
Abstract
The transport behavior and absorption mechanism of Ala-His-Leu-Leu (AHLL) intestinal absorption in Caco-2 cell monolayers were clarified systemically. The safe absorptive concentration of AHLL was 200 μg/mL, which was determined by the 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide assay. The permeation of AHLL was concentration dependent in a bidirectional transfer and reached a plateau at 90 min. The efflux ratio was above 0.5, suggesting that AHLL was absorbed by both active transport and passive diffusion. The apparent permeability coefficients (Papp) of AHLL both from the apical (AP) to basolateral (BL) side (PappAB) and from the BL to AP side (PappBA) decreased when the temperature was lowered from 37°C to 4°C.The uptake of AHLL was more at pH 7.4 than at other pHs. Both verapamil and (E)-3-[[[3-[2-(7-chloro-2- quinolinyl) ethenyl] phenyl]-[[(3-dimethyl amino)-3-oxopropyl]thio] methyl] thio]-propanoic acid (MK571) inhibited the absorption of AHLL, indicating that P-glycoprotein and multi-drug resistant proteins (MRPs) were all involved in AHLL secretion, especially multi-drug resistant protein 2 (MRP2). AHLL was transported through both trans- and paracellular pathways across the Caco-2 cell monolayer. This work first elucidates the AHLL absorption mechanism in Caco-2 cells and provides the basis for future studies on the improvement of bioavailability.
Introduction
A
Ala-His-Leu-Leu (AHLL) is a novel ACE inhibitory peptide that is isolated from loach (Misgurnus anguillicaudatus) on the basis of controlled loach protein enzymolysis, which exhibited strong ACE inhibitory activity both in vitro and in vivo in our previous studies. 7,8 It was found to be an absorbable natural compound with antihypertensive activity in spontaneously hypertensive rats. 8 Thus, the transport behavior and absorption mechanism of AHLL in the intestinal tract needs to be evaluated.
Cell models are important means to investigate and predict intestinal absorption mechanisms. The Caco-2 cell line is derived from human colorectal adenocarcinoma and shares many morphological and functional characteristics with mature enterocytes. 9 –11 Caco-2 cells exhibit a well-differentiated brush border on their apical (AP) surfaces and at their tight junctions. 12 In addition, similar expressions of various transport systems and metabolic enzymes existing in normal intestinal epithelium were found in Caco-2 cells. 13 It has good reproducibility, applicability, and correlation between peptide absorption in the human intestine and the Caco-2 cell monolayer model; has become a standard screening tool for predicting the absorption and transport mechanism of drugs in the human intestine; and is accepted by the FDA. 14
Peptide uptake from the membrane of the Caco-2 cells is a complicated system involving concentration, pH, temperature, secretion, and an energy-dependent process. 15 Usually, sensitivity to temperature is a common feature of carrier protein, and efflux carrier protein shows strong activity only at the appropriate temperature. If the peptide was taken up by a mediated carrier, the uptake should be increased nonlinearly with the increasing concentration and should reach the saturation in a high concentration. Liu et al. found that the transport of antihypertensive peptide Val-Leu-Pro-Val-Pro (VLPVP) was related to the pH and had a dependent concentration in the Caco-2 cell model. Efflux protein multi-drug resistant protein 2 (MRP2) strongly mediated the absorption process of VLPVP. 16
Our previous investigations showed that AHLL exhibited strong ACE inhibitory activity in vivo. During the long-term oral administration of AHLL, the systolic blood pressure decreased significantly in spontaneously hypertensive rats. 8 The ACE activity of the abdomen aorta and lung were significantly reduced in the high-dose (3 mg/kg body weight) group after 2 months. This study was the continuation of our previous work. The aim was to characterize the transport behavior and mechanism of AHLL involved in the intestinal absorption by using the human intestinal epithelial Caco-2 cell line.
Materials and Methods
Materials and chemicals
The human colon adenocarcinoma Caco-2 cells were purchased from the Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences. Live loach (Misgurnus anguilliacaudatus) of 8.0 ± 1.5 g body weight and 9.0 ± 2.5 cm body length was purchased from a local market in Nanjing of China. Transwell systems were purchased from Corning Incorporated. Dulbecco's modified Eagle's medium (DMEM) was from HyClone Laboratories, Inc. Heat-inactivated fetal bovine serum (FBS), a non-essential amino acid solution, trypsin-ethylenediaminetetraaceticacid (EDTA),
Research method for AHLL transport across Caco-2 cells
The flow chart for the study of the transport of AHLL across Caco-2 cells is shown in Figure 1. First, AHLL was prepared from loach hydrolysates after consecutive purification steps. The cytoxicity of AHLL on Caco-2 cells was assessed by using the MTT assay. Then, the absorption behavior of AHLL across Caco-2 cells was studied systematically. The secretion of AHLL transport was assessed by various inhibitors of efflux carrier proteins. Then, the transport pathway of AHLL across the Caco-2 cells was investigated through the inhibitors and accelerator of three pathways, mainly for oligo-peptides across enterocytes. Finally, the mechanism of AHLL transport was obtained after a comprehensive and systematic summary.
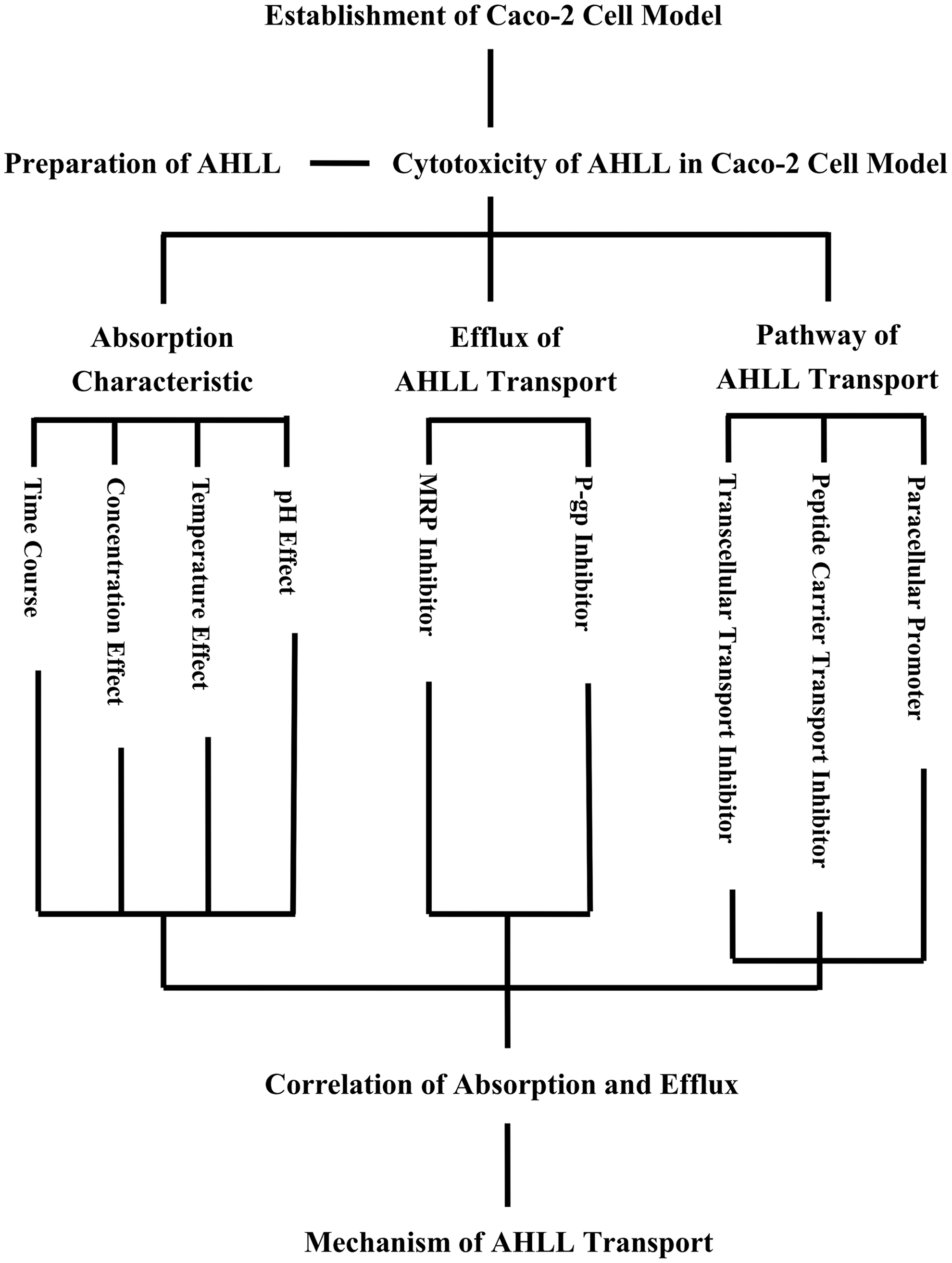
Research flow diagram for the transport of a novel angiotensin-I-converting enzyme inhibitory peptide Ala-His-Leu-Leu across Caco-2 cells. AHLL, Ala-His-Leu-Leu.
Preparation of AHLL from loach
AHLL was prepared from loach according to the method previously reported. 7 Briefly, loach protein was hydrolyzed with bromelain (3 × 105 U/g; Shanghai Kayon Biological Technology Co., Ltd.) at a ratio of 3:1000 w/w in a 55°C water bath shaker (SHZ-22) with stirring for 6 h. The hydrolysate was centrifuged, and the supernatants were filtered by ultrafiltration to obtain a fraction with a molecular weight smaller than 2.5 kDa. Then, the fraction was isolated and purified by consecutive purification steps using gel filtration chromatography and reverse-phase high performance to obtain a purified peptide AHLL (452 Da) with the IC50 (the peptide concentration required to produce 50% ACE inhibition under the described conditions) of 18.2 ± 0.9 μg/mL. The purity of the prepared AHLL was 97%.
Stability of AHLL in simulated intestinal digestive enzyme system
To simulate the human intestinal digestion process, a hydrolysis using trypsin was performed. 17 The buffer was adjusted to pH 6.8 with 0.1 mol/L potassium phosphate and 0.1 mol/L NaOH. AHLL was hydrolyzed by trypsin (E/S, 5 ‰) with moderate agitation for 4 h at 37°C. Samples were taken out at 1, 2, 3, and 4 h, and they were placed in a boiling water bath for 10 min to inactivate the enzyme. After centrifugation for 15 min at 2504 g the ACE inhibitory activity of supernatants was determined. The assay of ACE inhibitory activity in vitro assayed was according to the method in the previous paper. 7
Cell culture and Caco-2 cell model establishment
Caco-2 cells were grown in DMEM that was supplemented with 10% heat-inactivated FBS,
The transport assays were carried out in six-well Transwell plates with polycarbonate membrane inserts (24 mm diameter, pore size 0.4 μm, Costar; Corning Incorporated). In this system, the cells are seeded at a density of 3 × 105 cells/mL onto the porous membrane of the insert that separates the well into two compartments: AP (upper) and basolateral (BL, lower) ones. The cell number was determined by a direct count method using a hemocytometer. 18 The cell number per milliliter is equal to the total cellular score divided by 4 and multiplied by 10,000. Cells were seeded onto the inserts, with the addition of 1.0 mL DMEM to the AP chamber and 1.5 mL DMEM to the BL chamber. The cells were incubated at 37°C, with 5% CO2 and 95% relative humidity. Cell culture medium was changed every 1–2 days until cell differentiation was reached (10 days post-seeding). The integrity of the cell layer was evaluated by measuring the transepithelial electrical resistance (TEER) with a Millicell-ERS volt ohmmeter (Millipore Corporation). The cell monolayer considered completely formed was used for transport experiments when the TEER value was greater than 250 Ω cm2.
Assay of cell viabilities in the presence of AHLL
The effects of AHLL on the viability of Caco-2 cells were assessed by using an MTT assay. Briefly, Caco-2 cells were seeded in 96-well plates (1 × 104 cells/well) in 200 μL HBSS. The incubation was conducted at 37°C for 48 h. After that, the cells were treated with various concentrations of AHLL (0, 10, 50, 100, 200, 400, 600, 800, 1000 μg/mL) for 4 h. The culture was replaced with 100 μL MTT (1 mg/mL) for a 4 h incubation at 37°C. Then, the medium was replaced by 0.1% DMSO in 200 μL HBSS and shaken for 10 min. At the end, the absorbance was measured by a microplate reader (Tecan) at 490 nm. Each experiment was repeated six times, and the mean value was used to calculate the survival rate of cells by using Equation (1).
19
The AHLL concentrations at which the cell survival rate was above 95% were chosen as the transport dose.
Absorption characteristics of AHLL across Caco-2 cell monolayer in bidirectional transport
A transport experiment of AHLL was conducted with Caco-2 cell monolayers that were grown in the inserts of a transwell plate. The monolayer was rinsed twice with warm HBSS solution and subsequently pre-incubated (37°C, 15 min). Afterward, the HBSS solution on both sides of the cell monolayer was removed by aspiration. To measure the AP to BL transport, 1 mL HBSS containing AHLL was added to the AP side, whereas 1.5 mL blank HBSS was added to the BL side. To measure AHLL transportation from the BL to AP side, 1.5 mL HBSS containing AHLL was added to the BL side, whereas 1.0 mL blank HBSS was added to the AP side. The AHLL solution was freshly prepared in HBSS.
AHLL transport across the Caco-2 cell monolayer was assessed in different concentrations (10, 50, 100 μg/mL) at pre-established time points (0, 30, 60, 90, 120, 150 min) in both directions. To investigate the temperature effect on AHLL transport, the experiments were conducted at 4°C and 37°C in both directions. The effect of pH on AHLL transport was studied with the following pH: 6.0, 7.0, 7.4, and 8.0 in the donor compartments and pH 7.4 in the acceptor compartments.
Inhibition of AHLL transport and accumulation
The cell monolayer was pre-incubated with 100 μmol/L verapamil, 50 μmol/L MK-571, and 10 mmol/L sodium azide in the donor compartment, respectively. Without eliminating the inhibitor, 50 μg/mL AHLL solution prepared in HBSS was added to the donor compartment. Ninety minutes later, the quantification of AHLL in the acceptor media was assayed. The apparent permeability coefficients (Papp) and the efflux ratio (PDR) were calculated as Equations (2) and (3) after determining the concentration of AHLL.
The pathway of AHLL transport
Three pathways, including transcellular, paracellular, and carrier-mediated transport, for AHLL transport were evaluated. Carrier transport competitive inhibitor of Gly-Pro (10 mmol/L), transcellular transport inhibitor of phenylarsine oxide (25 μmol/L), and paracellular transport promoter of sodium deoxy cholate (100 μmol/L) were added to the AP side containing 50 μg/mL AHLL, respectively. Ninety minutes later, the quantity of AHLL in the BL media was assayed.
AHLL assays
All assays were performed by using a reverse-phase high-performance liquid chromatographic (HPLC) system (Series 1100; Agilent Technology). The analytical column was an Agilent XDB-C18 column (5 μm, 4.6 mm i.d. × 250 mm). The mobile phase contained (A) acetonitrile with 0.1% trifluoroacetic acid and (B) water with 0.1% trifluoroacetic acid. The column was eluted by using a linear gradient of 10–40% buffer A (0–10 min). The flow rate was 1 mL/min at 25°C, and its absorbance was monitored at 220 nm. A standard calibration curve was generated by using the purified AHLL standard. Peak area measurements were employed to determine the amount of AHLL in samples (The limit of detection of AHLL in the HPLC system is 0.11 μg/mL, and the limit of quantitation of AHLL in the HPLC system is 0.37 μg/mL.).
Analysis and determination of Papp and efflux ratio
The Papp (cm/sec) were calculated from Equation (2):
10
where dQ/dt is the flow rate (mg/mL/sec) determined by the linear slope of the equation defining the variation in AHLL concentration (corrected for dilution) versus time; Vr is the volume of the receptor compartment (AP 1.0 mL; BL 1.5 mL); A is the membrane surface area of the cell monolayer (4.67 cm2); and C0 is the initial concentration of AHLL in the donor compartment (mg/mL).
The efflux ratio (PDR) was calculated from Equation (3):
20
where PappBA is the apparent permeability coefficient in the BL to AP direction (cm/sec), and Papp(AB) is the apparent permeability coefficient in the AP to BL direction (cm/sec).
Statistical analysis
Results are expressed as means ± standard deviation (n = 3). The results obtained were subjected to one-way analysis of variance. Duncan's multiple-range tests were performed to determine the significant difference by using SPSS 13.0 software (SPSS, Inc.). P < .05 was considered statistically significant.
Results
Stability of AHLL in simulated intestinal digestive environment
Usually, protein dwells for 4–6 h in the intestine and is digested by the intestinal enzyme system at a pH of 6.8. In the simulated digestive environment, as shown in Figure 2, the content of AHLL decreased gradually with time increase. After a 1 h digestion by trypsin, AHLL was significantly degraded. Two hours later, there was no significant decrease in AHLL content and the retention rate of ACE inhibitory activity. At the end of digestion for 4 h, the retention rate of ACE inhibitory activity was 69%, which showed that most of the ACE inhibitory activity of AHLL remained after intestinal digestion, and AHLL exhibited a certain tolerance to intestinal digestion.
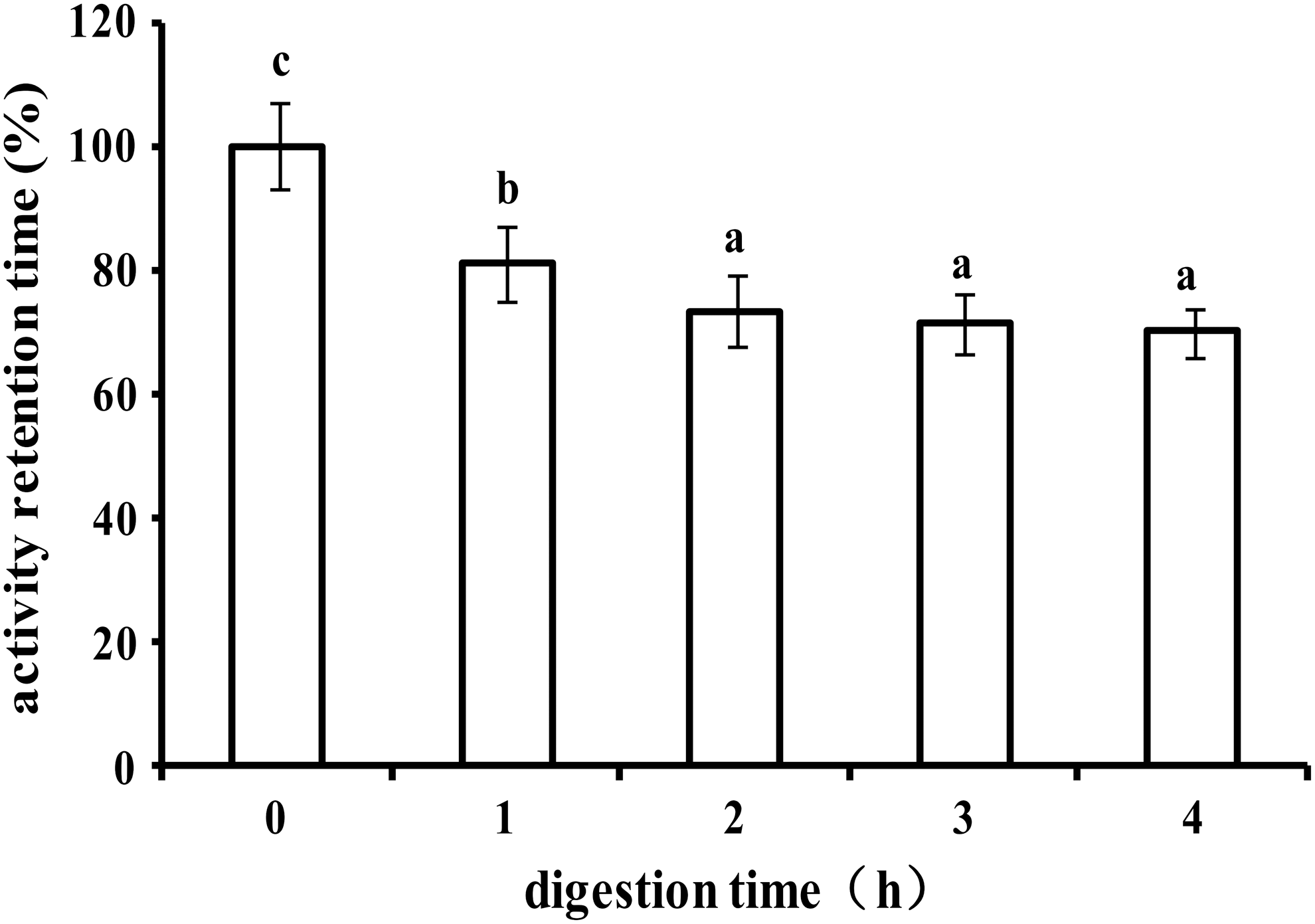
The activity retention rate of AHLL during intestinal digestion. Different lowercase letters indicate significant differences among different times (P < .05).
Cytotoxicity of AHLL
A 95% cell survival was used as the threshold to indicate the AHLL concentration sensitivity. As shown in Table 1, the survival rate of Caco-2 cells was just 93% with AHLL concentrations of 400 μg/L. When the concentration was 200 μg/mL, the survival rate of Caco-2 cells was as high as 98%. It demonstrated that the safe absorptive concentration of AHLL was 200 μg/mL.
Values are expressed as mean ± SD (n = 5).
Data values are reported as mean ± SD.
Different lowercase letters indicate significant differences among different concentrations (P < .05).
SD, standard deviation.
Time courses of the AHLL uptake across Caco-2 cell monolayers at various concentrations
The AHLL transport across Caco-2 cell monolayers was investigated over an AHLL concentration range from 10 to 100 μg/mL in both directions. The Caco-2 cells with AHLL were incubated at 37°C. The uptake of AHLL across the Caco-2 cell monolayer increased in both AP → BL and BL → AP directions with the increase of AHLL concentration and incubation time. The AHLL transport was time- and concentration dependent in a bidirectional transfer. As shown in Figures 3 and 4, the permeation of AHLL across the Caco-2 cell monolayer increased linearly within 90 min and then saturated at the test range of concentration (10–100 μg/mL).
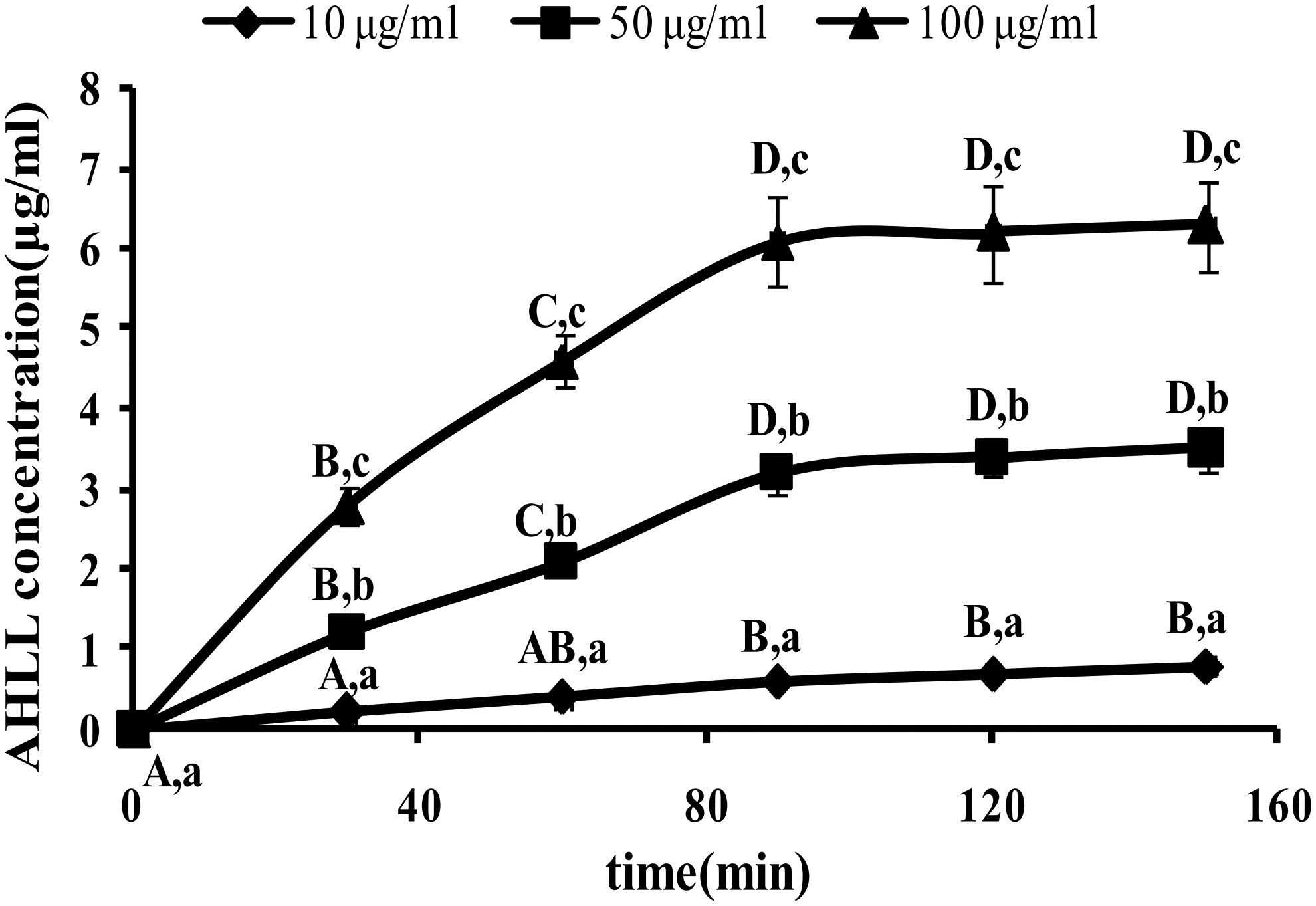
AP–BL transport in relation to time at various concentrations of AHLL (10, 50, 100 μg/mL). Different capital letters indicate significant differences among different times at the same concentration of AHLL (P < .05). Different lowercase letters indicate significant differences among different concentrations of AHLL at the same time (P < .05). AP, apical; BL, basolateral.
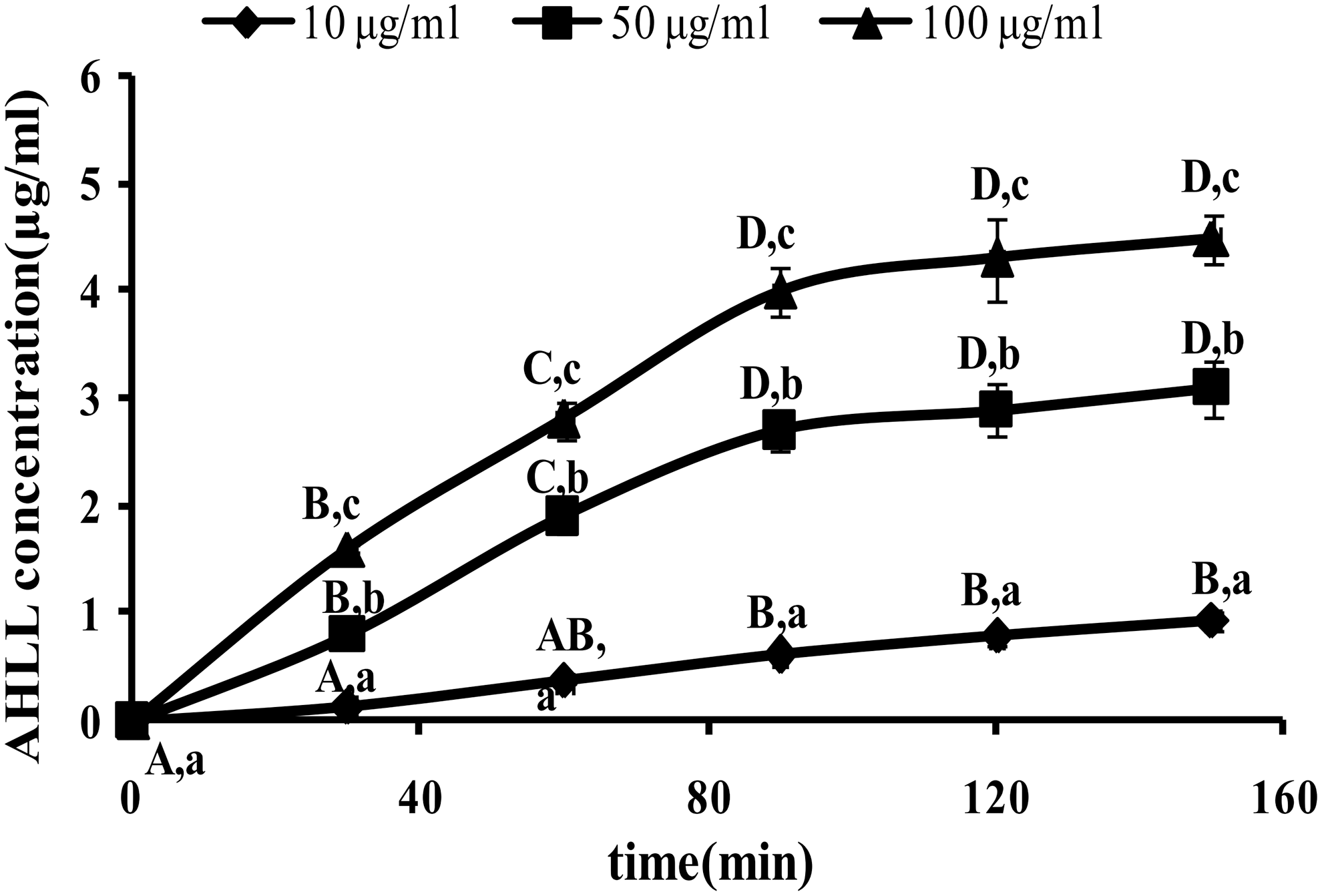
BL–AP transport in relation to time at various concentrations of AHLL (10, 50, 100 μg/mL). Different capital letters indicate significant differences among different times at the same concentration of AHLL (P < .05). Different lowercase letters indicate significant differences among different concentrations of AHLL at the same time (P < .05).
There was an obvious difference of the Papp values in the BL → AP direction with a concentration increase, whereas the Papp values remained stable in the AP → BL direction (Table 2). The efflux of AHLL was strong and the Papp value was (2.43 ± 0.23) × 10−6 cm/sec at the concentration of 10 μg/mL. When the concentration increased to 100 μg/mL, the Papp value decreased to (1.57 ± 0.21) × 10−6 cm/sec and the PDR also declined from 1.03 to 0.59.
Values are expressed as mean ± SD (n = 3).
Data are reported as mean ± SD.
Different capital letters indicate significant differences of Papp values in the AP → BL direction among different concentrations (P < .05).
Different lowercase letters indicate significant differences of Papp values in the BL → AP direction among different concentrations (P < .05).
Different Roman symbols indicate significant differences of PDR among different concentrations (P < .05).
Papp, the apparent permeability coefficients; PDR, efflux ratio; AP, apical; BL, basolateral.
Temperature effect on AHLL transport
When the temperature was lowered from 37°C to 4°C, the PappAB was changed from (2.50 ± 0.19) × 10−6 cm/sec to (1.08 ± 0.10) × 10−6 cm/sec and the PappBA was changed from (1.66 ± 0.13) × 10−6 cm/sec to (0.91 ± 0.08) × 10−6 cm/sec. In both AP → BL and BL → AP directions, the Papp obtained at 37°C were markedly higher than the values obtained at 4°C. Meanwhile, the permeability coefficient in the AP → BL direction was (1.08 ± 0.1) × 10−6 cm/sec, close to (0.91 ± 0.08) × 10−6 cm/sec obtained in the BL → AP direction at 4°C. The Papp difference between the two directions was diminished when the temperature was decreased to 4°C. Transport was obviously inhibited when the temperature was lowered from 37°C to 4°C (P < .05); similar results were obtained in other researches. 21,22
The pH effect on AHLL transport across Caco-2 cell monolayer
Caco-2 cells were incubated with 50 μg/mL AHLL at pH 6.0, 7.0, 7.4, and 8.0 in the donor compartment and pH 7.4 in the acceptor compartment for 90 min to estimate the pH effect on AHLL transport across the Caco-2 cell monolayer. As presented in Figure 5, the Papp value at pH 7.4 was higher than the Papp values measured at other pHs, and a change of pH from 7.0 to 8.0 did not significantly affect the Papp values in both AP → BL and BL → AP directions. However, the Papp values from pH 7.0 to 8.0 were significantly higher than those at pH 6.0 in both directions (P < 0.05), especially the Papp values at pH 7.4: PappAB 7.4 = (2.50 ± 0.19) × 10−6 cm/sec > PappAB6.0 = (1.74 ± 0.11) × 10−6 cm/sec, PappBA 7.4 = (1.66 ± 0.13) × 10−6 cm/sec > PappBA6.0 = (1.02 ± 0.09) × 10−6 cm/sec. AHLL transport was easier at the physiological pH of 7.4 than at the lower pH of 6.0.
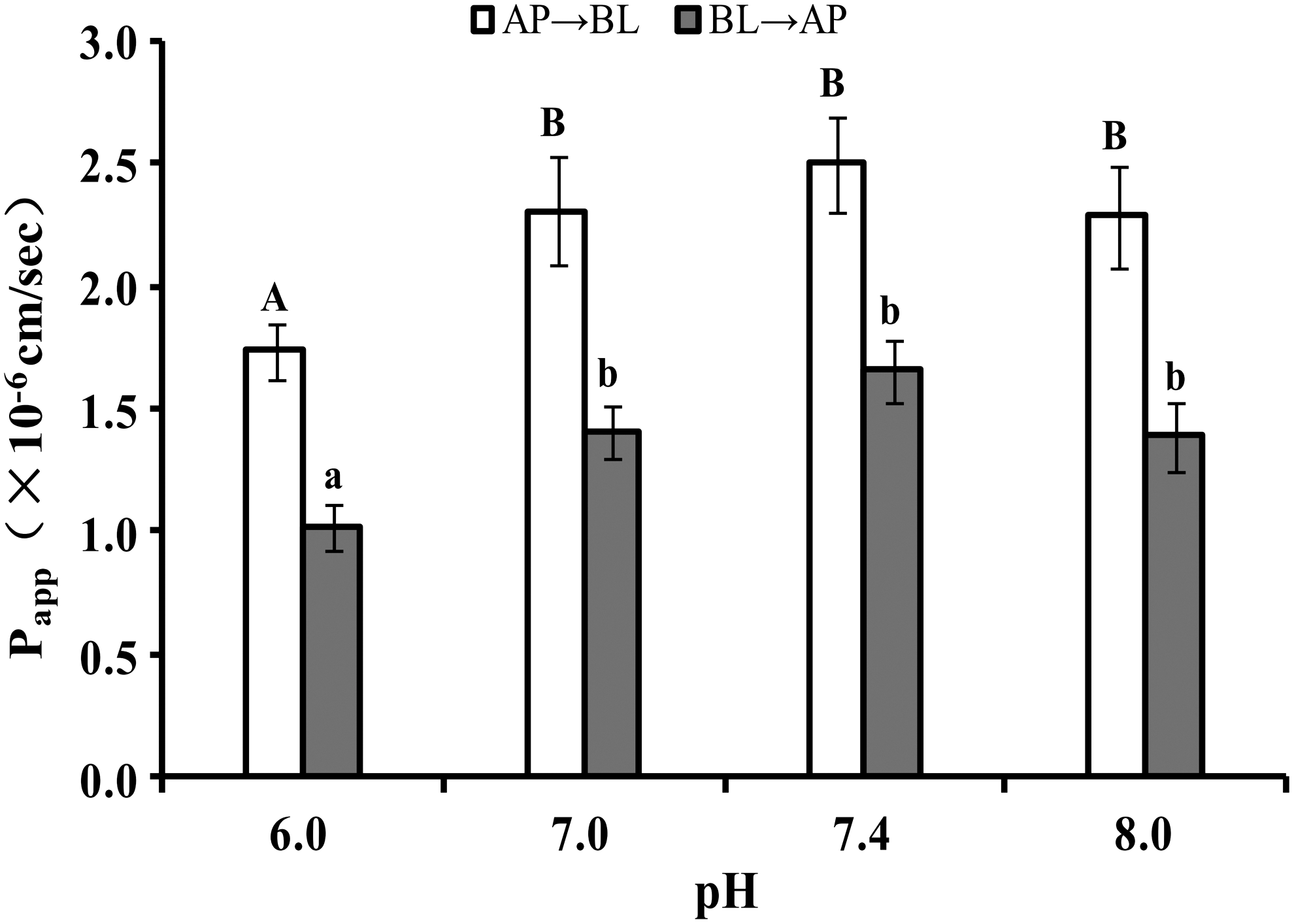
Effects of pH on AHLL transport in both directions. Different capital letters indicate significant differences among different pHs in the AP → BL direction (P < .05). Different lowercase letters indicate significant differences among different pHs in the BL → AP direction (P < .05).
Inhibition of efflux transporters on AHLL transport across Caco-2 cell monolayers
Table 3 presented the results from transport experiments by using various selective efflux transport inhibitors. When verapamil as a typical P-glycoprotein (P-gp) inhibitor was added to the AP side, the PappAB value was increased from (2.50 ± 0.19) × 10−6 cm/sec to (2.88 ± 0.20) × 10−6 cm/sec, and the PDR value was significantly decreased (P < 0.05). The results indicated that AHLL was a P-gp substrate under our experimental conditions. Sodium azide, the ATP energy generation inhibitor, significantly accelerated the absorption of AHLL from the AP side to the BL side, corresponding to the PappAB value that increased from (2.50 ± 0.19) × 10−6 cm/sec to (3.06 ± 0.22) × 10−6 cm/sec. This also illustrated that the ATP-binding cassette transporter involved the uptake of AHLL across the Caco-2 cell monolayer. MK571 was chosen as a multi-drug resistant protein (MRP) inhibitor, because it was more selective for MRPs than other inhibitors. The PappAB was significantly increased (P < .05) from (2.50 ± 0.19) × 10−6 cm/sec to (3.19 ± 0.19) × 10−6 cm/sec in the presence of MK571. At the same time, a 26% reduction in PDR was observed, which strongly indicated that AHLL transport was competitively inhibited by the MRP inhibitor.
Mean ± SD, n = 3.
Data are reported as mean ± SD.
Different capital letters indicate significant differences of Papp values in the AP → BL direction among different transporter inhibitors (P < .05).
Different lowercase letters indicate significant differences of Papp values in the BL → AP direction among different transporter inhibitors (P < .05).
Different Roman symbols indicate significant differences of PDR among different transporter inhibitors (P < .05).
MRP, multi-drug resistant protein; P-gp, P-glycoprotein.
The pathway of the AHLL transport across the Caco-2 cell monolayer
As presented in Figure 6, the concentration of AHLL in the acceptor compartment decreased from 3.90 ± 0.24 μg/mL to 3.61 ± 0.38 μg/mL in the presence of Gly-Pro, a competitive inhibitor of peptide carrier transport, but not significantly. Meanwhile, phenylarsine oxide as the inhibitor of transcellular transport inhibited the AHLL transport significantly (P < .05), corresponding to the AHLL concentration of 3.15 ± 0.19 μg/mL in the BL side. When sodium deoxy cholate was added to the AP side with 50 μg/mL AHLL, the concentration of AHLL in the BL side was significantly increased (P < 0.05). These results illustrated that transcellular and paracellular pathways were all involved in AHLL transport across the Caco-2 cell monolayer.

Effects of Gly-Pro, phenylardine oxide, and sodium deoxy cholate on the AP to BL transport of AHLL. Different lowercase letters indicate significant differences from the control (P < .05).
Discussion
The AHLL transport was time- and concentration dependent in a bidirectional transfer at a safe absorptive concentration of 200 μg/mL. The permeation of AHLL across the Caco-2 cell monolayer increased linearly within 90 min and then saturated. The same tendency was reported for inorganic mercury absorption across Caco-2 cells. 22 The platform may be caused by the lack of concentration gradient and may involve a saturable process in bidirectional transfer. 15 The efflux of AHLL transport was strong at low concentrations, but the efflux carrier could reach saturation in high concentrations, leading to a decreased PDR. The efflux ratios (PDR) in both directions were all above 0.5, suggesting that AHLL was absorbed by both active transport and passive diffusion. 23 Furthermore, the data showed that the absorption of AHLL was much greater than the secretion, meaning that active transport dominated the AHLL transport at the concentrations above 10 μg/mL. The Papp values of AHLL in both the AP → BL and BL → AP directions were 1.0 × 10−6 cm/sec, indicating that AHLL was well absorbed in human intestinal absorption.
A decrease in temperature reduces cellular metabolism and acts as an inhibitor of energy-dependent transport. 24 The decrease in PappAB was much more than in PappBA when the temperature was lowered from 37°C to 4°C (P < 0.05). The phenomenon indicated that AHLL transport was possibly energy dependent and was obviously inhibited when the temperature was lowered. The uptake of AHLL across the Caco-2 cell monolayer was affected more than the secretion by decreasing the temperature.
The AHLL transport was more active at pH 7.4 than at other pHs, which was similar to transcellular isorhamnetin transport across the Caco-2 cell monolayer. 25 This may occur because the influx and efflux transporters of AHLL were activated at different pHs. 25 –27 When the efflux transporters were critical at pH 6.0 and the influx transporters were centralized at physiological pH values, the uptake of AHLL at pH 7.4 would be more than at other pHs. Therefore, the AHLL transport across the Caco-2 cell monolayer was pH dependent.
Several studies have reported that efflux transporters such as P-gp or MRP were important biochemical barriers of bioactive substance absorption across the cells. 28 –30 It can pump out the bioactive substance in the cell, leading to the decrease of the bioavailability, which is a fatal factor, affecting the concentration of bioactive substances in the blood. An ATP-dependent mechanism was critical for the AHLL transport process. The selected ATP-binding cassette transporters (P-gp, MRPs) were all present during AHLL secretion, which were fatal biochemical barriers in AHLL absorption. Furthermore, MRP2 and MRP3 are expressed more than other MRPs in Caco-2 cells. MRP2 and MRP3 are located at the AP sides and BL sides of the Caco-2 cells separately. 31 PappAB was significantly increased, but no significant difference of PappBA was observed in the presence of MK571. Therefore, AHLL might have been preferentially effluxed by MRP2.
Transcellular transport, paracellular transport, and peptide-mediated transport are three pathways that are mainly meant for oligo-peptides across enterocytes. The inhibitor of transcellular transport significantly inhibited the AHLL transport. At the same time, the promoter of paracellular transport significantly accelerated the AHLL transport. These illustrated that the AHLL could be transported across the Caco-2 cell monolayer through both trans- and paracellular pathways.
To the best of our knowledge, this is the first study to reveal the transport behavior and mechanism of AHLL across human intestinal epithelial Caco-2 cells. The AHLL transport across Caco-2 cells involved passive diffusion and active transport through trans- and paracellular pathways. However, the secretion of AHLL in Caco-2 absorption still exists to some extent. How to control the secretion of AHLL and improve the absorptive efficiency will be our future research focus.
Footnotes
Acknowledgment
This work was supported by a grant from the National Science Foundation of China (No. 31301439), whose financial support is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.