
Abstract
Several reports indicate anti-hyperglycemic effects of Syzygium aromaticum. In the present study, we report for the first time that clove extract (SAM) and its compound nigricin (NGC) decreases free fatty acid-mediated insulin resistance in mouse myoblasts. In addition, NGC was able to diminish insulin resistance in a diabetic mouse model. We observed that SAM and its compound NGC exhibited significant antioxidant activity in murine skeletal muscle cells. They also modulated stress signaling by reducing p38 MAP kinase phosphorylation. NGC and SAM treatments enhanced proximal insulin signaling by decreasing serine phosphorylation of insulin receptor substrate-1 (IRS-1) and increasing its tyrosine phosphorylation. SAM and NGC treatments also modified distal insulin signaling by enhancing protein kinase B (PKB) and glycogen synthase kinase-3-beta (GSK-3 beta) phosphorylation in muscle cells. Glucose uptake was enhanced in muscle cells after treatment with SAM and NGC. We observed increased glucose tolerance, glucose-stimulated insulin secretion, decreased insulin resistance, and increased beta cell function in diabetic mice treated with NGC. The results of our study demonstrate that clove extract and its active agent NGC can be potential therapeutic agents for alleviating insulin resistance.
Introduction
I
Improval of daily nutrition is one of the important approaches to avoiding metabolic diseases. 6 The relative safety of edible plants compared with synthetic drugs suggests their significance in preventive medicine. A major advantage is that they can be ingested daily. 7 Studies have shown that food plant extracts have been evaluated for their antidiabetic effects. 8 However, the molecular mechanisms of action of many of the antidiabetic food plants are unknown. Syzygium aromaticum, commonly known as clove, is native to Asia, but it is cultivated in several other parts of the world. 9 Clove is a very valuable spice and has been used for medicinal purposes, such as lowering the blood glucose levels 10 for centuries. Animal studies also support antidiabetic effects of clove. 11
Our group has recently isolated and characterized seven compounds from S. aromaticum. 12 We have also reported the antifungal and antibacterial potential of clove. 13 The mechanisms by which clove reduces the symptoms of diabetes are not clear. Our limited understanding of the molecular basis of the antidiabetic effects of clove limits its use in standard medical care. In the present investigation, we aimed at deciphering the role of clove and its active compounds in upregulating insulin signaling and insulin action in myoblasts and in increasing insulin sensitivity in diabetic mice.
Materials and Methods
Preparation of the plant extract
The flower buds of S. aromaticum (clove) were purchased from the Karachi region and identified by senior taxonomist Dr. Jan Alam in the Department of Botany, University of Karachi, and Karachi, Pakistan. A voucher specimen (KUH-GH No. 01) has been deposited in the herbarium. The clove methanol extract was prepared as previously described. 12 Briefly, the flower buds (5 kg) were powdered and repeatedly extracted with methanol (five times) at room temperature. The solvent was removed under reduced pressure by using a rotary evaporator to obtain a syrupy residue (clove extract, SAM; 554 gm).
Isolation of the compounds from S. aromaticum
Compounds from S. aromaticum were isolated as previously described. 12 Briefly, oleanolic acid lactone (OAL; 10 mg), nigricin (NGC; 18.6 mg) oleanolic acid (45.5 mg), and 2α-hydroxyoleanolic acid (HOA; 14 mg) were obtained by using successive chromatographic techniques, including vacuum liquid chromatography, column chromatography, and preparative thin-layer chromatography. Semi-synthetic compound oleanolic acid acetate (OAA) was derived from oleanolic acid by acetylation with acetic anhydride and pyridine. 12
Cell lines, reagents, and antibodies
C2C12 cells were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). The cell culture media and supplements were purchased from Gibco (Sigma, St. Louis, MO, USA); fetal calf serum (South American origin) was purchased from Biowest; the antibody against phospho serine 307 of insulin receptor substrate-1 (IRS-1) protein was purchased from Millipore; and the antibodies against total IRS-1 protein, phospho tyrosine (p)100, phospho serine 473 of protein kinase B (PKB), the total PKB protein, phospho serine 9 of glycogen synthase kinase-3-beta (GSK-3 beta), total GSK-3 beta protein, phospho-p38 MAPK, and p38 MAPK were from Cell Signaling Technology (Beverly, MA, USA). Palmitic acid and insulin were purchased from Sigma, and glucose-6-phosphate dehydrogenase (G6PDH) was purchased from MP Biomedicals (France).
Cell culture and western blot analysis
C2C12 (mouse skeletal muscle) cells were cultured in Dulbecco's modified Eagle's medium containing 25 mM glucose, 10% fetal calf serum, 2 mM glutamine, 100 units/mL penicillin, and 100 μg/mL streptomycin. A stock solution of palmitic acid was prepared by making a complex of palmitate with fatty acid-free bovine serum albumin (BSA) as previously described. 14 Briefly, palmitic acid was dissolved in 0.1 N NaOH and then diluted in BSA (12% solution) at 37°C to make 10 mM stock solution. The cells were lysed with lysis buffer (50 mM Tris [pH 7.6], 150 mM NaCl, 1% Triton X-100, protease inhibitors, and phosphatase inhibitors). Fifty micrograms of the total protein was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) (7.5%) co-immunoprecipitations, and western blot analysis was performed as previously described. 15
Glucose uptake assay
Myoblasts were cultured in Dulbecco's modified Eagle's medium with 10% fetal calf serum, 100 units/mL penicillin, 100 μg/mL streptomycin, 25 mM glucose, and 2 mM glutamine. Myoblasts were stimulated with palmitic acid, followed by treatment with clove extract and its compounds for 24 h. After stimulation with insulin, muscle cells were incubated with 1 mM 2-deoxy-
Reactive oxygen species levels using dihydroethidium
Dihydroethidium was used to detect intracellular superoxide levels. Muscle cells were stimulated with palmitic acid and/or clove extract and its compounds for 24 h, followed by incubation with dihydroethidium. Fluorescent microscopy was used to record fluorescence, as previously described. 17
Experimental design for animal experiments
Female albino BALB/c mice were housed in controlled conditions with a 12 h light cycle, a temperature of 20°C ± 2°C, and a relative humidity of 60% ± 10%, with free access to food and water. All animal experiments were approved by the Animal Care and Use Committee of the International Centre for Chemical and Biological Sciences (Protocol No. 2014-0015) and were conducted in accordance with the Guide for the Care and Use of Animals published by the National Institutes of Health. The insulin-resistant mouse model was developed by feeding mice a high-fat diet (HFD, ETPF1916; Mucedola) and administering a low dose (50 mg/kg) of streptozotocin (STZ). The mice were designated as high-fat-diet- and streptozotocin-induced diabetic mice (HFD-STZ-DM). 18 Briefly, the mice were randomly assigned into two groups according to their diet for 4 weeks (n = 10–12): normal chow diet AIN-93G or HFD. The HFD mice were then intraperitoneally injected with a low dose (50 mg/kg) of streptozotocin for 3 days (HFD-STZ). Seven days after the STZ injection, the mice with fasting blood glucose levels ≥200 mg/dL were considered diabetic. The diabetic mice were continued on the HFD throughout the study. The diabetic mice were randomly divided into three additional groups, according to the treatment administered by oral gavage for 21 days (n = 10–12): a low dose [HFD-STZ-NGC +5 mg/(kg·day)] or a high dose [HFD-STZ-NGC +10 mg/(kg·day)] of NGC or 100 mg/(kg·day) metformin (HFD-STZ-DM+MET). NGC and metformin were dissolved in 0.5% carboxymethylcellulose (CMC). Normal chow and HFD-STZ mice were administered 0.5% CMC as vehicle by oral gavage.
Glucose tolerance test
The mice were fasted for 14–16 h before the glucose tolerance test. Their blood glucose levels were measured at 0, 15, 30, 90, and 120 min after glucose injection (2 g/kg body weight), as previously described. 19 The plasma glucose levels were measured with a One Touch Ultra glucometer (Life Scan, Inc., USA). The results were expressed as area under the curve (AUC 0–120 min).
Glucose-stimulated insulin secretion
After an overnight fast, the mice were injected with 2 g/kg body weight of glucose. 20 To determine the plasma insulin levels in response to glucose, blood samples were collected from the tail vein at 0, 15, 30, and 60 min after glucose injection. The collected samples were separated by centrifugation at 4°C and stored at −20°C until the assay. The plasma insulin levels were measured by using an enzyme-linked immunosorbent assay (Crystal Chem, USA).
Determination of insulin resistance by homeostatic model of insulin resistance and beta cell function by homeostasis model assessment (HOMA-β)
After an overnight fast, the fasting blood glucose and insulin levels in the mice were used to measure HOMA-IR (homeostatic model of insulin resistance) values according to the following formula: HOMA-IR = [fasting plasma insulin (mU/L) × fasting plasma glucose (mmol/L)/22.5]. 21 HOMA-β scores were determined by using the following formula: HOMA-β = [20 × fasting plasma insulin (mU/L)]/[fasting plasma glucose (mmol/L) −3.5]. 21
Statistical analyses
The sample sizes were determined by using G power software. 22 The results presented in this study are derived from at least three independent experiments. The means ± standard error of the means were calculated, and groups of data were compared by using one-way analysis of variance, followed by Tukey's post hoc test using Sigma Plot software. For all analyses, the level of statistical significance was set at P < .05.
Results
Selection of the active compound
We performed an initial screening of compounds, OAL, OAA, NGC, and HOA, from the methanol extract of clove (SAM). The compounds were evaluated for their influence on p38 MAP kinase activation in C2C12 cells. We observed that NGC suppressed the phosphorylation of p38 MAP kinase comparably to SAM (Fig. 1A). Therefore, NGC was selected to decipher the mechanism of action of S. aromaticum in alleviating insulin resistance. The characterization of the compounds is described in Experimental S1: Characteristics of Nigricin Isolated from Syzygium aromaticum section in Supplementary Data (Supplementary Data are available online at
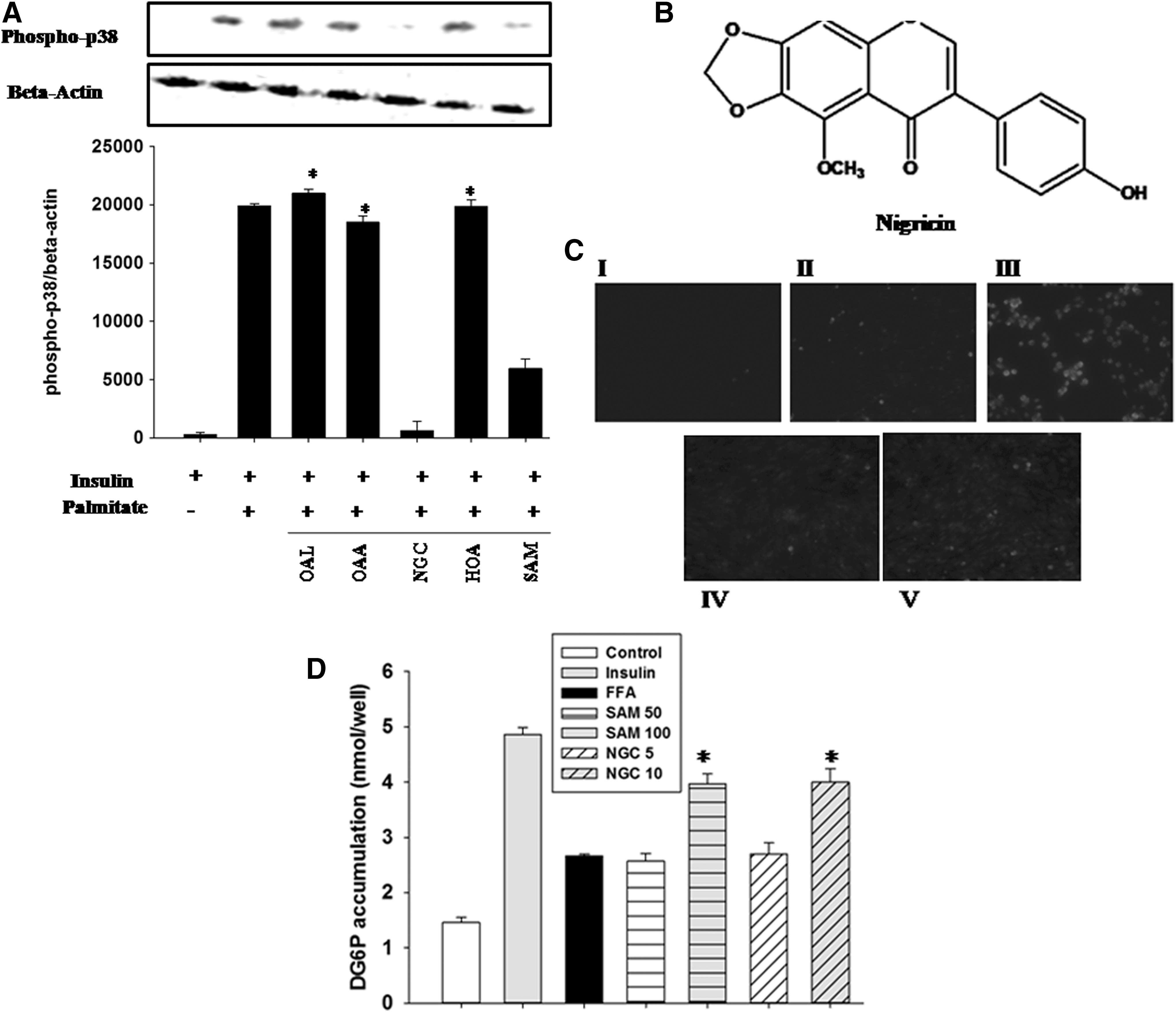
Screening of compounds isolated from Syzygium aromaticum by modulation of stress kinase activity.
Role of SAM and NGC in reactive oxygen species production in muscle
To further validate the antioxidant capacity of SAM and NGC, we evaluated their antioxidant properties by using reactive oxygen species (ROS)-activated fluorescent dye dihydroethidium. We observed that superoxide levels in the FFA-stimulated C2C12 cells were increased compared with the cells stimulated with insulin alone, positive control (Fig. 1C). SAM- and NGC-treated cells exhibited a significant reduction in FFA-induced ROS production.
Determination of the effect of SAM and NGC on glucose uptake in muscle
To assess the potential effect of clove and its active compound on glucose clearance in myotubes, we measured glucose uptake in C2C12. A standard curve was generated by placing standard solutions of 2DG6P in wells of a culture plate without cells. We found that C2C12 cells that had been treated with SAM (100 μg/mL) and NGC (10 μM) were able to restore insulin-mediated effects on glucose uptake (Fig. 1D).
Effect of SAM and NGC on proximal insulin signaling
Agents such as FFAs can trigger serine phosphorylation of IRS-1. 4 C2C12 cells were stimulated with palmitic acid and treated with the plant extract and its active compound to determine the effects of SAM and NGC on lowering FFA-induced serine phosphorylation of IRS-1 in muscle cells. We found that higher doses of SAM and NGC significantly diminished IRS-1 phosphorylation at serine 307 in muscle cells compared with the cells stimulated with FFA alone (Fig. 2A).
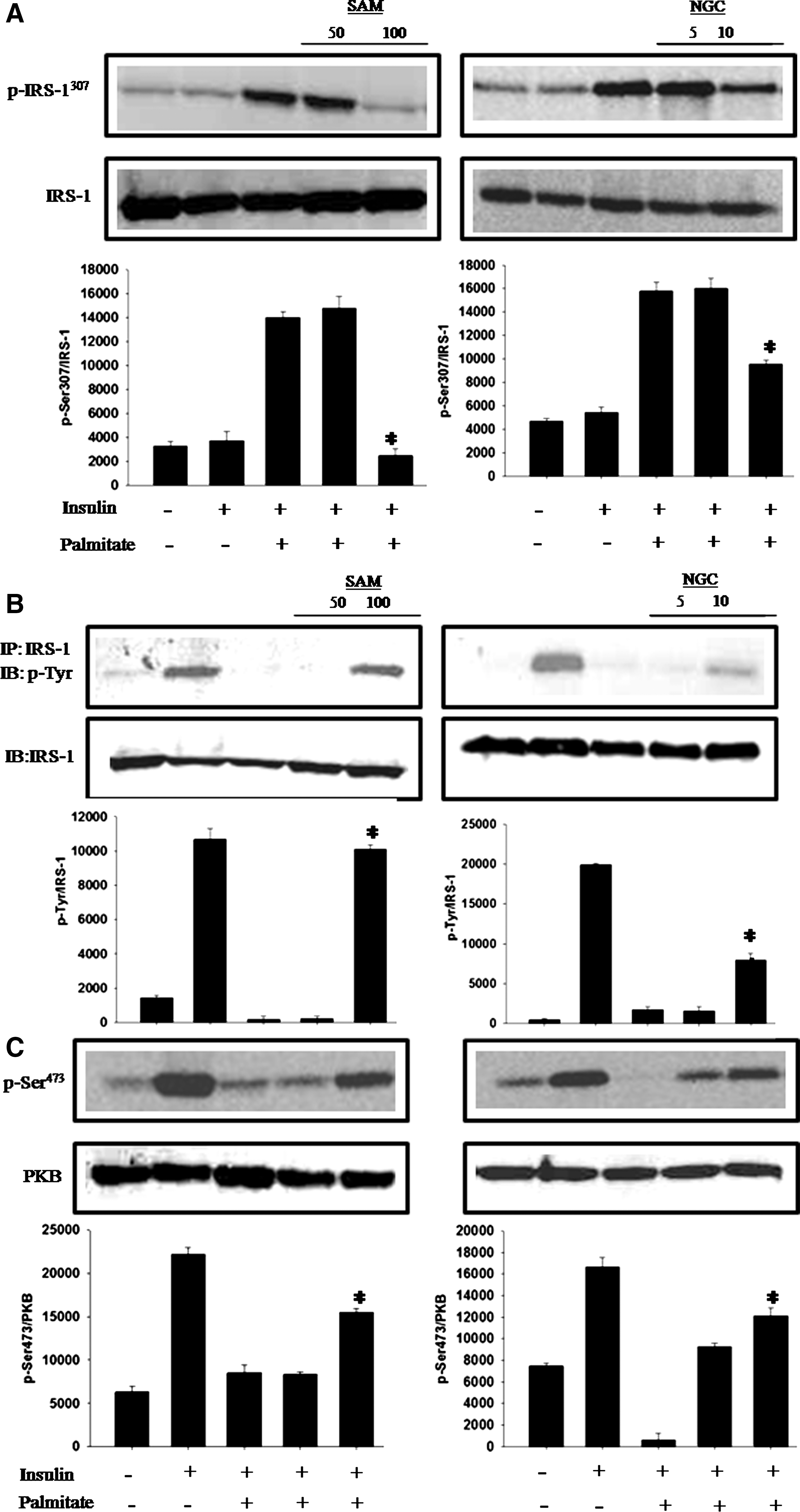
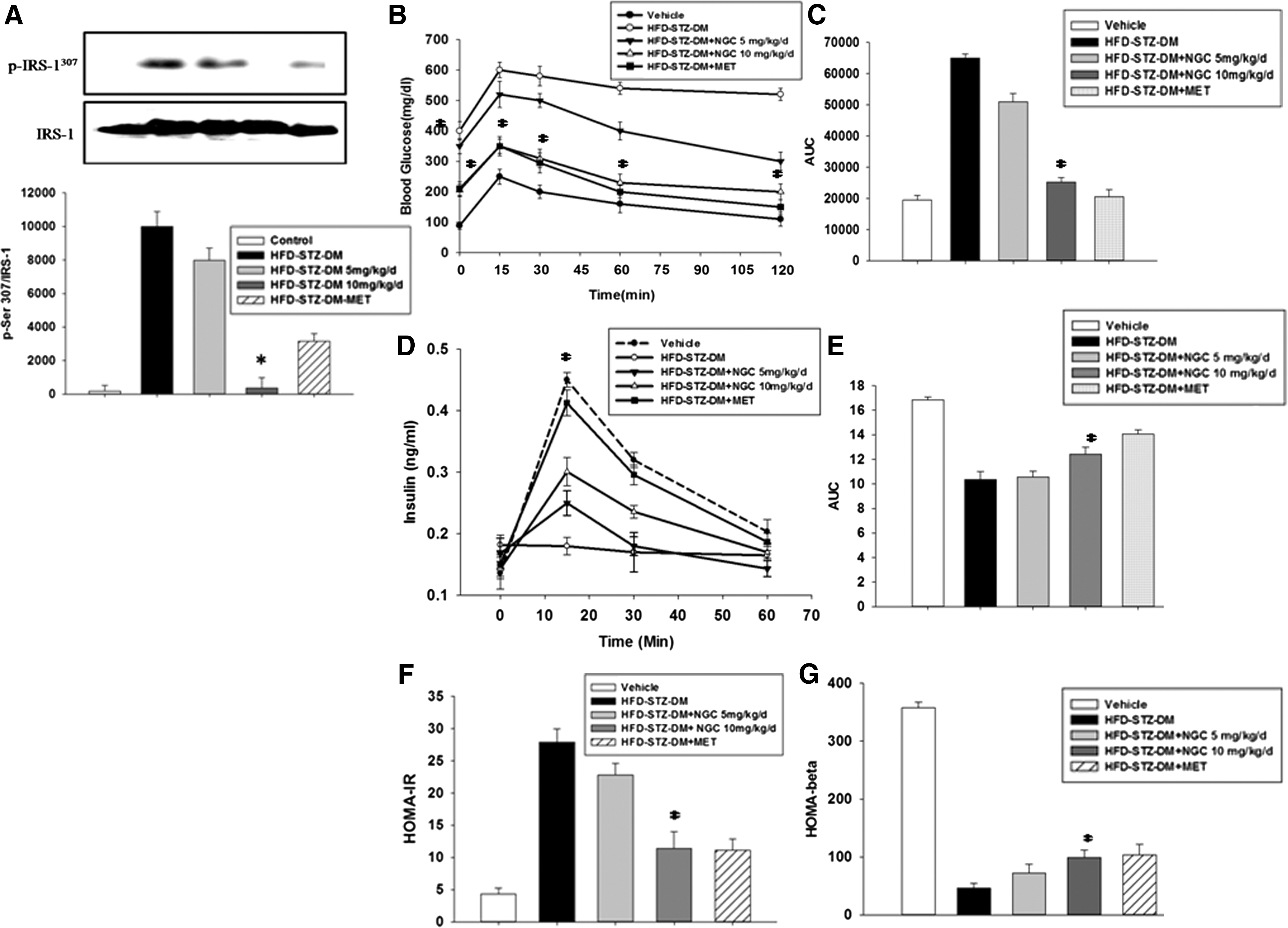
IRS-1 tyrosine phosphorylation has a central role in the upregulation of insulin signal transduction. 23 Muscle cells were treated with SAM and NGC to evaluate their ability to restore FFA-mediated downregulation of tyrosine phosphorylation. We observed significantly enhanced phosphorylation of the immunoprecipitated IRS-1 after SAM and NGC treatment compared with the cells treated with FFA alone (Fig. 2B). In addition, NGC- and metformin-treated mice also exhibited a significant reduction in serine phosphorylation of IRS-1 in skeletal muscle tissue as compared with the HFD-STZ-DM (Fig. 3A), indicating the in vivo effects of NGS on key insulin signaling protein.
Assessment of the effect of SAM and NGC on distal insulin signaling
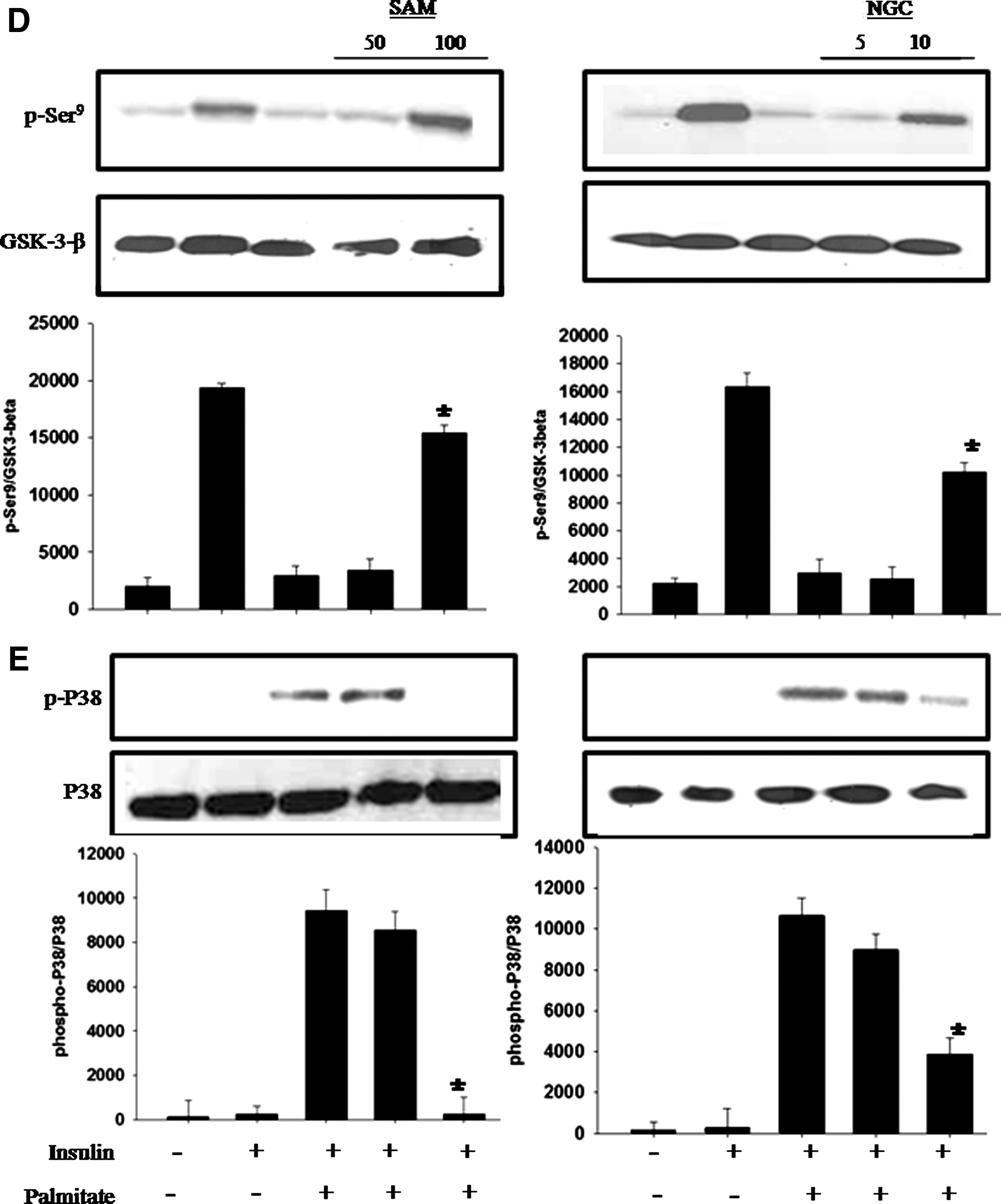
To decipher the mechanism of action of SAM and NGC, we questioned whether they can rescue the FFA-induced decrease in PKB phosphorylation? When C2C12 cells were treated with SAM and NGC and stimulated with insulin, PKB phosphorylation at serine 473 was restored (Fig. 2C). Similarly, we found that muscle cells exposed to FFA alone showed reduced levels of GSK-3 phosphorylation (Fig. 2D), whereas cells that had been treated with higher concentrations of SAM and NGC restored GSK-3 phosphorylation to the levels of the cells treated with insulin alone (positive control).
Determination of mechanisms behind restoration of insulin signal transduction
Because S. aromaticum methanol extract (SAM) and NGC have shown significant antioxidant activity, we hypothesized that they decrease p38 MAPK activity by reducing oxidative stress. We evaluated p38 MAPK activity in response to treatment with a combination of FFA and different concentrations of SAM or NGC. We found that after FFA stimulation, the insulin-stimulated C2C12 cells exhibited increased p38 MAPK phosphorylation at threonine 180 and tyrosine 182, whereas this phosphorylation was significantly diminished after SAM and NGC treatment (Fig. 2E).
Effect of SAM and NGC on glucose tolerance and glucose-stimulated insulin secretion in diabetic mice (HFD-STZ-DM)
NGC- and metformin-treated mice exhibited a significant improvement in glucose tolerance compared with the diabetic mice model without treatment (Fig. 3B). NGC- and metformin-treated mice exhibited reduced blood glucose levels at all time points, that is, 0, 15, 30, 60, and 120 min after glucose loading. The AUC calculation also showed that NGC treatment reduced glucose excursion in a dose-dependent manner that was statistically significant at the dose of 10 mg/(kg·day) (Fig. 3C). In the glucose-stimulated insulin secretion (GSIS) experiments, the insulin levels of the NGC-treated mice were increased after glucose loading compared with the diabetic mice (HFD-STZ-DM). In addition, the insulin levels in the NGC-treated mice were comparable to those of the normal mice (Fig. 3D, E).
Effect of SAM and NGC on insulin resistance and beta cell function in insulin-resistant diabetic mice
Compared with diabetic mice (HFD-STZ-DM), insulin resistance was significantly reduced in the NGC- and metformin-treated mice (Fig. 3F). Pancreatic beta cell function was not affected in mice treated with a low dose of the compound (Fig. 3G). However, the HOMA-β score was slightly increased in mice treated with a high dose of NGC [10 mg/(kg·day)].
Discussion
Previous studies have shown that clove and its active compounds can alleviate diabetes symptoms in animals. 24 S. aromaticum has been reported to increase glucose uptake in liver cells. 10 A recent study pointed out the role of clove extract in enhancing energy metabolism in myocytes. 25 In the current study, we deciphered a few of the mechanisms involved in the alleviation of insulin resistance by clove and its active compounds (Fig. 4). We report for the first time that clove extract (SAM) and its active compound NGC decrease FFA-mediated insulin resistance in mouse myoblasts. Also, NGC was able to diminish insulin resistance in a diabetic mice model, hence proving our hypothesis. In mouse muscle cells, we found that SAM and NGC decreased ROS production. A significant body of evidence has demonstrated that FFAs and oxidative stress mediate insulin resistance by activating stress signaling pathways in muscles, specifically the JNK and p38 MAP kinase pathway. 4,26 Therefore, we tested their ability to inhibit the stress signaling pathways. Interestingly, we found that SAM and NGC were able to reduce p38 MAP kinase phosphorylation at physiological doses. Therefore, we further deciphered the role of clove extract and its active compound in proximal insulin signal transduction. Tyrosine phosphorylation of IRS proteins (IRS-1) enhances insulin signaling, and serine phosphorylation of IRS-1 diminishes insulin signaling. 27 The stimulation of skeletal muscle cells with NGC and clove extract resulted in a reduction in serine phosphorylation and increased tyrosine phosphorylation of IRS-1, indicating that clove extract and NGC improve insulin signaling. We suggest that clove extract and NGC modulated IRS-1 phosphorylation indirectly by deactivating p38 MAPK. 26 While investigating the distal insulin signaling, we observed that treatment of murine muscle cells with clove and its active compound enhanced PKB and GSK-3 phosphorylation, the downstream effectors of insulin signal transduction. 28 These results indicate a possible mechanism for the improved insulin sensitivity mediated by SAM and NGC treatment. One of the hallmarks of insulin resistance is the incompetence of insulin-responsive cells to clear glucose from the blood stream. 29 Most of the insulin-stimulated glucose uptake occurs in skeletal muscle cells. 30 We hypothesized that clove and its compound-mediated upregulation of insulin signaling can be reflected in the enhanced glucose uptake by murine muscle cells. We established that treatment with SAM and NGC was able to restore the insulin effect on glucose uptake in C2C12 cells. We further tested the ability of NGC to alleviate insulin resistance in a mouse model that mirrors the progression of diabetes seen in diabetic patients. At the dose of 10 mg/(kg·day), NGC significantly improved glucose tolerance in mice. Because GSIS and pancreatic beta cell function were enhanced to a lesser extent, we concluded that NGC improved glucose clearance in mice, by increasing insulin sensitivity. This was also reflected in the HOMA-IR results. The limitation of our study is a lack of SAM and NGC function in the liver and pancreas; in addition, clinical studies on clove and its active components are merited. Based on our findings, we conclude that our data validate the use of clove as an antidiabetic spice and support the value of SAM and NGC as potential treatment for diabetes and related diseases.
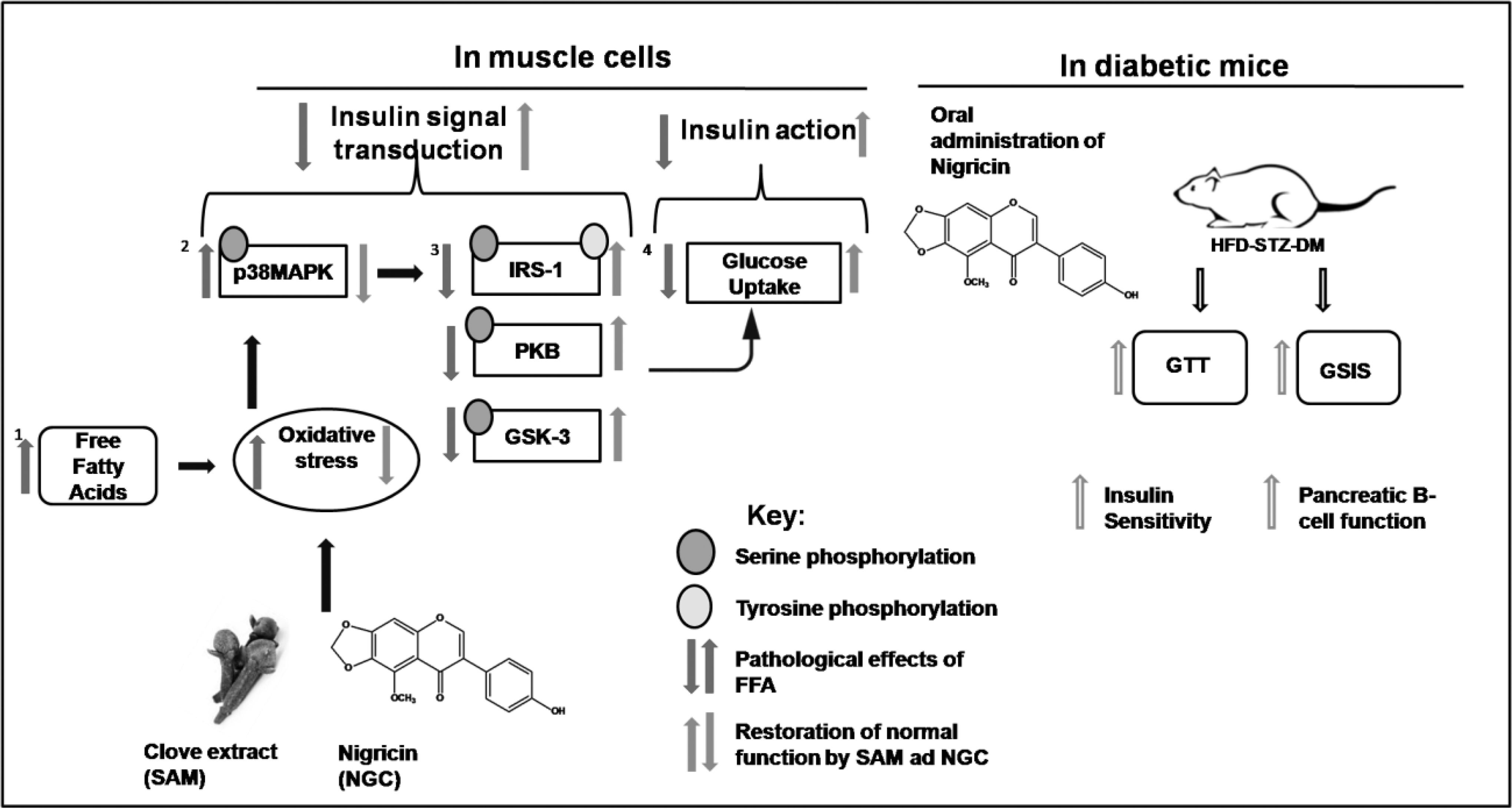
Graphical abstract. Clove extract and its compound nigricin up-regulated FFA mediated diminished insulin signaling and enhanced glucose uptake in muscle. They also enhanced insulin sensitivity in diabetic mice. The graphical abstract is original artwork made in Microsoft PowerPoint 2007.
Footnotes
Acknowledgment
This work was funded by the Higher Education Commission of Pakistan under Grant No. 20-1141 to R.S.W.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.