
Abstract
Asthma is a chronic inflammatory disease of bronchial airway, which is characterized by chronic airway inflammation, airway edema, goblet cell hyperplasia, the aberrant production of the Th2 cytokines, and eosinophil infiltration in the lungs. In this study, the therapeutic effect and the underlying mechanism of Citrus tachibana leaves ethanol extract (CTLE) in the ovalbumin (OVA)-induced allergic asthma and compound 48/80-induced anaphylaxis were investigated. Oral administration of CTLE inhibited OVA-induced asthmatic response by reducing airway inflammation, OVA-specific IgE and IgG1 levels, and increasing OVA-specific IgG2a levels. CTLE restored Th1/Th2 balance through an increase in Th2 cytokines tumor necrosis factor-α, interleukin (IL)-4, and IL-6 and decreases in Th1 cytokines interferon-γ and IL-12. Furthermore, CTLE inhibited the total level of NF-κB and the phosphorylation of IκB-α and NF-κB by OVA. In addition, CTLE dose-dependently inhibited compound 48/80-induced anaphylaxis via blocking histamine secretion from mast cells. The anti-inflammatory mechanism of CTLE may involve the modulation of Th1/Th2 imbalance via inhibiting the NF-κB signaling and histamine secretion. Taken together, we suggest that CTLE could be used as a therapeutic agent for patients with Th2-mediated or histamine-mediated allergic asthma.
Introduction
A
Citrus fruits are one of the most important dietary sources of bioactive compounds 14 and have received attention not only for their nutritional properties but also for their medicinal value. 15,16 Citrus fruits are rich in vitamin C and several bioactive compounds such as coumarins, carotenoids, and flavonoid, 15,16 which were found to have potent antioxidant, anti-inflammatory, anticholinesterase, radical-scavenging, and hypoglycemic activities. 15 –18 In addition, methanol extract of Citrus aurantifolia leaves exhibit chemical variability, radical scavenging, and total antioxidant capacity. 18 Especially, Citrus tachibana leaf extract has been reported to mitigate ovalbumin (OVA)-induced food allergy symptoms including increased rectal temperature, diarrhea, and anaphylaxis by decreasing cytokine release form Th2 cells in mesentery lymph nodes of mice. 19 However, the protective effects of C. tachibana on compound 48/80-induced systemic anaphylactic shock and OVA-induced asthma have not been revealed. In this study, we investigated the protective effects of C. tachibana leaves ethanol extract (CTLE) on the OVA-induced murine asthma model to evaluate the therapeutic efficacy of CTLE in allergic asthma via the suppression of Th2 cytokine production from Th2 cells.
Materials and Methods
Animals
Male SPF Balb/c and ICR mice aged 6 weeks were purchased from Damool Science (Daejeon, Korea) and housed in the clean barrier animal facilities under standard 12 h light/dark cycles and allowed free access to food and water. All animal experiments were performed in accordance with the Guidelines for Animal Care and Use and were approved by the Institutional Animal Care and Use Committee of Chonbuk National University Laboratory Animal Center (CBN 2013-0011, CBN 2016-37).
Preparation of CTLE
Korea Food Research Institute provided CTLE extracted by the method described previously. 19 In brief, its leaves were ground under reflux and extracted twice in 95% ethanol using a Soxwave100 apparatus. The extract solution was dried under a vacuum in a rotary evaporator. The dried ethanol extract powders were kept at −80°C, and dissolved and freshly diluted in saline before use.
Induction of systemic anaphylaxis reaction
Systemic anaphylaxis reaction was investigated by the method described previously. 20 ICR mice intraperitoneally received 8 mg/kg body weight (BW) of mast cell degranulator compound 48/80 or saline. CTLE (100, 200, and 400 mg/kg BW) and dexamethasone (Dex, 2.5 mg/kg BW) were dissolved in saline and administered orally during 6 days before the injection of compound 48/80 (n = 7/group). Mortality was monitored for 1 h after induction of systemic anaphylactic shock. After the mortality test, mesentery was obtained from intestines of each mouse, which was then stained with toluidine blue solution and the rate of mast cell degranulation was calculated by the method previously described. 21 The mast cells were classified ( × 1000) as follows: (1) severely degranulated (>50% of the cytoplasmic granules exhibiting fusion, staining alterations, and extrusion from the cell), (2) slight to moderately degranulated (10–50% of the granules exhibiting fusion or discharge), or (3) normal.
Induction of OVA-induced asthma model
An asthma murine model induced by OVA was established as previously described with some modification. 22 Mice were divided into four groups according to treatment: (1) Saline as a vehicle control, (2) OVA-induced asthmatic mice, (3) 200 mg/kg/day CTLE in OVA-induced asthmatic mice, and (4) 2.5 mg/kg/day Dex in OVA-induced asthmatic mice as a reference drug for positive control. Mice of groups (2), (3), and (4) were immunized by intraperitoneal injection of 50 μg OVA with 1 mg Imject Alum in a total volume of 200 μL on days 1 and by intraperitoneal injection of 50 μg OVA without Alum on 14. On days 27, 28, and 29 after the beginning of the sensitization, these mice were challenged for 30 min with an aerosol of 5% (wt/vol) OVA in saline using ultrasonic nebulization (NE-U12; Omron Corp., Tokyo, Japan). On days 15 to 26, the treatment groups were also orally treated once daily with 200 mg/kg CTLE or 2.5 mg/kg Dex. Vehicle control group and OVA-induced asthma groups received only saline. Animals were sacrificed 24 h after the last challenge on day 30 to investigate the inhibitory effect of CTLE.
Collection of bronchoalveolar lavage fluid and cell count
Twenty-four hours after the final OVA challenge, bronchoalveolar lavage fluid (BALF) was collected by cannulating the upper part of the trachea and lavaging, as described previously. 22 The total number of viable cells in BAL fluid was determined by trypan blue exclusion using a hemocytometer. Differential cell counts were determined using cytosine (Centrifuge 5403; Eppendorf, Hamburg, Germany) preparations, followed by Diff Quik staining (Sysmex Co., Kobe, Japan).
Histopathological examination of lung tissues
Histopathologic analysis of lung was performed as previously described. 22 Animals were sacrificed with an overdose of ether 24 h after the last OVA challenge and histologic specimens were collected. Tissue specimens were taken from the mid zone of the lung of the mice, and fixed in 10% formalin and embedded in paraffin. Serial 5 μm thickness sections were stained with hematoxylin and eosin (H&E), periodic acid-Schiff, and Masson trichrome.
Measurement of Th1/Th2 cytokines, total NF-κB, p-NF-κB, and p-IκBα levels in BALF and OVA-specific IgE, IgG1, and IgG2a in serum
Levels of Th1-related cytokines such as interferon-γ (IFN-γ) and IL-12 and Th2-related cytokines such as tumor necrosis factor-α (TNF-α), IL-4, IL-5, and IL-10, and total nuclear factor (NF)-κB, phospho-NF-κB (p-NF-κB), and phospho-inhibitor of κB (p-IκB) in the BALF from each mouse were measured using enzyme-linked immunosorbent assay (ELISA) as described previously. 23 And total immunoglobulin (Ig) E and OVA-specific IgE, IgG1, and IgG2a in serum were measured using ELISA kits according to the manufacturer's instructions.
Statistical analysis
Results were expressed as mean ± standard deviation for the number of experiments. Student's t-test and ANOVA with Dunnett's test were used for statistical comparison among the groups. Results with P < .05 were considered statistically significant.
Results
Effect of CTLE on compound 48/80-induced systemic anaphylaxis
Compound 48/80 is a basic amine polymer and one of the powerful stimulators of mast cell degranulation and, therefore, is useful in anaphylaxis studies. 24 To investigate the effect of CTLE in anaphylactic shock, we first used the in vivo model of systemic anaphylaxis using compound 48/80. As shown in Figure 1A, the intraperitoneal injection of compound 48/80 (8 mg/kg BW) resulted in 85.71% death of mice. However, oral administration of CTLE (100–400 mg/kg BW) reduced compound 48/80-induced mortality rate in a dose-dependent manner (Fig. 1A). Also, dexamethasone (2.5 mg/kg BW) significantly inhibited the mortality rate induced by compound 48/80 (Fig. 1A). The effect of CTLE on compound 48/80-induced mesentery mast cell degranulation in mice was also investigated. As shown in Figure 1B–D, the oral treatment of CTLE significantly inhibited the compound 48/80-induced mesentery mast cell degranulation and histamine secretion, and the inhibition rate by CTLE was showed in a dose-dependent manner (Fig. 1B–D). In addition, dexamethasone significantly decreased mesentery mast cell degranulation induced by compound 48/80. These results suggest that CTLE effectively block compound 48/80-induced anaphylaxis via the inhibition of mast cell degranulation and histamine release.
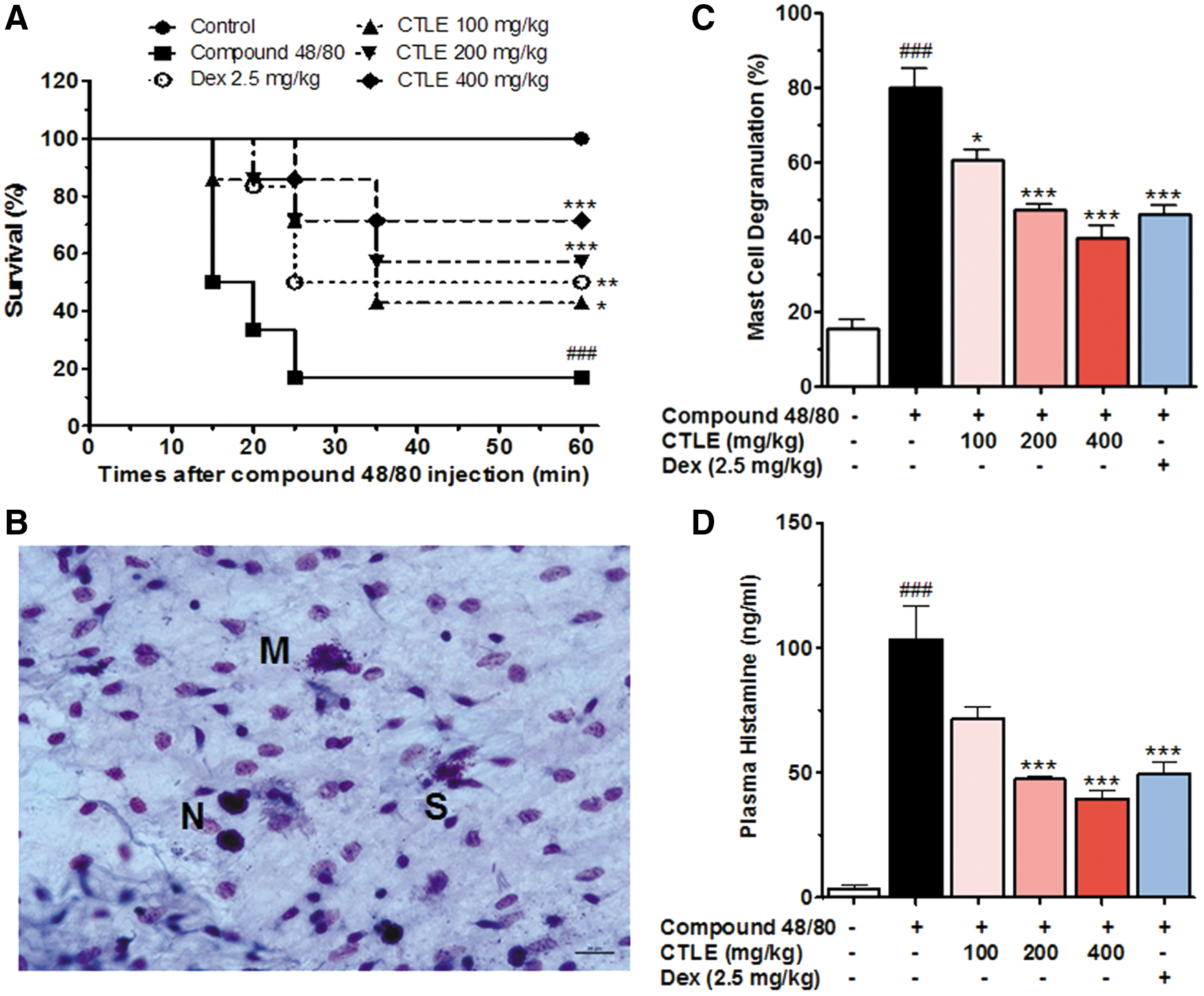
Effect of CTLE on compound 48/80-induced anaphylaxis. Mice were orally administered with CTLE, Dex, or saline as a vehicle (n = 7 per group) for 6 days. The compound 48/80 (8 mg/kg) was intraperitoneally injected to mice 1 h after the last administrations of CTLE, Dex, or saline.
Effect of CTLE on OVA-induced allergic asthma
As shown in Figure 2, the number of total BALF cells and inflammatory cells in BALF from OVA-induced asthmatic mice was significantly elevated compared to that of saline-treated mice (Fig. 2B–C). Oral administration of 200 mg/kg/day CTLE or 2.5 mg/kg/day Dex significantly reduced the increase of eosinophils and lymphocytes and had inflammatory cells similar to those of the mice treated with saline alone (Fig. 2D–F).
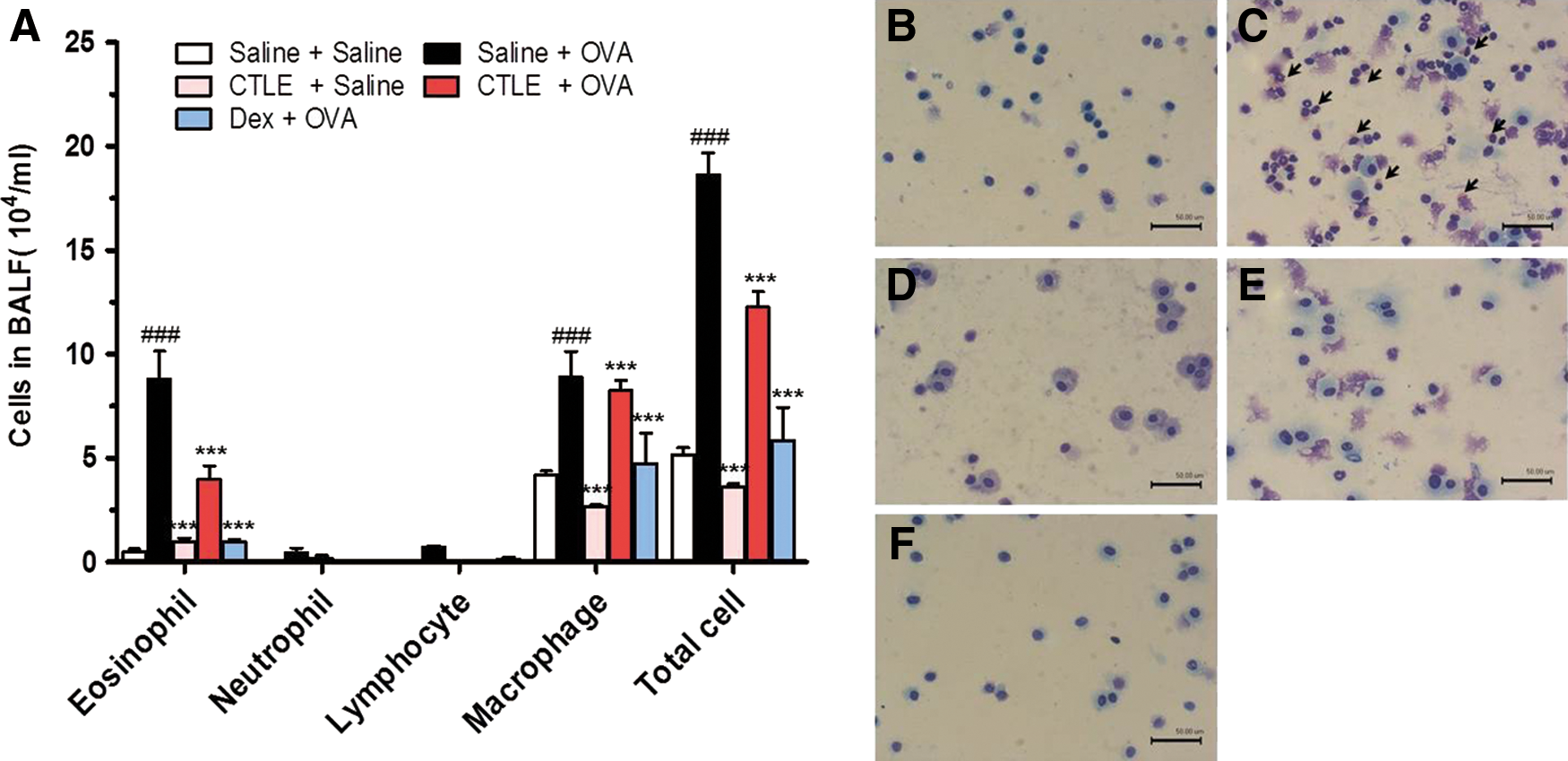
Effect of CTLE on
Because CTLE suppresses inflammatory cell influx into the BALF, the antiasthmatic effects of the CTLE were next examined. Histopathological examinations revealed typical pathological features of asthma in OVA-induced asthmatic mice. Significant infiltrations of inflammatory cells and eosinophils were observed in both perivascular and peribronchial areas of lungs from OVA-induced asthmatic mice (Fig. 3). Moreover, the lung tissues of mice treated with OVA exhibited increased accumulation of mucus in the lumen of bronchioles, and hyperplasia of goblet cells at the epithelium of bronchioles compared to saline-treated mice. In addition, collagen fiber deposits among inflammatory cells and between submucosal and muscular layers were observed in lung tissues of OVA-induced asthmatic mice (Fig. 3). However, oral treatment of either CTLE or Dex significantly decreased the infiltration of inflammatory cells, the accumulation of mucus in the lumen of bronchioles, and hyperplasia of goblet cells in the epithelium of bronchioles in lung induced by OVA. Also both CTLE and Dex treatment significantly attenuated collagen fiber deposits among inflammatory cells and between submucosal and muscular layers of lungs induced by OVA (Fig. 3). These results suggest that CTLE may improve the histopathological condition of allergic asthma.
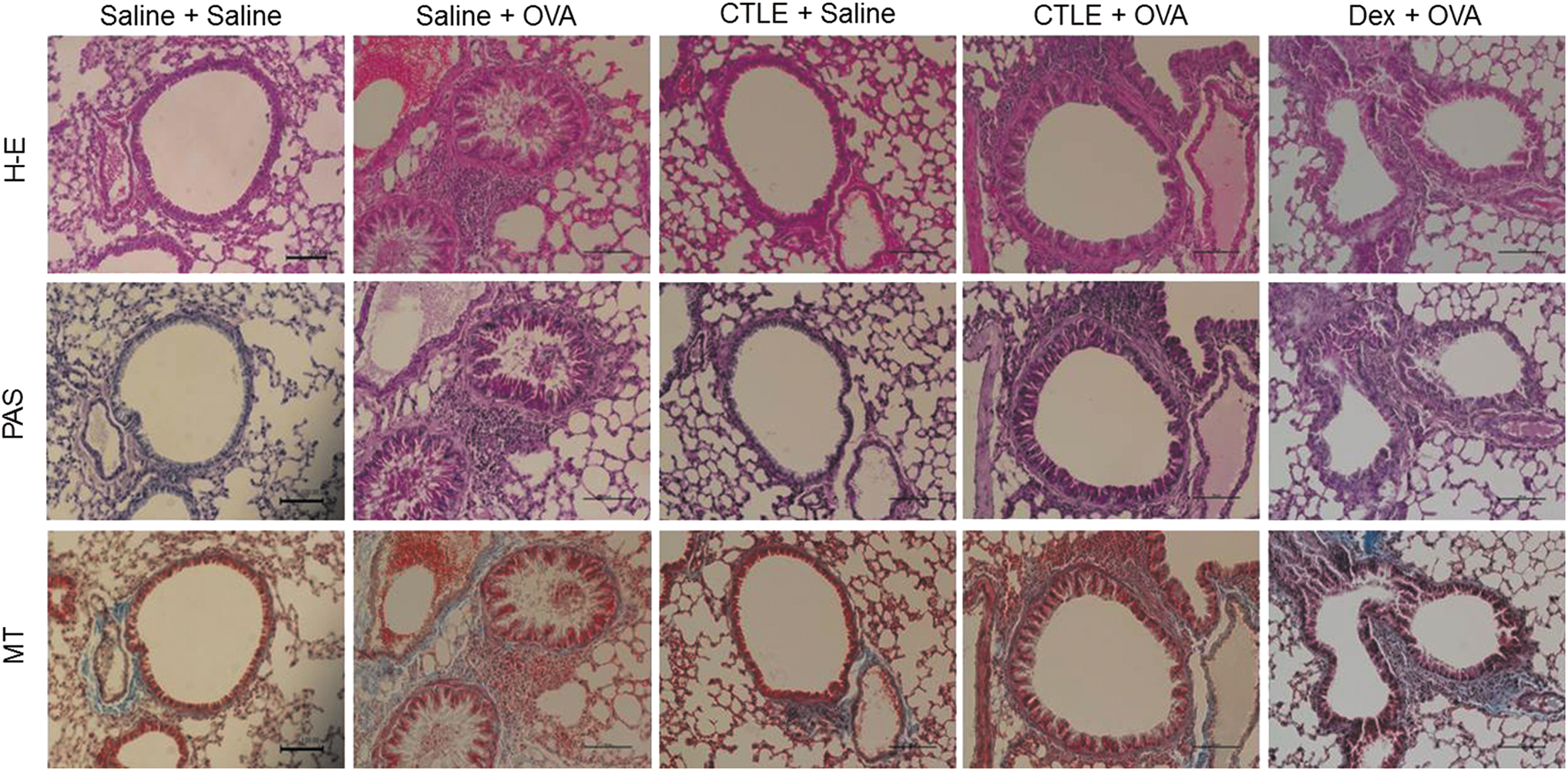
Effect of CTLE on OVA-induced histopathological changes. Mice were sensitized on days 1 and 14, and challenged on days 27, 28, and 29 by OVA. On days 15 to 26, the treatment groups were also orally treated once daily with 200 mg/kg CTLE or 2.5 mg/kg Dex. Lung tissues from each group were stained with H&E for inflammatory cells including eosinophils, PAS for goblet cells and mucus and MT for collagen fiber deposits. Scale bar = 100 μm. H&E, hematoxylin and eosin; MT, Masson trichrome; PAS, periodic acid-Schiff. Color images available online at
Effect of CTLE on OVA-induced Th2- and Th1-related cytokine levels
The imbalance of Th1 and Th2 cytokines has been known to be responsible for the pathogenesis of asthma. 25 Th2-related cytokines including TNF-α, IL-4, IL-5, and IL-10 and Th1-related cytokines including IFN-γ and IL-12 have been known to regulate IgE-mediated allergy and asthma were investigated in the BALF of OVA-induced asthmatic mice using ELISA. As shown in Figure 4, the levels of TNF-α, IL-4, IL-5, and IL-10 in BALF of OVA-induced asthmatic mice were significantly increased compared with those of mice treated with saline alone, whereas the levels of IFN-γ and IL-12 in BALF of OVA-induced asthmatic mice were greatly reduced. However, CTLE treatment significantly reduced the levels of TNF-α, IL-4, IL-5, and IL-10 and increased the levels of IFN-γ and IL-12 in BALF of mice induced by OVA (Fig. 4). The reference drug used as a positive control, Dex, exhibited similar inhibitory effects on the increase of Th2 cytokines and the reduction of Th1 cytokines. These data indicated that CTLE might effectively downregulate Th2-immune response and upregulate Th1-immune response in a mouse of OVA-induced allergic asthma.
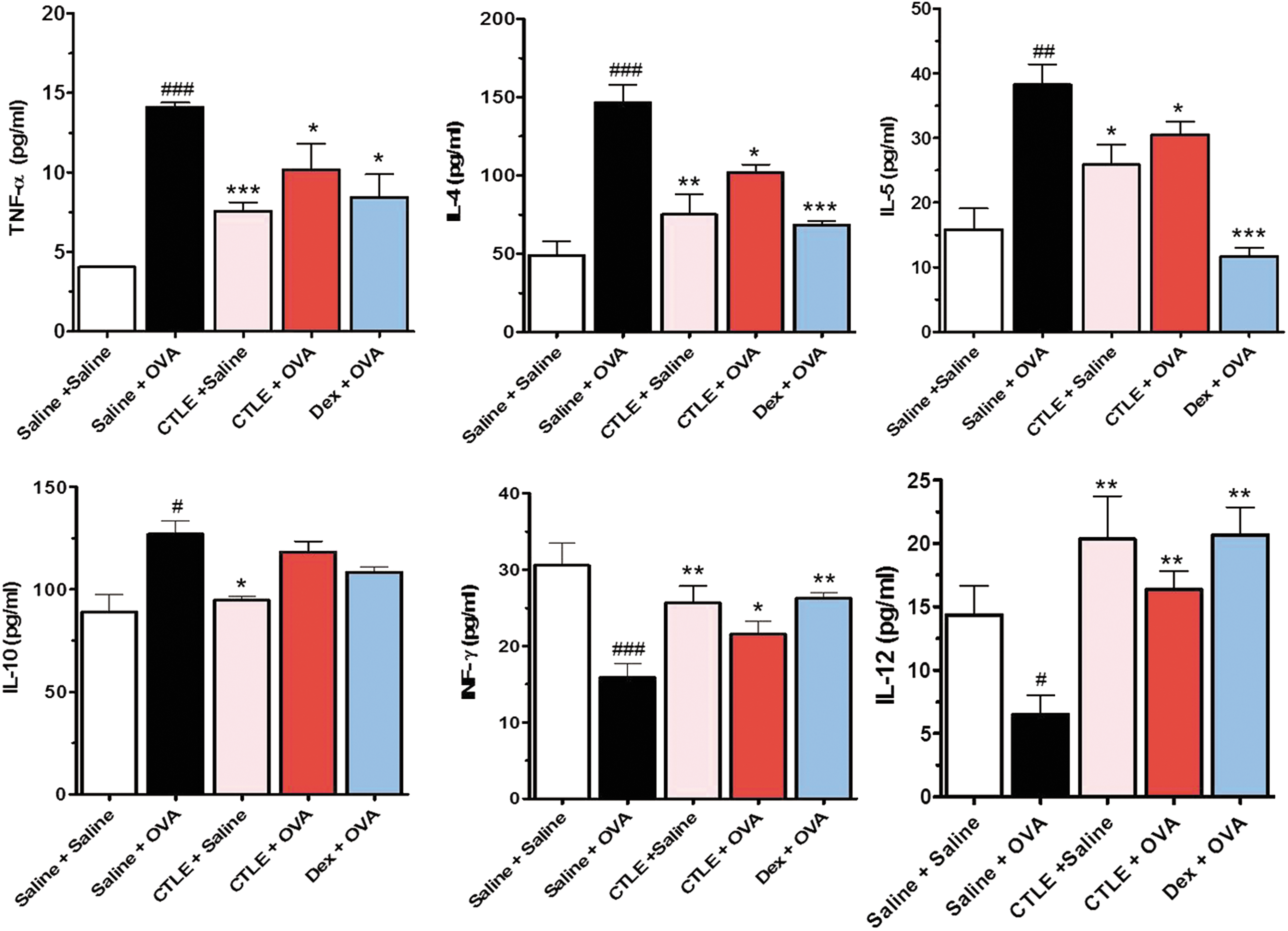
Effect of CTLE on the levels of Th1/Th2 cytokines in BALF; TNF-α, IL-4, IL-5, IL-10, IFN-γ, and IL-12. The levels of Th2 cytokines including TNF-α, IL-4, IL-5, and IL-10 in BALF of OVA-induced asthmatic mice were considerably increased, while Th1 cytokines IFN-γ and IL-12 levels were greatly reduced with those of saline-treated mice. However, CTLE significantly reduced the increased levels of Th2 cytokines and increased the decreased levels of Th1 cytokines by OVA. Data were analyzed using ANOVA followed by Student's t-test. ###
P < .001, ##
P < .01, #
P < .05, significantly different from the value of Saline group. ***P < .001, **P < .01, *P < .05, significantly different from the value of OVA group. Results are presented as the mean ± SD (n = 6 per group). IFN-γ, interferon-γ; IL, interleukin; TNF-α, tumor necrosis factor-α. Color images available online at
Effect of CTLE on OVA-induced OVA-specific IgE, IgG1, and IgG2a levels
Asthma is mainly mediated by the production and effects of IgE antibodies and other immunoglobulin classes such as IgG1 and IgG2a, which have been focus of most allergy research. 26 To investigate the underlying immunoregulatory mechanism of CTLE on the secretion of Th2- and Th1-related cytokines in BALF induced by OVA, we detected the serum levels of Th2-related immunoglobulins such as OVA-specific IgE and IgG1 and Th1-related immunoglobulin such as OVA-specific IgG2a. As shown in Figure 5, the levels of OVA-specific IgE, OVA-specific IgG1, and OVA-specific IgG2a were markedly increased in serum of OVA-challenged mice compared with saline-challenged mice. However, both CTLE and Dex significantly suppressed the levels of both the OVA-specific IgE and IgG1 in serum. In addition, CTLE tended to increase the OVA-specific IgG2a serum levels that were reduced by OVA (Fig. 5). These results showed that CTEL ameliorate allergic asthma symptoms through inhibiting secretion of OVA-specific IgE and OVA-specific IgG1 and enhancing OVA-specific IgG2a.
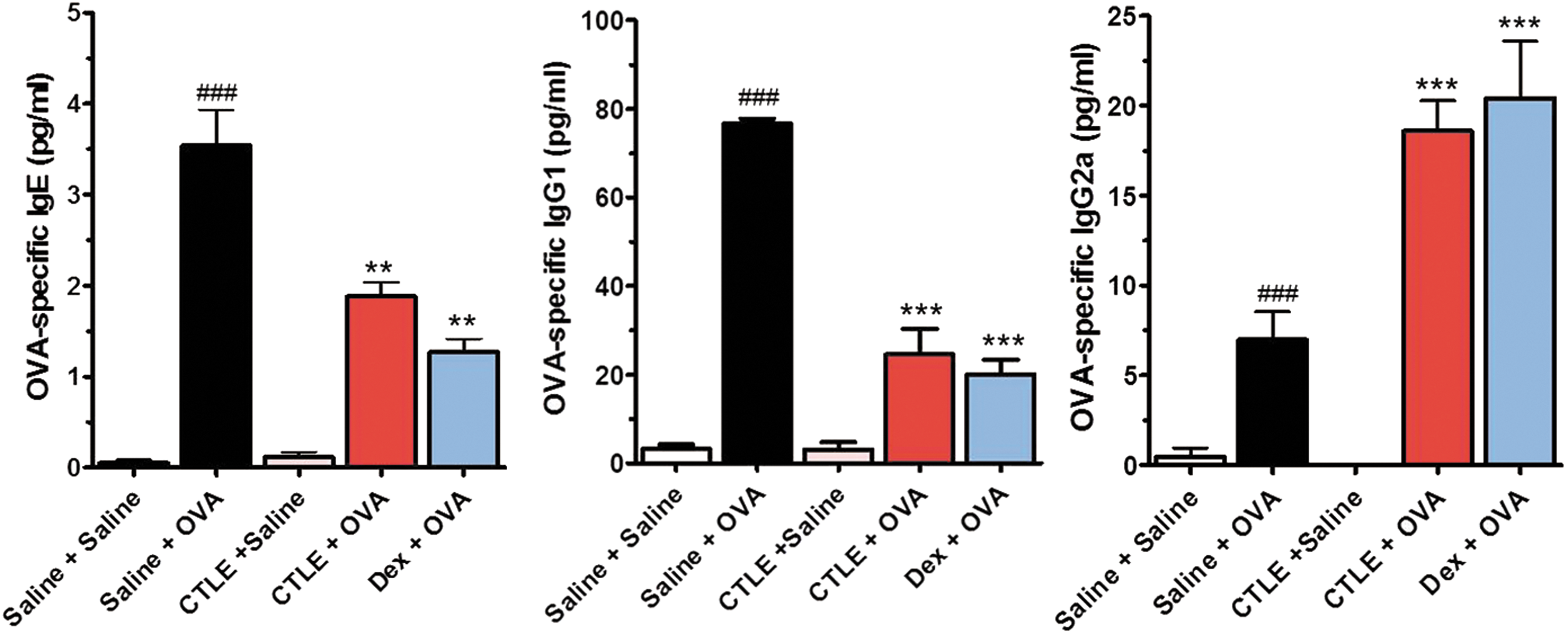
Effect of CTLE on the serum levels of OVA-specific IgE, OVA-specific IgG1, and OVA-specific IgG2. OVA-induced asthmatic mice increased the serum levels of OVA-specific IgE, OVA-specific IgG1 and OVA-specific IgG2a. CTLE notably suppressed the levels of OVA-specific IgE and OVA-specific IgG1 and tended to increase the OVA-specific IgG2a serum levels. Data were analyzed using ANOVA followed by Student's t-test. ###
P < .001, significantly different from the value of Saline group. ***P < .001, **P < .01, significantly different from the value of OVA group. Results are presented as the mean ± SD (n = 6 per group). Color images available online at
Effect of CTLE on OVA-induced NF-κB signaling activation
NF-κB, an important transcription factor, is well known to play an important role in the progression of asthma and has been reported to regulate Th2 cell differentiation and cytokine productions, such as TNF-α, IL-4, and IL-5 expression. 27 To reveal the inhibitory effect of CTLE on airway allergic asthma, NF-κB signaling activation that regulates Th2 cytokines expression in asthmatic mice was measured. The levels of total NF-κB, p-NF-κB, and p-IκBα in the BALF of OVA-induced asthmatic mice were considerably higher than those in the BALF of saline-treated mice (Fig. 6). However, CTLE treatment remarkably reduced the OVA-induced NF-κB, p-NF-κB, and p-IκBα levels, although it showed higher than those in the control mice. In addition, Dex showed similar inhibitory effects on the NF-κB signaling (Fig. 6).
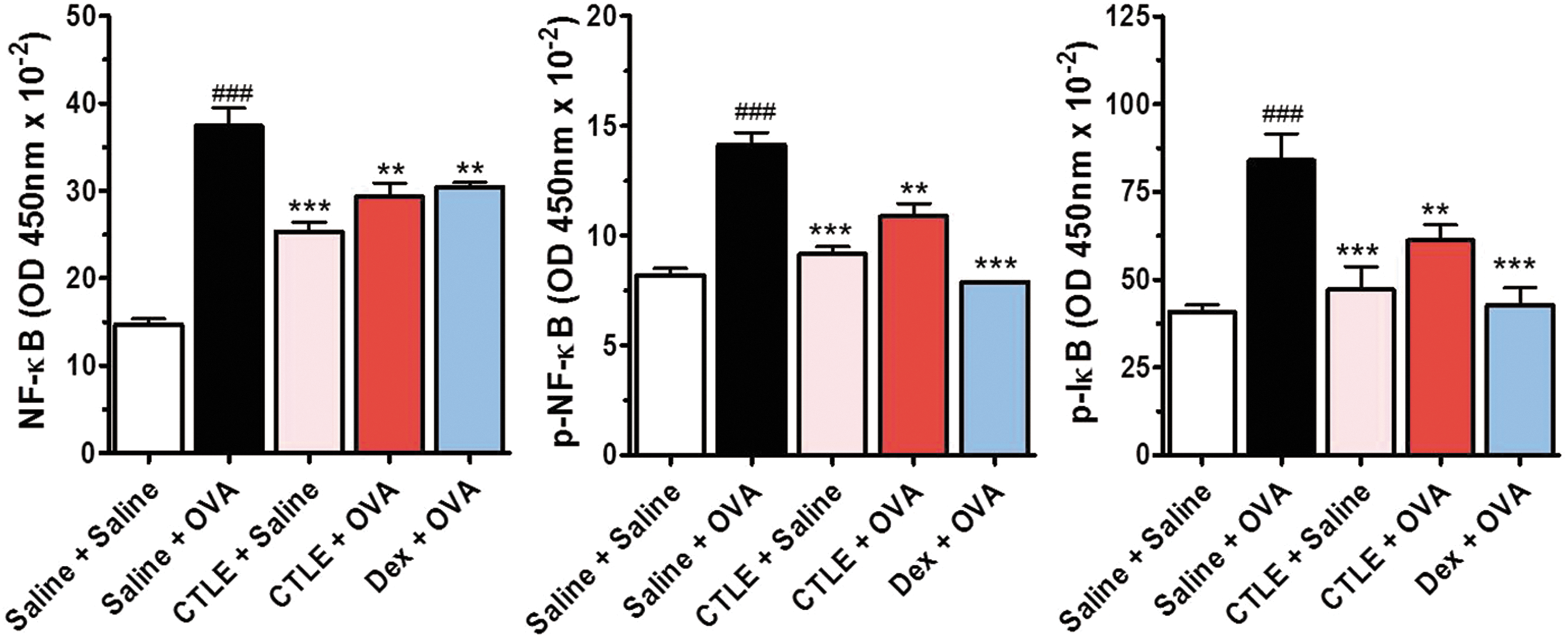
Effect of CTLE on the levels of NF-κ signaling. The level of NF-κB, p-NF-κB, and p-IκB-α in BALF was measured by ELISA. OVA increased the levels of total NF-κB, p-NF-κB, and p-IκBα in the BALF of OVA-induced asthmatic mice. However, CTLE treatment attenuated the increase of total NF-κB, p-NF-κB, and p-IκBα induced by OVA. Data were analyzed using ANOVA followed by Student's t-test. ###
P < .001, significantly different from the value of Saline group. ***P < .001, **P < .01, significantly different from the value of OVA group. Results are presented as the mean ± SD (n = 6 per group). Color images available online at
Discussion
For a long time in Korea and China, medicinal herbs have been used as ingredients of traditional medicines. Recently, because most herbs are easily usable and have no toxicity, there is growing attention on the use of herbs and their bioactive compounds. So, we analyzed the antiallergic effect of CTLE in OVA-induced animal asthma models and evaluated the therapeutic efficacy of CTLE in allergic asthma via the suppressions of Th1/Th2 imbalance and histamine release from mast cells. In this study, we revealed that CTLE effectively block compound 48/80-induced anaphylaxis via the inhibition of mast cell degranulation and histamine release. Typical features of allergic asthma induced by OVA are inflammatory cells specially eosinophils infiltrations, goblet cell hyperplasia, and collagen fiber deposit into the lung tissue. Oral administration of CTLE noticeably suppressed the influx of inflammatory cells into the submucosal regions of both perivascular and peribronchial areas, the hyperplasia of goblet cells, and the deposit of collagen fibers, and reduced the numbers of eosinophils, lymphocytes, and macrophages in BALF of asthmatic mice. Moreover, CTLE decreased the production of Th2 cytokines and NF-κB signaling activation that regulates Th2 cytokines expression and inhibited the level of OVA-specific IgE and IgG1. In addition, CTLE elevated the levels of Th1 cytokines in BALF and OVA-specific IgG2a in serum of asthmatic mice.
Allergic asthma, a class of complex and chronic diseases, are characterized by airway inflammation, elevated IgE and IgG1 levels, and airway hyperresponsiveness. 1,2,27 Also, aberrant production of the Th2 cytokines such as IL-4, IL-6, and IL-10 is the pathogenesis of asthma and leads to the infiltration of inflammatory cells into the lung. 3 –5 Of these cytokines, IL-4 plays a critical role in the promotion of an allergic inflammatory eosinophilic reaction in allergic disease. 28,29 Also, IL-5 has an important role in allergic disease by controlling the formation and proliferation of eosinophils and could enhance IL-4-induced IgE production. 28 –30 However, Th1 cytokines such as IFN-γ and IL-12 suppress Th2 cytokine productions that play critical roles in the pathogenesis of asthma. 3,5,28,29 IL-12 is well known to promote T cell differentiation toward a Th1-mediated response while suppressing the expansion and differentiation of Th2 cells. 25,28 As mentioned in many reports, imbalance of Th1/Th2 cytokines is a very important index in the evaluation of asthma. 28 –30 Our results showed that CTLE pretreatment markedly mitigates infiltrations of inflammatory cells into the submucosal regions, mucus hypersecretion, and goblet cell hyperplasia in asthmatic mice. CTLE also attenuated Th2 cytokines TNF-α, IL-4, IL-6, and IL-10 levels and increased Th1 cytokines IFN-γ and IL-12 levels. These findings suggest that CTLE could improve resolution of allergic airway inflammation via modulating Th1/Th2 cytokines.
There is a very close relationship between IgE level and Th1/Th2 cytokine production in allergic asthma. 31 Th2 cytokine IL-4 induces class switching in IgG1 and IgE, whereas IFN-γ is involved in IgG2a class switching. Consequently, the estimation of IgE production and Th1/Th2 cytokine balance is an important indicator in the assessment of asthma. Therefore, we investigated the effect of CTEL on IgE, IgG1, and IgG2a serum levels of asthmatic mice. Oral administration of CTLE noticeably alleviated the serum levels of OVA-induced total IgE, OVA-specific IgE, and OVA-specific IgG1 and increased the serum level of OVA-specific IgG2a, which were consistent with the effects of CTLE on Th1/Th2 cytokine levels. Based on these findings, CTLE may be suggested to have therapeutic efficacy for allergic asthma through the inhibition of Th2 cytokines, OVA-specific IgE and IgG1 and the promotion of Th1 cytokines and OVA-specific IgG2a.
A master transcription factor NF-κB regulates the expression of inflammatory gene products in many different cells and has a function in the progression of asthma in both patients and animal model. 7,26,32 It has been well known that NF-κB regulates Th2 cell differentiation and cytokines IL-4 and IL-5 expressions. 26 Recently, report showed that suppression of NF-κB could ameliorate OVA-induced allergic airway inflammation. 33 To reveal the underlying mechanism of CTLE in the improvement of allergic asthma, the effect of CTLE on NF-κ signaling was measured. CTLE pretreatment diminished OVA-induced NF-κB, p-NF-κB, and p-IκBα levels that induced Th2 cytokines expression. These result indicated CTLE inhibited OVA-induced asthma by downregulating NF-κ signaling pathway.
In the immune system, histamine is a critical mediator of allergic disease and asthma and the development of some allergic reactions is associated to over production of histamine. 34 Histamine also influences cytokine production via H2 receptors and through the activation of PKA. Histamine enhances the secretion of Th2 cytokines such as Il-4 and IL-5 and inhibits the production of Th1 cytokines IFN-γ and IL-12. 29,34 The elucidation of histamine's control over the cytokine network and the Th1/Th2 balance provides a basis for the potential use of antihistamines in the prevention and treatment of asthma. 29 To investigate the antihistamine effect of CTLE, the effect of CTLE on the compound 48/80-ineudced anaphylaxis and histamine releases. When mice were pretreated with CTLE for 6 days and the compound 48/80 was intraperitoneally injected to mice 1 h after the last administration of CTLE, mortality rate reduced in a dose-dependent manner. And, CTLE significantly inhibited the compound 48/80-induced serum histamine levels. As shown in these results, there was correlation between mortality rate and serum histamine levels. These results suggest that CTLE has potential protective effects via the reduction of histamine release during a systemic anaphylactic shock.
In conclusion, CTLE inhibited OVA-induced asthmatic response by reducing airway inflammation, OVA-specific IgE and IgG1 levels, and increasing OVA-specific IgG2a levels. CTLE restored Th1/Th2 balance through an increase in Th1 cytokines and decreases in Th2 cytokines and NF-κ signaling activation. Also, CTLE significantly inhibited the compound 48/80-induced anaphylaxis via histamine secretion from mast cells. The underlying mechanism of CTLE may involve to the modulation of Th1/Th2 imbalance via inhibiting the NF-κ signaling and histamine secretion. Taken together, our data suggest that CTLE could be used as a therapeutic agent for patients with Th2-mediated or histamine-mediated allergic asthma.
Footnotes
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2012R1A1A303857) and by research grants (E0121304-05) from the Korea Food Research Institute.
Author Disclosure Statement
No competing financial interests exist.