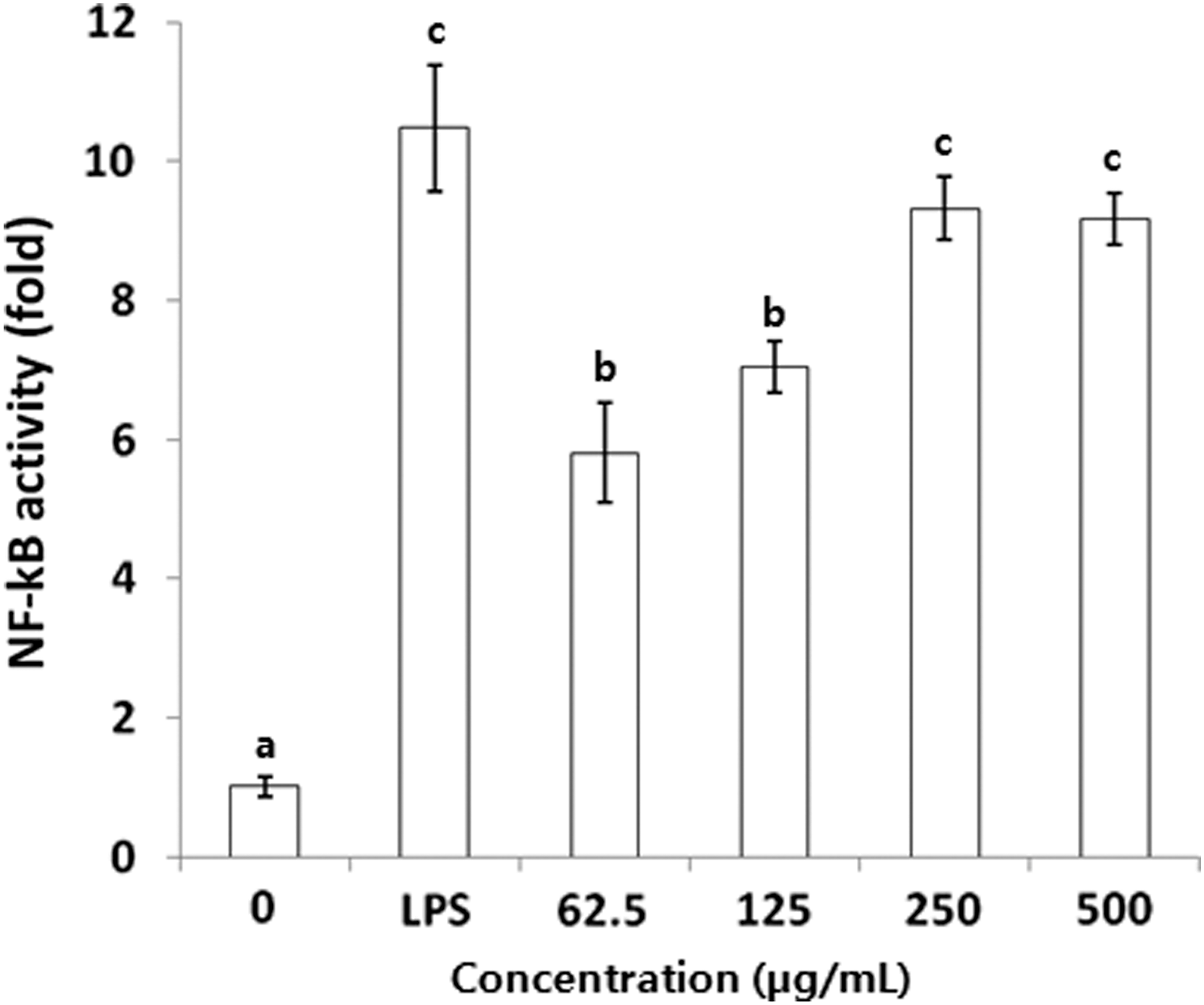
Effect of 80% methanol extracts of OFS cladodes fermented with B. subtilis on transcriptional activation of NF-κB in RAW 264.7 cells. The values followed by the same letter are not different significantly at P < .05. NF-κB, nuclear factor-κB.
Activation of p38, JNK1/2, and ERK1/2 MAPKs in LPS-activated RAW 264.7 cells
To determine whether the increase in NO, TNF-α, and IL-1β production because of BSCME was mediated through the MAPK pathway, the effects of BSCME on the phosphorylation of MAPKs in RAW 264.7 macrophages were measured by Western blot analysis (Fig. 7). BSCME induced the phosphorylation of p38, JNK, and ERK dose dependently, but did not affect total ERK levels. Compared with 500 μg/mL of LPS as the control, BSCME increased p38, JNK, and ERK phosphorylation by 379%, 244%, and 104%, respectively. Further studies are needed to confirm the effect of BSCME on MAPK: ERK, p38, and JNK by using specific blockers of each pathway.
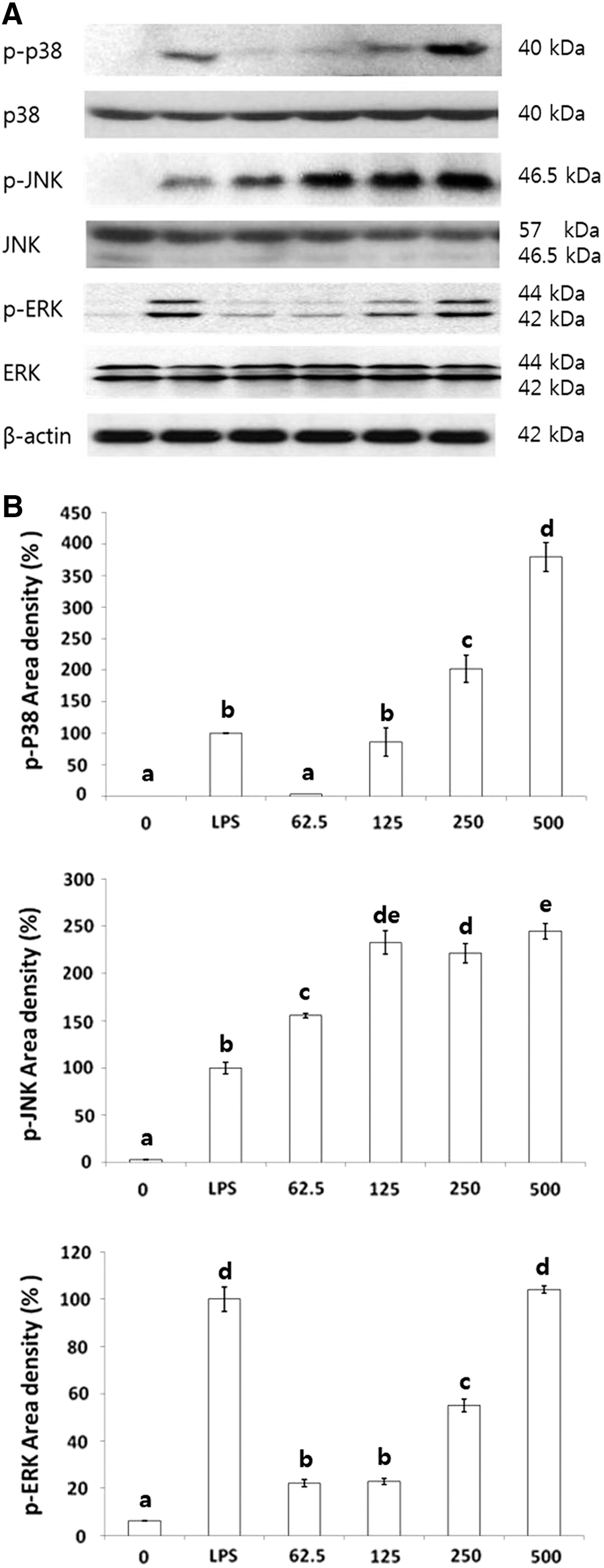
Effect of 80% methanol extracts of OFS cladodes fermented with B. subtilis on MAPK protein levels in RAW 264.7 cells.
Discussion
Macrophages and lymphocytes play major roles in nonspecific (innate immunity) and specific (adaptive immunity) defense systems. Macrophages are the first cells to recognize infectious agents and are central to cell-mediated and humoral immune responses. Activated macrophages exhibit increased production of NO and cytokines, which function as immune mediators. 25 NO is one of the primary effector molecules of activated macrophages. Maximal activation of macrophages in vitro requires at least two signals, for example, LPS and IFN-γ. 26
In this study, cultured mouse macrophages were treated with LPCME and BSCME, and NO production and cytokine secretion were evaluated. LPCME and BSCME did not affect LPS-induced NO production (Fig. 1B, C). However, LPCME (Fig. 2B) and BSCME (Fig. 2C) did increase IFN-γ-induced NO production by RAW 264.7 cells in a dose-dependent manner. Interestingly, treatment with BSCME (Fig. 3B), but not LPCME (Fig. 3A), increased NO production in RAW 264.7 cells not treated with LPS and IFN-γ. Treatment with only LPCME (Fig. 3A) did not affect NO production, whereas treatment with combined IFN-γ and LPCME (Fig. 2B) increased NO production in RAW 264.7 cells because of the synergistic effect. This suggests that BSCME is capable of activating RAW 264.7 macrophages.
Chan and Riches 27 reported that RAW 264.7 cells stimulated with IFN-γ (10 U/mL) or LPS (1 ng/mL) alone did not produce NO. In contrast, there was marked synergistic NO production when cells were costimulated with IFN-γ and LPS.
LPS is the major etiologic component of Gram-negative bacteria. Unlike Gram-negative bacteria, Gram-positive bacteria lack LPS and instead contain peptidoglycan (PGN) and lipoteichoic acid (LTA) in their cell walls. 28 PGN and LTA induce NO production by RAW 264.7 cells. 29 As Gram-positive bacteria, L. plantarum and B. subtilis contain PGN and LTA. Therefore, OFSC fermented by L. plantarum and B. subtilis functions in a manner similar to IFN-γ to activate RAW 264.7 cells.
Flavonoids and aglycone derivatives in saboten cladodes are known to exert immune modulatory effects in in vitro and in vivo assays.
30,31
We analyzed flavonoid compositions in nonfermented and fermented samples using HPLC. Isoquercetin and other unknown compounds were identified in LPCME and BSCME. The content of isoquercetin in the nonfermented sample was higher than those in fermented samples. Kaemferol, isorhamnetin, and quercetin, known as major flavonoid derivatives in saboten cladodes, were not detected in our samples. Therefore, we assumed that PGN and LTA were major active compounds that might be associated with immunostimulatory activity in LPCME and BSCME. Further work should be done to determine amino acid compositions, different chain lengths of the phosphoglycerol backbone, and different degrees of the substitution (the ratio of
The major innate cellular defense mechanisms are phagocytosis and the killing of bacteria and fungi by neutrophilic granulocytes, monocytes, and macrophages, 32 and lysis of viral-infected cells by NK cells. 33 TNF is essential for host defense, as demonstrated by the infectious complications suffered by patients treated with anti-TNF biological agents. 34 TNF and IFN-γ, together with proinflammatory cytokines of the IL-1 family (notably IL-1β and IL-18), also play important roles in host defense against antimicrobial infection. IL-6 was originally identified as a T cell-derived factor that induced the final maturation of B cells to antibody-forming plasma cells. 35 Our data showed that BSCME increased mRNA (Fig. 4) and protein (Fig. 5) levels of TNF-α and IL-1β in RAW 264.7 cells. However, BSCME did not affect IL-6 mRNA (Fig. 4B) and protein (Fig. 5C) levels.
PGN and LTA induce the release of TNF-α, IL-1β, and IL-6 from cultured macrophage monocytes. 36 In a previous study, LTA from Staphylococcus aureus did not induce cytokine release from monocytes. 28 After stimulation of whole blood with PGN or LTA, highly pure populations of monocytes (CD14 positive), T cells (CD2 positive), B cells (CD19 positive), and granulocytes (CD15 positive) were analyzed by reverse transcription–PCR and reported as their TNF-α, IL-6, and IL-10 mRNA levels. 36 PGN stimulation enhanced IL-6 mRNA levels in CD14- and CD2-positive cells but not in CD19- and CD15-positive cells. The addition of LTA resulted in a marked increase in IL-6 mRNA levels in CD14-positive cells, and lesser increases in CD2- and CD15-positive cells. However, in our study, BSCME did not induce IL-6 mRNA expression and production. Further work should be done to determine the effect of BSCME on IL-6 production.
Several signaling pathways, including the MAPK pathway, have been proposed to be involved in the response to LPS stimulation. LPS activates three MAPKs (p38, JNK1/2, and ERK1/2) in mouse macrophages and human monocytes. 37 In stimulated macrophages, MAPK phosphorylation is a prerequisite for NO and cytokine production. 38 Activation of NF-κB has been reported to be triggered by MAPKs. 18,39 During infection with Gram-positive bacteria, LTA is recognized by Toll-like receptor 2 (TLR2). 40 The resulting TLR2 activation activates intracellular messengers (e.g., myeloid differentiation factor 88, TNF receptor-associated factor 6, and MAPK), leading to the activation of transcription factors, including NF-κB, which is required for the expression of inflammatory cytokines. 41 In our study, BSCME increased NF-κB transcriptional activity and MAPK activation in RAW 264.7 macrophages. NF-κB transcriptional activity (Fig. 6) and the phosphorylation levels of MAPKs (p-p38, p-JNK, and p-ERK; Fig. 7) were also significantly increased by BSCME treatment. These findings suggest that BSCME enhanced TNF-α and IL-1β production by inducing MAPK phosphorylation and NF-κB transcriptional activity.
In conclusion, BSCME exerted an immunostimulatory effect, which was mediated by MAPK phosphorylation and NF-κB activation. This led to elevated TNF-α and IL-1β gene expression in RAW 264.7 macrophages. Therefore, BSCMs show promise for use as an immunostimulatory therapeutic agent.
Footnotes
Acknowledgments
This research was supported by the Ministry of Trade, Industry and Energy (MOTIE) and Korea Institute for Advancement of Technology (KIAT) through the Research and Development for Regional Industry.
Author Disclosure Statement
No competing financial interests exist.