
Abstract
Humulus japonicus (HJ), popularly known as Japanese hops, is a traditional herbal medicine widely used for the treatment of pulmonary disease, skin disease, and hypertension in Korea. HJ exerts scavenging effects against reactive oxygen species (ROS), such as superoxide radical, hydroxyl radical, and hydrogen peroxide. Moreover, dysfunction and damage of mitochondria elicited by ROS are of critical importance in the pathogenesis of Parkinson's disease (PD). The present study aimed to examine neuroprotective potential of extracts of HJ using in vitro and in vivo 6-hydroxydopamine (6-OHDA) models. SH-SY5Y cells were cultured to explore the mechanisms for the neuroprotective effect of HJ in vitro. Unilateral 6-OHDA-induced mouse model of PD was established to investigate the neuroprotective effect of HJ on dopaminergic neurons in substantia nigra pars compacta (SNc) and striatum in vivo. Methanol extract of HJ (HJM) significantly attenuated cytotoxicity and the mitochondrial apoptosis pathway caused by 6-OHDA in SH-SY5Y cells. In addition, HJM significantly increased glutathione levels and decreased phosphorylation of ERK1/2 in SH-SY5Y cells exposed to 6-OHDA. In the in vivo study, the administration of methanol or ethanol extract of HJ improved the motor dysfunction and notably reduced dopaminergic cell death and fiber loss in the SNc and striatum caused by 6-OHDA. Our findings demonstrate that HJ may have therapeutic potential to protect dopaminergic neuron degeneration in Parkinson's disease.
Introduction
P
6-Hydroxydopamine (6-OHDA) as a neurotoxin provokes the formation of reactive oxygen species (ROS) and causes dopaminergic neuronal cell death in rodents following intrastriatal injection. 5 –7 Due to the richness in polyunsaturated fatty acids and iron content and high utilization of oxygen, the brain is very susceptible to oxidative stress injury 8 ; which leads to dysfunction and damage of mitochondria and is ultimately critical to the pathogenesis of PD. 2,9 Oxidative damage caused by 6-OHDA has been attributed to the activation of mitogen-activated protein kinase (MAPK) pathway, including p38 MAPK, c-Jun N-terminal kinase (JNK), and extracellular signal-regulated kinase 1/2 (ERK 1/2), and the MAPK signaling is closely related to 6-OHDA toxicity and neuronal apoptosis. 10 –12 It has been reported that antioxidants prevent oxidative damage and can reduce the risk of PD. 8,9 Moreover, growing evidence indicates that herbal extracts have potential benefits in attenuating the progression of PD in vitro and in vivo due to their neuroprotective actions as antioxidants, monoamine oxidase inhibitor free radical scavengers, chelators of harmful metal ions, and modulators of cell survival and apoptosis. 13 –15
Humulus japonicus Siebold & Zucc., a perennial herb in the Cannabaceae family, is commonly distributed in Asian countries, including Korea, Japan, and China. Previously, anti-inflammatory, antitumor, antimutagenic, antimicrobial, and antimycobacterial properties of extract from H. japonicus (HJ) have been reported. 16 –18 Moreover, the protective roles of HJ on neurodegenerative disease, such as Alzheimer's disease, and aging have been reported in recent studies. 19,20 Especially, the extract of HJ possessed strong scavenging effects on active oxygen molecules such as superoxide radicals, hydroxyl radicals, and hydrogen peroxide. 16,17,20 These results indicate that the HJ extracts are a good source of natural antioxidants, which might be helpful in preventing or delaying the progress of PD. However, the effects of HJ on PD have not been elucidated. In the present study, we investigate whether HJ has a beneficial effect on 6-OHDA-induced cytotoxicity in in vivo and in vitro systems, and demonstrated that HJ has protective properties that attenuate experimentally induced PD pathology.
Materials and Methods
Preparation of HJ
HJ was purchased from Gangwon Herbs, Gangwon, Korea, on July 2014. The voucher specimen was identified by Prof. Won-Keun Oh, and a voucher specimen (SNU-2014-0004) was deposited at the College of Pharmacy, Seoul National University, Korea. Then, the HJ extract was prepared and supplied by the Korea Bioactive Natural Material Bank (Seoul, Korea). The dried aerial parts of HJ were soaked in 100% methanol or ethanol in an extraction container for 2 days at room temperature. The methanol- or ethanol-soluble extracts of HJ were filtered through cheesecloth, concentrated exhaustively, and dried to produce a methanolic or ethanolic extract under reduced pressure. The methanol extract (HJM) and ethanol extract (HJE) of HJ were used in this study. The HJ extract was suspended in 0.5% carboxymethyl cellulose (CMC) at a concentration of 50 mg/mL as a stock solution, and the working solution of HJ was adjusted to the intended concentrations for use in the in vitro and in vivo experiments in the study.
Cell culture
Human neuroblastoma SH-SY5Y cells (Korean Cell Line Bank, Seoul, Korea) were cultured in Dulbecco's modified Eagle's medium (Gibco-BRL, Gaithersburg, MD)/F12 supplemented with 10% fetal bovine serum and antibiotics. Cells were maintained at 37°C in a humidified atmosphere of 5% CO2/95% air. Undifferentiated SH-SY5Y cells were seeded into well plates of suitable size and allowed to adhere to the culture plates for 2 days before experimentation. The cells were pretreated with HJ or dimethyl sulfoxide (DMSO) for 1 h and then incubated with 6-OHDA during the indicated time period.
Cell viability assay
Cells were incubated with 3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide (MTT) reagent (5 mg/mL, Sigma-Aldrich Co., St. Louis, MO), which is taken up only by viable cells, in 1/10 of the volume of the culture medium for 4 h at 37°C. An equal volume of DMSO was added to the cells to dissolve insoluble formazan formed in intact cells, followed by incubation in the dark at 37°C until completely dissolved. The absorbance was measured at 570 nm with microplate reader.
Measurement of intracellular glutathione level
The intracellular glutathione (GSH) level in the 6-OHDA-treated cells was quantified by using a GSH assay kit (Cayman Chemical, Ann Arbor, MI, USA) according to the manufacturer's instruments. Cell lysates were used for measurement of their GSH level at 405 nm.
Western blot analysis
Western blot was performed as described previously.
21
SH-SY5Y cells were seeded into six-well plates of suitable size and allowed to adhere to the culture plates for 2 days before experimentation. Cells were pretreated with HJ or DMSO for 1 h and then incubated with 6-OHDA for 24 h. Cells were centrifuged and cell pellets were resuspended in an ice-cold homogenization buffer (50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1% Nonidet P-40, 0.1% sodium dodecyl sulfate [SDS], and 0.1% sodium deoxycholate) containing a cocktail of protease inhibitors (Roche, Mannheim, Germany). In mice, 10 days after the 6-OHDA injection, the brain tissue was quickly removed and homogenized in the homogenization buffer containing a cocktail of protease inhibitors (Roche). In addition, to detect cytochrome c in the cytosol, after exposure to 50 μM 6-OHDA in the absence or presence of HJ for 24 h, the cell was pelleted by centrifugation and resuspended in ice-cold lysis buffer (210 mM
Animals
Eight-week-old male C57BL/6J mice weighing 23–28 g, derived from The Jackson Laboratory (Bar Harbor, ME, USA), were maintained at the Laboratory Animal Resource Center of KRIBB. All mice were housed in regular polycarbonate plastic cages in a temperature-controlled (21–22°C) and humidity-controlled (50–60%) environment with a 12-h light/12-h dark cycle. For HJM experiments, mice were randomly assigned to two groups: vehicle (0.5% CMC)-treated mice (n = 10) and HJM-treated (n = 11) mice and then fed HJM at 500 mg/kg/day of dose for 3 days before 6-OHDA injection and for 10 days after 6-OHDA injection. For HJE experiments, mice were randomly assigned to three groups: vehicle-treated (0.5% CMC, n = 16), 300 mg/kg HJE-treated (n = 8), and 500 mg/kg HJE-treated (n = 9) mice and the HJE was administered by oral gavage for 3 days before 6-OHDA injection and for 10 days after 6-OHDA injection. All animal experiments were approved by the Institutional Animal Care and Use Committee of KRIBB (KRIBB-AEC-15025 and KRIBB-AEC-16004) and were performed in accordance with the institutional guidelines at KRIBB.
6-OHDA lesion
6-OHDA lesion was performed as described previously. 21 Mice were anesthetized with a mixture of ketamine hydrochloride and xylazine hydrochloride, and were mounted in a stereotactic frame equipped with a mouse adaptor. Mice were pretreated with desipramine (25 mg/kg, i.p.) 30 min before the surgery to prevent noradrenergic neuron damage and received unilateral injections of 6-OHDA (6 μg/3 μL saline containing 0.02% ascorbic acid, at the injection speed of 1 μ/min) into the left side of the striatum at the following coordinates: anteroposterior, +1.2 mm; median lateral, −1.8 mm; and dorsoventral, −3.6 mm. Mice were left on a warming pad until they awoke from anesthesia and were returned to their home cage until use. To avoid dehydration, lesioned mice received sterile glucose-saline solution immediately after surgery and once a day for 3 days.
Immunohistochemistry
Immunohistochemistry was conducted as described previously. 22 Mice were transcardially perfused with saline followed by 4% paraformaldehyde/PBS. The perfused brains were dissected, postfixed overnight, and then cut into 40 μm coronal sections on a vibratome (Leica, Wetzlar, Germany). Free-floating sections were blocked with serum for 1 h and incubated overnight at 4°C with the primary antibodies. The primary antibodies were rabbit polyclonal antibodies for TH (Pel-Freez) and Iba-1 (ionized calcium-binding adapter molecule 1; Wako, Richmond, VA, USA). Immunohistochemistry was then performed using biotinylated secondary anti-rabbit IgG (Vector Laboratories, Burlingame, CA, USA), avidin-biotinylated peroxidase complex (Vector Laboratories), and 3,3′-diaminobenzidine (Sigma-Aldrich Co.). TH-stained neurons in the left and right SNc (−3.6 to −3.0 mm from the bregma) were counted for two sections per animal. Images were taken from two optical planes per section at 200 × magnification (10 × ocular and 20 × objective; Olympus Corporation, Tokyo, Japan). To avoid double counting of neurons with unusual shapes, TH-stained cells were counted only when their nuclei were visualized in a focal plane in SNc. Qualitative evaluations of immunoreactive cells were performed in a blinded manner.
D-amphetamine-induced rotational behavior test
D-amphetamine (D-AMPH) (5 mg/kg) was used to measure the drug-induced asymmetric rotational behavior after the 6-OHDA injection. Rotational behaviors were recorded in an observation cylinder (diameter, 20 cm; height, 13 cm). The number of rotations was analyzed by a SMART video tracking program.
Measurements of the MDA level
The malondialdehyde (MDA) assay was performed using the BIOXYTECH MDA-586 kit (Oxis International Inc., OR, USA), according to the manufacturer's instruments. Briefly, 6-OHDA-lesioned striatum was homogenized in ice-cold PBS containing 5 mM butylated hydroxytoluene. After centrifugation, the supernatant was mixed with probucol, diluted R1 reagent, and 12 N hydrogen chloride. The supernatant was measured at 586 nm. The protein concentration of the supernatant was determined by the Bradford method. MDA data were normalized with respect to the protein concentration and expressed as a percentage of the vehicle-treated control value.
Statistical analysis
Two-sample comparisons were carried out using Student's t-test, while multiple comparisons were made using one-way ANOVA followed by the Tukey–Kramer post hoc test. All data are presented as the mean ± standard error of the mean and statistical differences were accepted at the 5% level unless otherwise indicated.
Results
HJM shows neuroprotective effect on SH-SY5Y cells
The cytotoxic effect of various concentrations of HJM was evaluated 24 h after the treatment. Although showing slightly reduced cell viability at 200 μg/mL, HJM was not cytotoxic at any concentration examined (Fig. 1A). Treatment with 50 μM 6-OHDA for 24 h reduced cell viability compared to the control group, while pretreatment with HJM prevented the neuronal damage caused by 6-OHDA stress at concentrations of 50, 100, and 200 μg/mL compared to the group treated with 6-OHDA only (Fig. 1B).
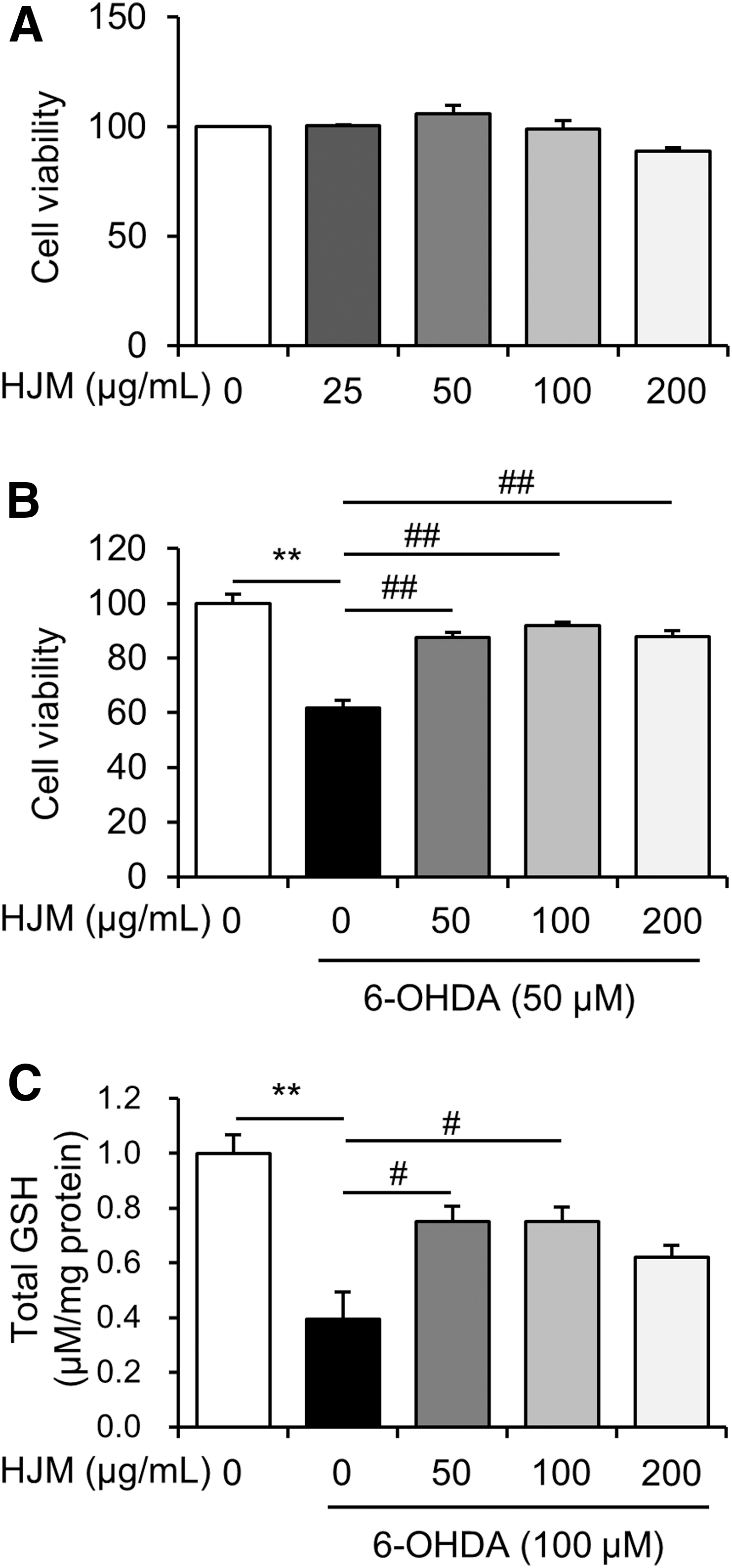
Effect of HJM on 6-OHDA-induced cytotoxicity in SH-SY5Y cells. Cells were incubated with HJM alone at the indicated concentrations for 24 h and cell viability was analyzed by MTT assay
HJM inhibits 6-OHDA-induced GSH decrease in SH-SY5Y cells
GSH is an important antioxidant for preventing damage to important cellular components caused by ROS in cells. Treatment with 50 μM 6-OHDA significantly decreased GSH levels to 39.38% ± 0.10 of control values, respectively (Fig. 1C), in the cells. However, pretreatment with 50 and 100 μg/mL HJM significantly increased the GSH level to 75.16% ± 0.05 and 74.93% ± 0.05 of control values.
HJM exhibits antiapoptotic effects in SH-SY5Y cells
Based on the neuroprotective effect of HJM in SH-SY5Y cells, we next determined the effect of HJM on apoptosis signaling. Treatment with 50 μM 6-OHDA increased expression levels of cleaved caspase-9, cleaved caspase-3, and cleaved PARP protein in the cells. The increased expression of cleaved caspase-9 and cleaved caspase-3 was significantly decreased by pretreatment with HJM at 100 and 200 μg/mL (Fig. 2B, C). Increases in expression levels of cleaved PARP were significantly inhibited by pretreatment with HJM at 50, 100, and 200 μg/mL (Fig. 2C). 6-OHDA treatment resulted in a significant increase of cytosolic cytochrome c protein expression in SH-SY5Y cells. Cytochrome c release from the mitochondria to the cytosol was significantly suppressed by pretreatment with HJM (Fig. 2E, F).
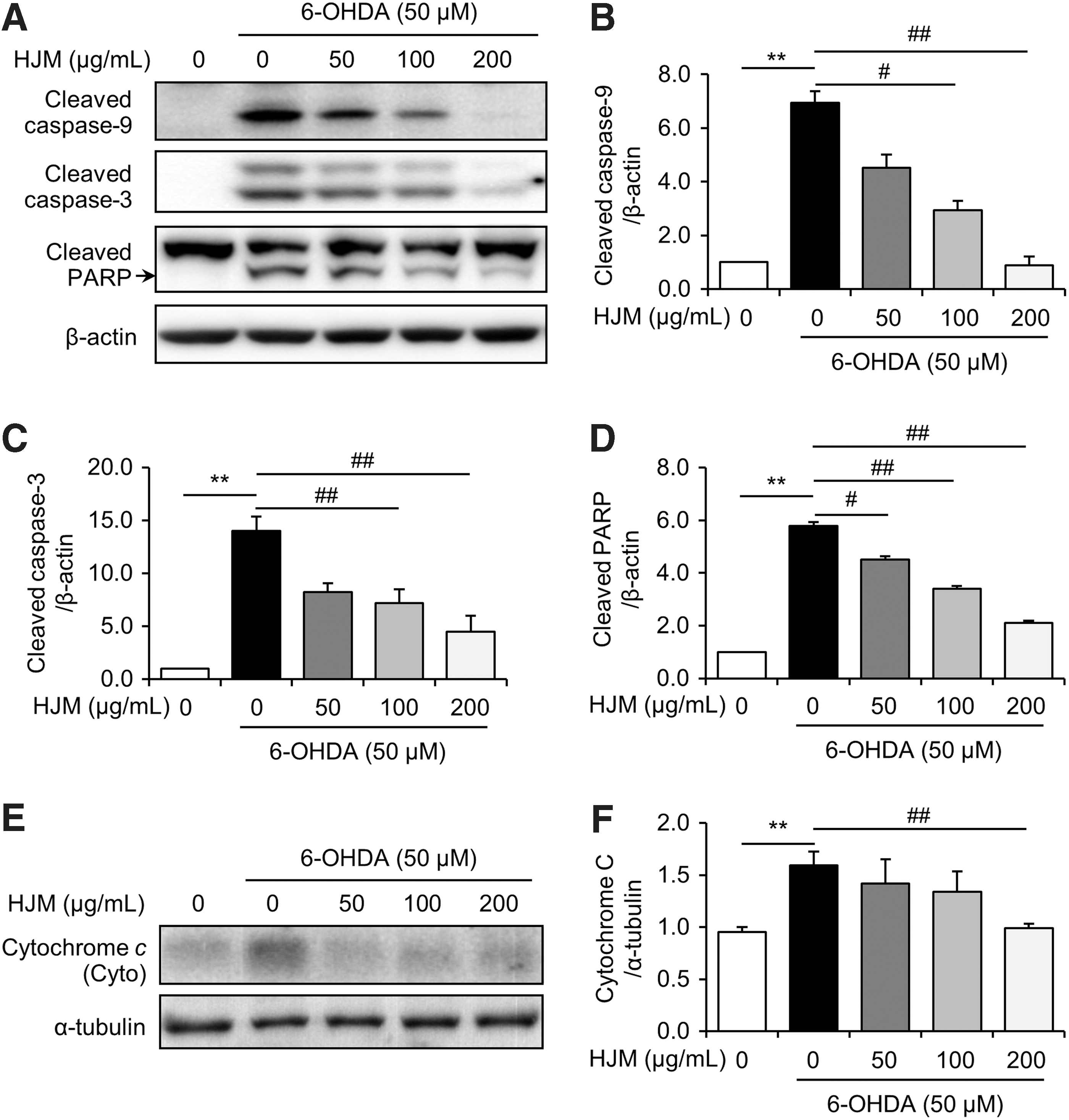
Effect of HJM on the 6-OHDA-induced apoptosis in SH-SY5Y cells. Cells were pretreated with the indicated concentrations of HJM for 1 h and then exposed to 50 μM 6-OHDA for 24 h. Levels of caspase 9, caspase 3, PARP, cytochrome c, β-actin, and α-tubulin were evaluated by Western blot analysis. Cleaved caspase-9, cleaved caspase-3, cleaved PARP, and cytosolic cytochrome c immunoreactivity
HJM inhibits 6-OHDA-induced phosphorylation of ERK1/2 in SH-SY5Y cells
To further explore the mechanism underlying the antiapoptotic properties of HJM, we determined the effect of HJM on the modulation of MAPK proteins. Treatment with 50 μM 6-OHDA increased phosphorylation of ERK1/2, p38, and JNK to 156.97% ± 0.12, 237.32% ± 0.29, and 130.97% ± 0.09 of control values, respectively. However, pretreatment with 200 μg/mL HJM significantly decreased phosphorylation of ERK 1/2 (Fig. 3B). The phosphorylation levels of p38 and JNK appeared not to be affected (Fig. 3C, D).
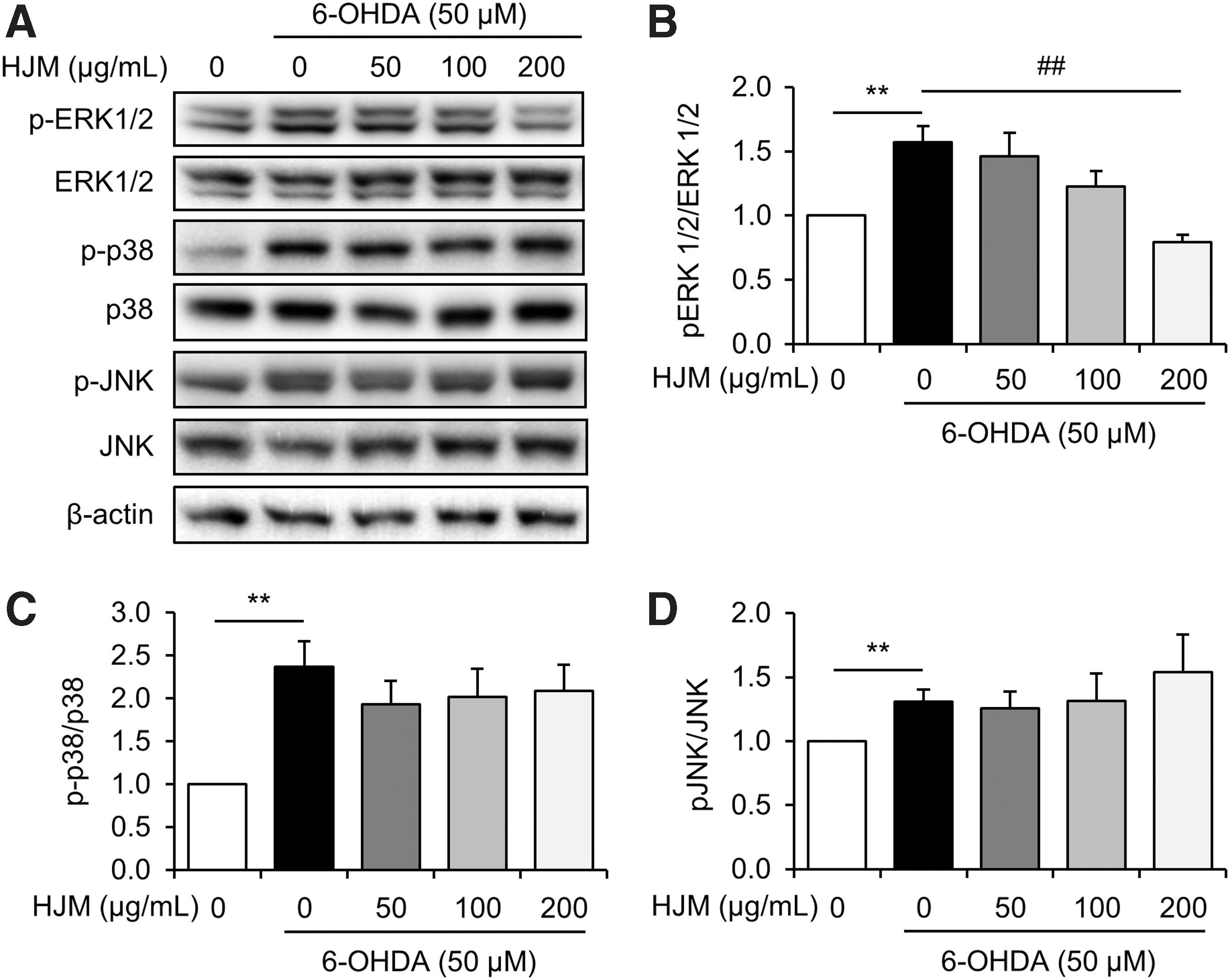
Effect of HJM on MAPK modulation in 6-OHDA-stimulated SH-SY5Y cells. Cells were pretreated with the indicated concentrations of HJM for 1 h and then exposed to 50 μM 6-OHDA for 24 h. Levels of ERK 1/2, p38, and JNK were evaluated by Western blot analysis. **P < .01 and ## P < .01 as analyzed by Student's t-test. ERK1/2, extracellular signal-regulated kinase 1/2; JNK, c-Jun N-terminal kinase.
HJ improves asymmetric motor behavior in a 6-OHDA mouse model
To determine the effects of HJ on PD, a mouse model with unilateral 6-OHDA lesion was used. The unilateral dopaminergic neuronal death induced by 6-OHDA in rodents leads to asymmetric rotational behavior, and the rotational behavior can be elicited by drugs that increase brain dopamine activity. At 8 days after 6-OHDA injection, circling behavior tests were performed on vehicle-treated mice, 500 mg/kg HJM-treated mice, 300 mg/kg HJE-treated mice, and 500 mg/kg HJE-treated mice. D-AMPH-induced ipsilateral rotations were significantly decreased in the 500 mg/kg HJM- and 500 mg/kg HJE-treated groups compared to the vehicle-treated group (Fig. 4A).
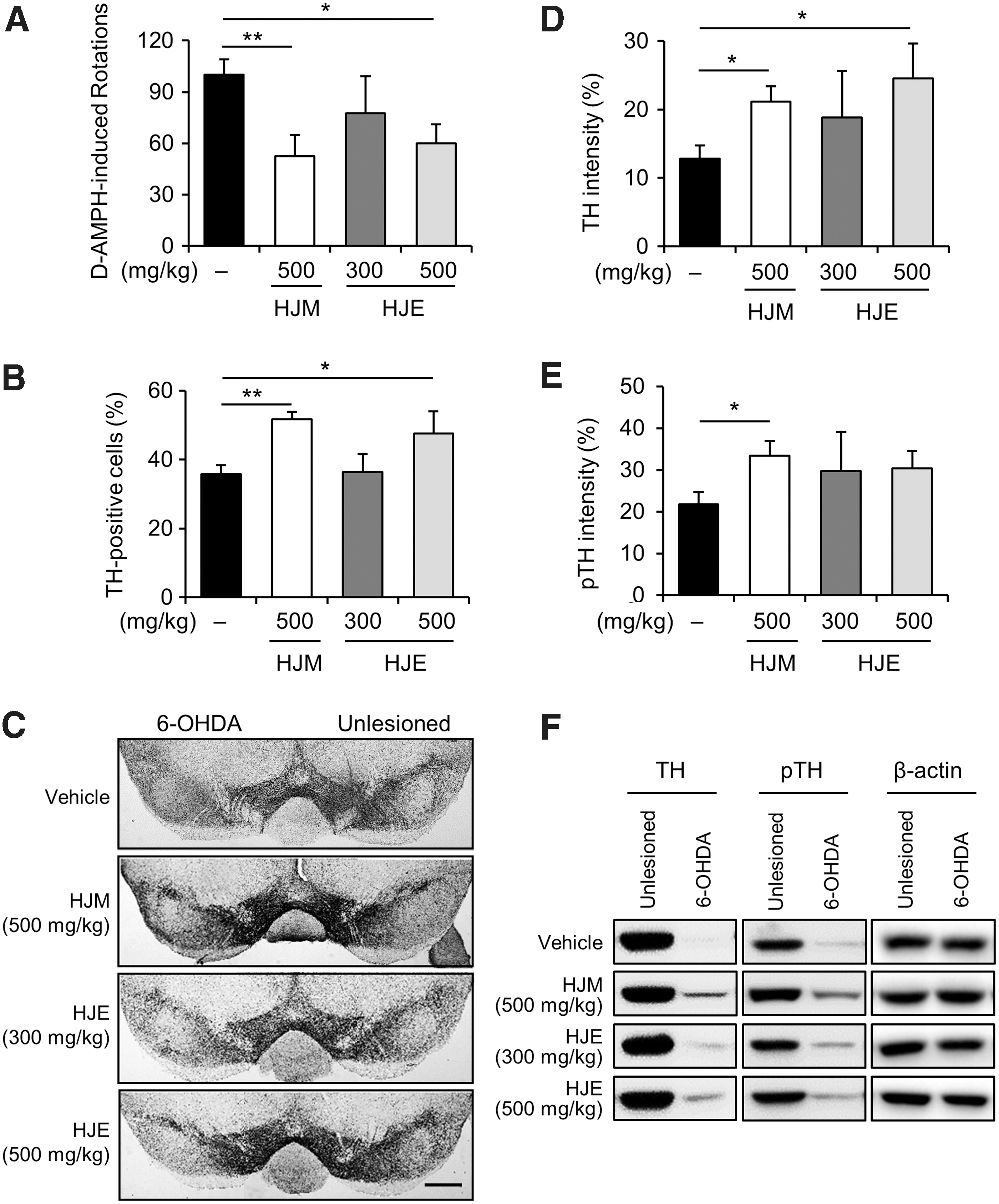
Effect of HJM and HJE on D-AMPH-induced circling behavior and dopaminergic cell death in 6-OHDA-lesioned mice. D-AMPH-induced rotational behavior was tested at 8 days after 6-OHDA lesion. Sum of total ipsilateral rotations recorded during 60 min (
HJ shows neuroprotective effects in a 6-OHDA mouse model
The number of TH-positive cells on the 6-OHDA-lesioned side was analyzed as a percentage of those on the intact side in the SNc (Fig. 4B, C). The percentages of TH-positive cells in the ipsilateral SNc regions in the vehicle-treated group, 500 mg/kg HJM-treated group, 300 mg/kg HJE-treated group, and 500 mg/kg HJE-treated group were 35.77% ±2.57, 51.71% ± 2.16, 36.44% ± 5.16, and 47.56% ± 6.49 (Fig. 4B), respectively. The number of TH-positive cells in the lesioned SNc was significantly increased in the 500 mg/kg HJM-treated group and 500 mg/kg HJE-treated group compared to the vehicle-treated group. Moreover, the percentages of TH-immunoreactivity in the lesioned striatum in the vehicle-treated group, 500 mg/kg HJM-treated group, 300 mg/kg HJE-treated group, and 500 mg/kg HJE-treated group were 12.81% ± 1.91, 21.17% ± 2.21, 18.79% ±6.82, and 24.52% ± 5.11 (Fig. 4D), respectively. The percentages of pTH-immunoreactivity in the lesioned striatum in the vehicle-treated group, 500 mg/kg HJM-treated group, 300 mg/kg HJE-treated group, and 500 mg/kg HJE-treated group were 21.74% ± 2.94, 33.31% ± 3.58, 29.79% ± 9.29, and 30.32% ± 4.24 (Fig. 4E), respectively. The loss of TH-positive fibers in the lesioned striatum was significantly decreased in the 500 mg/kg HJM-treated group and 500 mg/kg HJE-treated group compared to the vehicle-treated group. These data indicated that HJM and HJE had a neuroprotective effect in the 6-OHDA mouse PD model.
HJM exhibits anti-inflammatory and antioxidative effects in a 6-OHDA mouse model
To determine the anti-inflammatory potential of HJM in the 6-OHDA mouse model, the number of reactive microglia was analyzed using immunohistochemistry in the 6-OHDA-lesioned SNc of mice. The number of Iba-1-positive cells, a marker of activated microglia, was significantly increased by 6-OHDA injection in both groups (Fig. 5A, B). However, the number of activated microglia was significantly decreased in the 6-OHDA-lesioned SNc of HJM-treated mice compared to vehicle-treated mice (Fig. 5A, B) suggesting that HJM showed an anti-inflammatory response in vivo. Moreover, as one of the markers of oxidative stress, the lipid peroxidation product MDA was detected to assess the oxidative stress in 6-OHDA-lesioned side of striatum (Fig. 5C). The MDA value was significantly decreased in the HJM-treated group compared to the vehicle-treated group, suggesting that HJM showed an anti-oxidative response in vivo.
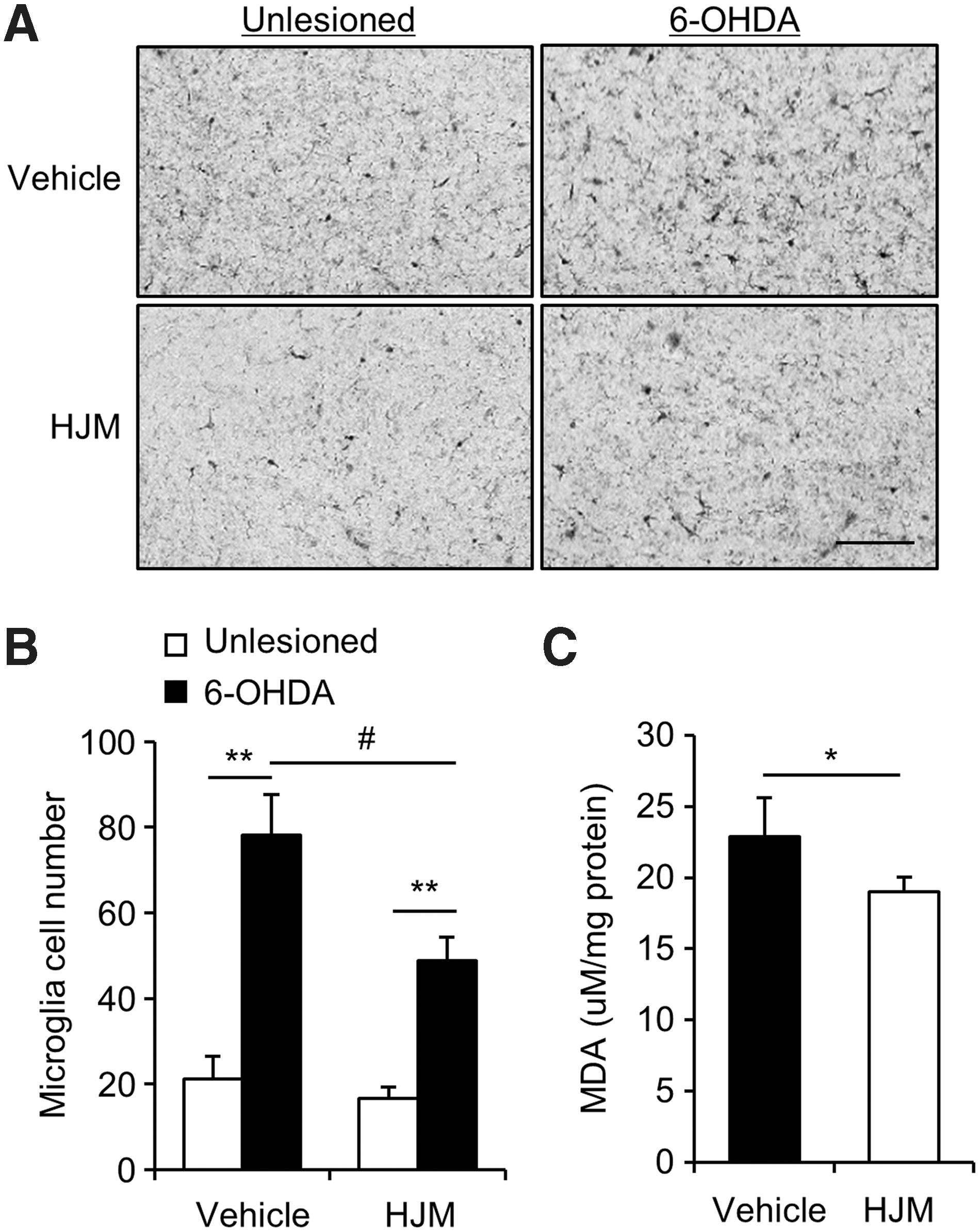
Effects of HJM on inflammation and oxidative stress in 6-OHDA-lesioned mice. Ten days after the 6-OHDA injection, immunohistochemical analysis was performed in vehicle-treated (vehicle) and HJM-treated (HJM) mice. Photomicrograph
Discussion
The present study demonstrated the protective effects of methanol and ethanol extracts of HJ against 6-OHDA-induced neurotoxicity in SH-SY5Y cells and in a mouse model of PD. The HJ effect appeared to be associated with the regulation of oxidative stress, pERK1/2 regulation, mitochondrial-mediated apoptosis, and inflammation.
In this study, HJM significantly increased GSH levels in SH-SY5Y cells exposed to 6-OHDA and decreased lipid peroxidation in 6-OHDA-lesioned mice (Figs. 1 and 5). ROS production, detoxification, and signaling have been interesting targets for intervention in neurodegenerative diseases, including PD. 2,9 Endogenous antioxidants, such as GSH, thioredoxin/thioredoxin reductase/peroxiredoxin, and catalase, delay or prevent oxidative damage to proteins, lipids, and DNA. 23 However, decreased GSH levels, increased ROS production, and increased mitochondrial dysfunction have been reported in brain tissue of patients with PD. 9,24 Thus, the initial blockade of intracellular generation of ROS caused by stresses might be an important strategy for protecting neural cells in PD. In addition, mitochondrial dysfunction caused by oxidative stress and cytochrome c release from damaged mitochondria triggers activation of cleaved caspase-3 and PARP through upregulation of cleaved caspase-9. 25,26 The HJM pretreatment inhibited the activation of the mitochondrial apoptosis pathway caused by 6-OHDA in SH-SY5Y cells (Fig. 2), suggesting that HJM might be potentially beneficial for protecting neural cells in PD.
MAPK signaling cascades in neurodegeneration are important for the translation of oxidative stimuli into cellular responses such as apoptosis. 27 –29 Thus, the modulation of these signal pathways has been reported as a potential strategy for the therapeutic interventions of neurodegenerative diseases. 10 –12 In this study, ERK1/2 activation and cytotoxicity induced by 6-OHDA were significantly reduced by pretreatment of HJM in SH-SY5Y cells (Figs. 1 and 3), but p38 and JNK activation was not affected. The phosphorylation of ERK1/2 was enhanced in the postmortem brain of PD patients, 29 and the inhibition of ERK1/2 activation has been shown to protect against neuronal cell death from oxidative stress-induced neurotoxicity. 30,31
Besides antioxidative and antiapoptotic effects, HJM exhibited inhibited inflammation in 6-OHDA-lesioned SNc and striatum (Fig. 5). Neuroinflammation, as well as oxidative stress, is a potential contributor of PD. 32 The chronic activation of microglia in PD may cause neuronal damage through the release of various cytotoxic molecules such as proinflammatory cytokines and ROS. 33 HJ has shown strong anti-inflammation effects in LPS-treated BV2 cells, an Alzheimer's disease mouse model. 19 In this study, increased microglia activation caused by 6-OHDA was significantly decreased by treatment of HJM in lesioned SNc (Fig. 5).
Previously, it was reported that methanol extract of HJ contained various antioxidant polyphenols and flavonoids such as luteolin, luteolin 7-glucoside, quercetin, and apigenin. 17,20,34 Especially, luteolin and apigenin exhibit strong antioxidant and anti-inflammatory efficacy 35,36 and have been suggested as potential compounds for prevention and treatment of neurodegenerative disorders. 37,38 In this study, we demonstrated that both methanol extract and ethanol extract of HJ showed neuroprotective effect against 6-OHDA mouse model (Fig. 4). The loss of TH-positive cells in the lesioned SNc and TH-fibers in the lesioned striatum was significantly decreased in the 500 mg/kg HJM-treated and 500 mg/kg HJE-treated group compared to the vehicle-treated group and both HJM and HJE groups improved asymmetric motor behavior in a 6-OHDA mouse model. Based on these results, further studies are needed to identify bioactive compounds on PD and optimize the extraction of the bioactive compounds using the methanol and ethanol extracts of HJ.
The results of this study demonstrated that pretreatment with HJ protected against the PD pathology in the 6-OHDA model. HJ markedly attenuated 6-OHDA-induced cell death in SH-SY5Y cells, through inhibition of intracellular ROS accumulation by decreasing of an antioxidant molecule, GSH, as well as activation of ERK1/2. In addition, the protective effects of HJM against 6-OHDA-induced mitochondrial apoptosis were associated with inhibition of release of cytochrome c release and activation of cleaved caspase-9, cleaved caspase-3, and cleaved PARP. Moreover, the regulation of inflammation by HJ might mediate the protective effects of HJ in the PD animal model. As the potential of HJ in PD therapy is investigated, further studies will be required to define whether benefits would be maintained with long-term treatment and whether there is a beneficial effect of HJ on patients with PD.
Footnotes
Acknowledgments
This work was supported by the KRIBB Research Initiative Program of the Republic of Korea, and by the Development of Platform Technology for Innovative Medical Measurements Program from Korea Research Institute of Standards and Science (KRISS-2016-16011064).
Author Disclosure Statement
No competing financial interests exist.