
Abstract
Gelidium amansii is an edible and economically important red alga consumed in South Eastern Asia. In previous studies, we reported that the ethanol extracts of G. amansii (GAE) has promising modulatory activity with respect to the morphological and functional maturation of hippocampal neurons in culture. In this study, we show that the chloroform (CHCl3) subfraction of GAE and the ethyl acetate (EtOAc) fraction dose-dependently promoted neurite outgrowth, and their effects were comparable with that of GAE. We further assessed in cultured cortical neurons, proteins differentially expressed in the presence/absence of the GAE, CHCl3 subfraction, and the EtOAc fraction by 2D-PAGE and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Proteomic data revealed that a number of proteins responsible for multiple cellular and biochemical functions vital for neuronal development and maturation were significantly upregulated in neurons treated with the GAE, CHCl3 subfraction, and the EtOAc fraction. Of the identified proteins, profilin 2a, septin 7, cdc42, protein phosphatase 2A, DA11, eukaryotic translation initiation factor 5A-1, and γ-enolase are known to play important roles in neuritogenesis and dendritic arborization. Immunofluorescence data demonstrate that GAE-treated hippocampal neurons showed greater intensity ratios in the expressions of the septin 7 and cdc42 compared to vehicle control, validating their proteomic profiles. Together these results suggest that the GAE/CHCl3 subfraction and EtOAc fraction promote neurite development by up or downregulating several key proteins.
Introduction
P
Dendritic atrophy with synapse and spine loss is observed in the aging human brain. 3 Reduction in dendritic arbor size is also associated with psychiatric illnesses, such as schizophrenia and major depressive disorder, as well as neurodegenerative diseases, such as Alzheimer's disease. 4,5 Neurotrophins have proven to promote neuritogenesis, synaptogenesis, and neuronal survival. 6 –8 The brain-derived neurotrophic factor (BDNF) promotes neurogenesis. 9,10 BDNF gene transfection triggers dendritic and axonal branching in dentate gyrus granule cell cultures. 11 Therefore, there are strong rationales suggesting that increasing supply of the neurotrophins to degenerating neurons may be a potent way to restore neuronal function in neurodegenerative conditions.
Identification of neuritogenic and neurotrophic molecules from natural sources may provide important drug leads for activation of neuroregeneration in various forms of neuronal injuries. Macroalgae have been explored as potential sources of bioactive compounds and have been reported to possess various pharmacological effects, including antioxidative, antiaging, anticancer, 12 and antiobesity 13 activities. Furthermore, several authors have demonstrated the suitability of marine algae for neurotrophic research. 14 –16 In addition, recently, we reported the neurotrophic activities of marine algae, particularly Gelidium amansii, Sargassum fulvellum, and Undaria pinnatifida, in primary cultures of hippocampal neurons. 17 –19
In this study, we performed proteomic analysis to identify molecular target proteins up- or downregulated during neuronal development in vitro. We demonstrate that the ethanol extracts of G. amansii (GAE), CHCl3 subfraction, and EtOAc fraction can promote neurite outgrowth by regulating the expressions of proteins responsible for neurite synthesis and maturation.
Materials and Methods
Extraction and fractionation of G. amansii
The ethanol extract of G. amansii was prepared as described previously. 18 Briefly, a pulverized algal sample was extracted with 95% aqueous ethanol and filtered through sterile cotton. The filtrate was concentrated in vacuo and completely dried to produce GAE. The filtration residue was re-extracted with EtOAc to obtain the EtOAc fraction. A flowchart of solvent fractionation is provided in Figure 1 (modified). 20 GAE, its subfractions, and the EtOAc fraction were dissolved in dimethyl sulfoxide (DMSO) at 16 mg/mL and stored in foil-wrapped vials at −20°C in the dark before experiments.

Flowchart of the extraction and fractionation procedure.
Primary neuronal culture and treatment
Animal care and use were in accord with the institutional guidelines issued by the Institutional Animal Care and Use Committee of the Dongguk University College of Medicine and were approved by the same committee. Rat (Sprague-Dawley) primary neuronal cultures (dissociated hippocampal cells for immunocytochemistry and dissociated cortical cells for proteomics) were prepared as previously described
18,21
with the following exceptions: dissociated cells were seeded onto poly-
Fluorescence labeling with DiO
At DIV3 (3 days in vitro), neurons were live-stained with Vybrant DiO (Molecular Probes, Eugene, OR, USA), according to the manufacturer's instructions. DiO is a lipophilic dye that binds to plasma membranes, and thus, enables entire neurons to be visualized.
Image acquisition and analysis
A Leica Research Microscope (DM IRE2) equipped with I3 S, N2.1S, and Y5 filter systems (Leica Microsystems AG, Wetzlar, Germany) was used for phase-contrast and epifluorescence microscopy. Images (1388 × 1039 pixels) were acquired using a high-resolution CoolSNAP™ CCD camera (Photometrics, Tucson, AZ, USA) using Leica FW4000 software. Morphometric analyses and quantification were performed using ImageJ (version 1.45) software with the simple neurite tracer plug-in (National Institute of Health, Bethesda, MD, USA). Fluorescent images taken of DiO stained cultures were converted into grayscale mode and inverted. Morphometric parameters, such as the numbers of primary neurites (neurites that originated directly from soma), total primary neurite lengths (the sum of the lengths of primary neurites), and the numbers of branching points, were measured.
Extraction of protein sample
At DIV6, proteins were harvested from cultured neurons using the Nuclei EZ Prep Nuclei Isolation Kit (Sigma-Aldrich). Protein concentrations were determined using the Bradford method. 22
2D-PAGE and image analysis
2D-PAGE was performed on whole-cell lysate protein fractions. IPG dry strips (4–10 NL IPG, 24 cm; Genomine, Inc., Pohang, Korea) were equilibrated for 12–16 h in a 7 M urea, 2 M thiourea solution containing 2% 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS), 1% dithiothreitol (DTT), and 1% Pharmalyte and loaded with 200 μg of samples. Isoelectric focusing (IEF) was performed at 20°C using a Multiphor II electrophoresis unit and EPS 3500 XL power supply (GE Healthcare, Little Chalfont, UK), according to the manufacturer's instructions. For IEF, the voltage was linearly increased from 150 to 3500 V over 3 h for sample entry followed by a constant 3500 V. Complete focusing was achieved at after 96 kVh. Before the second dimension, strips were incubated for 10 min in equilibration buffer (50 mM Tris-Cl, pH 6.8 containing 6 M urea, 2% SDS, and 30% glycerol), first with 1% DTT and second with 2.5% iodoacetamide. Equilibrated strips were inserted into SDS-PAGE gels (20 × 24 cm, 10–16%). SDS-PAGE was performed using the Hoefer DALT 2D system (Amersham Biosciences), according to the manufacturer's instructions. Two-dimensional gels were run at 20°C for 1700 Vh and then silver stained as described. 23 Quantitative analysis of digitized images was carried out using PDQuest software (version 7.0, Bio-Rad Laboratories, Hercules, CA, USA), according to the manufacturer's instructions. Spot intensities were normalized versus total valid spot intensities. Protein expressions over or under 1.5-fold versus the vehicle control were further analyzed.
Protein identification by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry and peptide mass fingerprinting
For protein identification by peptide mass fingerprinting (PMF), protein spots were excised, digested with trypsin (Promega, Madison, WI, USA), mixed with α cyano-4-hydroxycinnamic acid in 50% acetonitrile containing 0.1% TFA, and subjected to matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) analysis (Microflex LRF 20; Bruker Daltonics, Billerica, MA, USA), as described.
24
Spectra were collected using 300 shots per spectrum over the m/z range 600–3000 and calibrated by two-point internal calibration using trypsin autodigestion peaks (m/z 842.5099 and 2211.1046). Peak lists were generated using Flex Analysis version 3.0. The thresholds used for peak-picking were as follows: 500 for minimum resolution of monoisotopic mass and 5 for S/N. The search program MASCOT, developed by The Matrix Science (
Immunocytochemistry
Neurons on coverslips were rinsed briefly with Dulbecco's phosphate-buffered saline (D-PBS; Invitrogen, Carlsbad, CA, USA) and fixed by a sequential paraformaldehyde/methanol fixation procedure 25 on DIV12. For immunostaining, the following antibodies were used: primary antibodies to tubulin α-subunit (mouse monoclonal 12G10, 1:1000 dilution; Developmental Studies Hybridoma Bank, University of Iowa, USA); Septin 7 (rabbit polyclonal, 1:500 dilution; Santa Cruz Biotechnology, Inc., Dallas, TX, USA); and Cdc42 (rabbit polyclonal, 1:200 dilution; Thermo Fisher Scientific, Waltham, MA, USA) and secondary antibodies (Alexa Fluor 488-conjugated goat anti-mouse IgG [1:1000] and Alexa Fluor 568-conjugated donkey anti-rabbit IgG [1:1000], Molecular Probes). Fixed neurons were incubated with primary antibody followed by secondary antibody and mounted on slides as described. 25 Fluorescent images (1388 × 1039 pixels) were acquired as described in an earlier section. Puncta for Septin 7, Cdc42, and α-tubulin were counted per 50 μm length of dendrite using puncta analyzer plug-in. 26 The puncta intensity ratios for Septin 7/α-tubulin and Cdc42/α-tubulin were then measured.
Statistical analysis
Results are expressed as the means ± standard error of means (SEMs) of at least three independent experiments. Statistical comparisons were made using one-way analysis of variance (ANOVA) with Duncan's post hoc multiple comparisons using SPSS ver. 16.0. Statistical significance was accepted for P values <.05.
Results
Neurite outgrowth promoting activities of GAE, its subfractions, and of the EtOAc fraction
In a previous report, we showed that GAE has promising neuritogenic activity in primary neuronal cultures. 18 In this study, we investigated whether GAE subfractions and the EtOAc fraction have similar neuritogenic activities. Hippocampal neurons were incubated with various concentrations of GAE or its subfractions (CHCl3, methanol [MeOH], CHCl3-MeOH, or aqueous subfractions) or the EtOAc fraction (Fig. 1). In this study, we only present the data of fractions that exhibited visible neuritogenic activity. Typical microscopic fields of cultures grown with GAE, or its CHCl3 subfraction, or EtOAc fraction at 3.75 to 30 μg/mL for 3 days are shown in Figure 2A. Both the EtOAc fraction and the CHCl3 subfraction significantly and dose-dependently increased neurite numbers, lengths, and branchings. Furthermore, their effects were comparable with that of GAE (Fig. 2B). Notably, at 15 and 30 μg/mL, the CHCl3 subfraction had little inhibitory effect, but at lower concentrations it had neuritogenic effects.
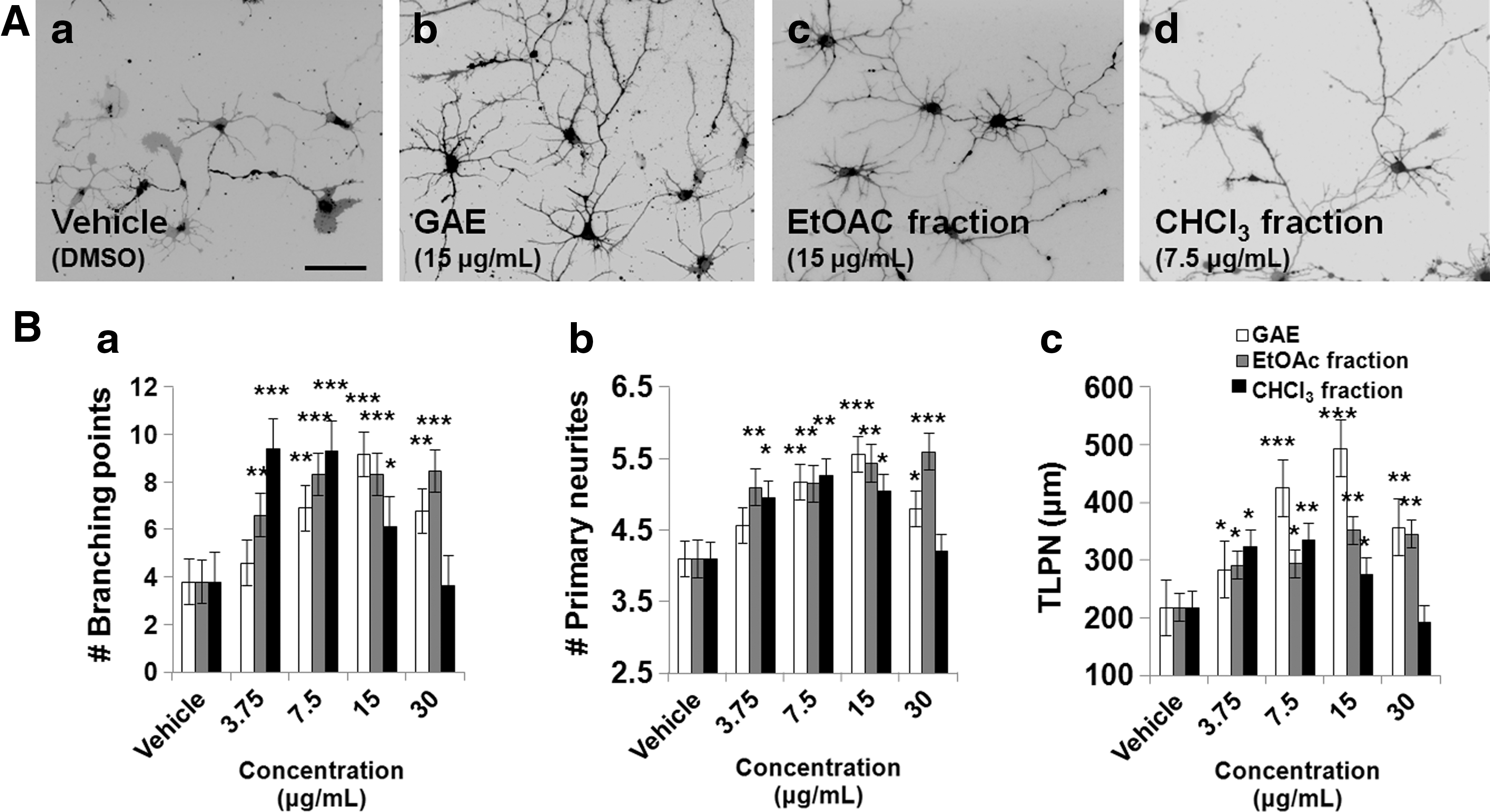
Dose-dependent neuritogenic activities of GAE, the CHCl3 subfraction, and of the EtOAc fraction in embryonic (E19) rat hippocampal neurons cultured on poly-
Proteomic profiles of cortical neurons treated with GAE, its subfractions, or the EtOAc fraction
2-DE based proteomic analysis was carried out on cortical neurons treated with GAE, the CHCl3 subfraction, the EtOAc fraction, or vehicle to evaluate changes in protein expression profiles. Based on their neurite outgrowth promoting activity, we used GAE and EtOAc fraction at 15 μg/mL and CHCl3 subfraction at 7.5 μg/mL for proteomic analysis. Because large amounts of sample are needed for proteomic analysis, cortical neurons were used instead of hippocampal neurons. In primary neuronal cultures, the neurite promoting activity of extracts commenced as soon as undifferentiated neurons attached to surfaces. Although dendritic arbor formation started after 4 days in culture, the molecular events associated with dendritic arborization were induced between day 5 and 6 in culture. Thus, 6 days of incubation was chosen for proteomic analysis. Representative 2D-PAGE images are shown in Figure 3. More than 800 protein spots were detected in each gel, and 200 spots were differentially expressed. Among them, 14 showed significant upregulation and 4 significant downregulation (indicated by circles in Fig. 3). Figure 4 shows magnified spot images of the identified proteins and the relative intensities of each protein spot.

Representative 2-DE gel images. Embryonic (E19) rat cortical neurons were cultured on poly-
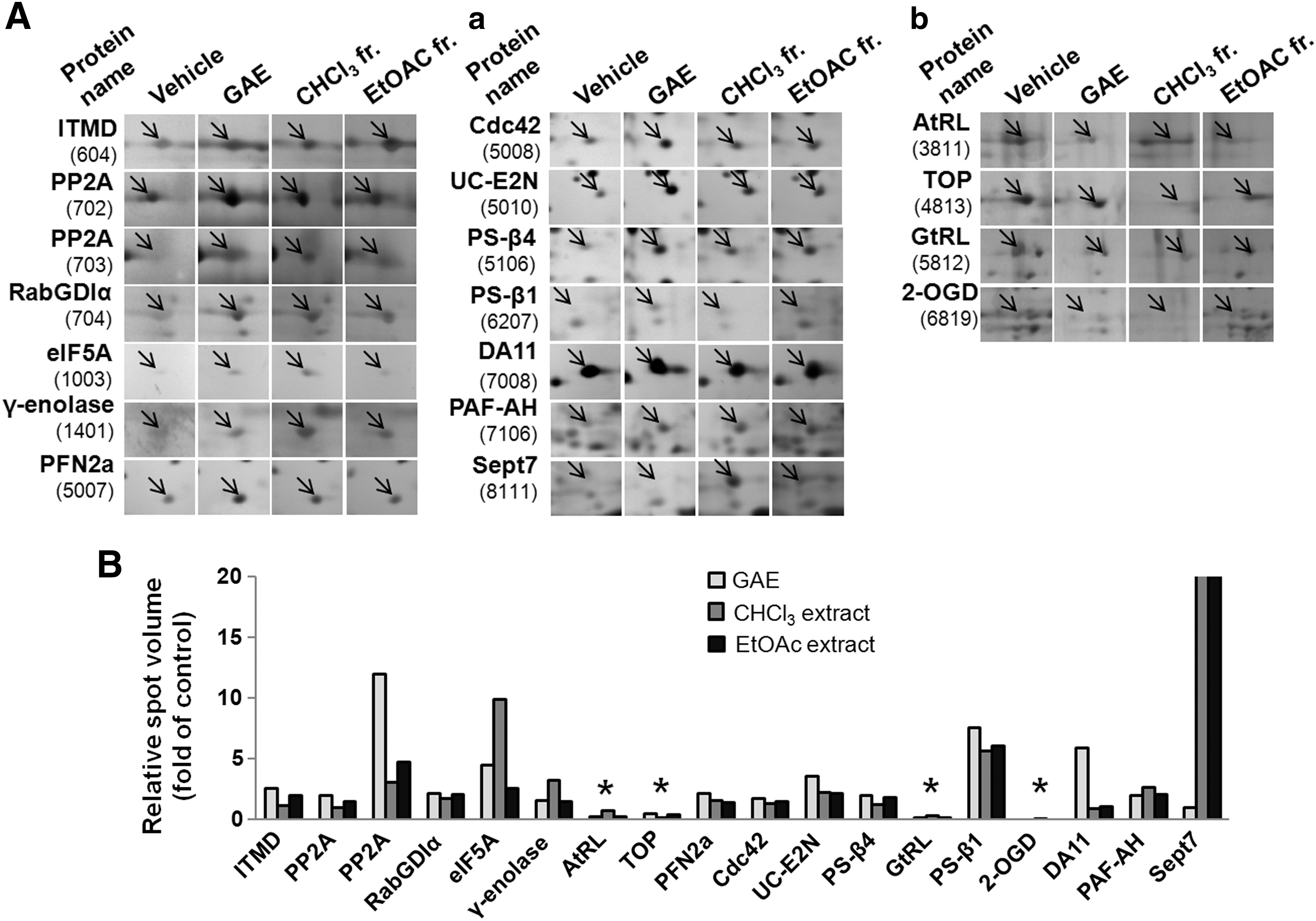
Intensities of differentially expressed proteins.
Identification of differentially expressed protein spots by MS
A total of 17 proteins were unequivocally identified by MALDI-TOF-MS mass spectrometry (MALDI-TOF-MS) and PMF. The characteristics of the identified proteins are summarized in Table 1.
GAE, ethanol extracts of Gelidium amansii.
Validation of proteomic data by immunocytochemistry
We then performed immunocytochemistry of some representative upregulated proteins such as Septin 7 and Cdc42 to validate their proteomic results using hippocampal neuronal cultures, because they are more homogeneous than cortical ones. Septin 7 is involved in cytoskeleton formation, and Cdc42 is involved in the formation of filopodia and neurite outgrowth. In this study, as in proteomic analysis, we used GAE and EtOAc fraction at 15 μg/mL and CHCl3 subfraction at 7.5 μg/mL for immunocytochemical analysis. The expressions of septin 7 and cdc42, as determined by the ratios of the fluorescence intensities of septin 7 to α-tubulin and cdc42 to α-tubulin, were significantly higher in the CHCl3 subfraction and the EtOAc fraction-treated hippocampal neurons than vehicle controls (Fig. 5), suggesting their potentiating roles not only in earlier stages of maturation but also to advanced stages of development. However, GAE itself did not induce the expression of septin 7, but significantly promoted cdc42 expression.

Immunocytochemical validation of the expression of some proteomics-identified proteins. Hippocampal neurons were cultured on poly-
Discussion
The expressions of multiple proteins within neurons are attributable to the effects of NFs. Several authors have described the NF-regulated expressions of proteins vital for neuronal differentiation, maturation, and survival. 27,28 In a previous study, we demonstrated that GAE (the filtrate fraction of G. amansii ethanol extracts) has a dose-dependent neurotrophic effect in primary hippocampal neurons. 18 In the present study, we further fractionated G. amansii into different subfractions, and we found that the GAE, its CHCl3 subfraction, and the EtOAc fraction had neurotrophic activities comparable with that of GAE, whereas the CHCl3-MeOH, MeOH, and aqueous subfractions of GAE did not exhibit any neurotrophic activity. These findings suggest that GAE contains NF-like compounds that are predominantly extracted by a nonpolar solvent CHCl3, which suggests they are phenolics. 20 The CHCl3 subfraction showed maximum neuritogenic activity at 7.5 μg/mL concentration, but higher concentrations, especially 30 μg/mL, significantly reduced the activity to the control level. This might be due to the presence of some negative ingredients within CHCl3 subfraction.
We performed proteomic analysis to explore changes in protein expressions in cortical neurons in response to the presence or absence of the GAE, CHCl3 subfraction, or its EtOAc fraction. As a result, 18 differentially expressed proteins were identified by 2D-PAGE and MALDI-TOF-MS.
Proteins involved in dendritic arborization
Profilin 2a (spot 5007)
Profilins are key regulators of actin dynamics and thus play roles in the actin-dependent regulation of neuronal architecture. Among the different isoforms, profilin 2a (PFN2a) was upregulated in extract-treated neurons compared with control neurons. PFN2a deficient neurons display significantly less dendritic complexity and fewer spine numbers. 29 In another report, it was suggested that profilins have postsynaptic functions with respect to dendritic spine stabilization and synaptic plasticity. 30 Thus, it appears that profilin 2a may be involved in dendritic outgrowth in extract-treated neurons.
Septin 7 (spot 8111)
Septin 7 (Sept7) was also highly upregulated in neurons treated with the EtOAc fraction or the CHCl3 subfraction. Septins are involved in cytokinesis, membrane trafficking, and structural scaffolding and to do so localize at the bases of dendritic protrusions and dendritic branch points. 31 –33 Furthermore, the overexpression of Sept7 increases dendrite branching and the densities of dendritic protrusions, whereas knockdown of Sept7 by RNA interference reduces dendrite arborization. Thus, it seems that Sept7 is critical for spine morphogenesis and dendrite development during neuronal maturation. 31,32 However, why the CHCl3 subfraction but not GAE itself upregulated Sept7 expression is inexplicable at present.
Proteins involved in neurite outgrowth
Cdc42 (spot 5008)
Many cellular processes that lead to changes in neuronal morphology are under the control of different small GTPases, 25,34 for example, Cdc42 is involved in the formation of filopodia and neurite outgrowth. In the present study, Cdc42 (spot 5008) was upregulated in CHCl3 subfraction and EtOAc fraction–treated neurons, which suggest that the neurotrophic activities of these fractions are mediated through the upregulation of Cdc42.
Protein phosphatase 2A (spot 702 and 703)
Protein phosphatase 2A (PP2A) is an important serine/threonine protein phosphatase and has been suggested to be crucial for neural development and normal function of the eukaryotic nervous system. 35 Liu et al. 36 reported that the upregulation of PP2A promotes extensive outgrowth of long neurites in Neura2A cells and the developments of long single axons or multiple axons in hippocampal neurons. Furthermore, zinc-induced tau hyperphosphorylation, which is responsible for neurofibrillary tangle formation in Alzheimer's disease, was completely reversed by the pharmaceutical and genetic upregulation of PP2A. 37 In the present study, PP2A was upregulated several fold when neurons were treated with the CHCl3 subfraction or the EtOAc fraction, which suggests that they may have therapeutic value during the early phase of AD.
DA11 (spot 7008)
DA11 is a fatty acid binding protein, and the strong association between DA11 gene expression and CNS development suggests that DA11 plays important roles in axonal growth and neuronal differentiation in many different neuronal populations. 38 These previous findings indicate that the upregulation of DA11 (spot 7008) by GAE is related to the neurotrophic activity of GAE.
Eukaryotic translation initiation factor 5A-1 (eIF5A; spot 1003)
eIF5A is an important translation machinery protein that is responsible for neuronal survival and neurite extension in the nervous system 39 and was found to be upregulated by GAE, the CHCl3 subfraction, and the EtOAc fraction. eIF5A, the only known protein containing the polyamine-derived amino acid hypusine, mediates neurotrophic and neuroprotective actions of nerve growth factor in neurite outgrowth and cell survival of PC12 cells. 40
γ-enolase (spot 1401)
γ-enolase was upregulated in CHCl3 subfraction and EtOAc fraction–treated neurons. γ-enolase is a highly conserved cytoplasmic glycolytic enzyme involved in cell differentiation and growth, 37,41 and neuroprotective and neurite outgrowth effects have been reported for the C-terminal part of γ-enolase. 42,43
Other proteins
Other upregulated proteins include a number of proteins involved in neurotransmission, such as Rab GDP dissociation inhibitor alpha (RabGDIα, spot 704), proteins involved in protein degradation such as ubiquitin-conjugating enzyme E2N (UC-E2N and spot 5010), proteasome subunit, beta type 4 (PS-β4 and spot 5106), and proteasome subunit alpha type-1 (PS-α1 and spot 6207), and proteins in other metabolic pathways such as iodothyronine 5′ monodeiodinase (spot 604) and platelet-activating factor acetylhydrolases (spot 7106). In contrast, a mitochondrial enzyme 2-oxoglutarate dehydrogenase (2-OGD and spot 6819), a neuropeptide-metabolizing enzyme thimet oligopeptidase (spot 4813), 44 and protein biosynthesis enzymes alanine-tRNA ligase (spot 3811) and glycine-tRNA ligase (spot 5812) were downregulated.
We believe up- or downregulations were caused by the net effects of multiple ingredients in each of these extracts. Further functional studies are required to elucidate the mechanisms by which these proteins are involved in the neurotrophic effects of these extracts.
Footnotes
Acknowledgments
This research was partially supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning to ISM (2015R1A2A2A01007104). M.A.H. wishes to thank the Korean National Institute of International Education (NIIED) for graduate assistance.
Author Disclosure Statement
No competing financial interests exist.