
Abstract
Metabolic diseases, such as glucose intolerance and nonalcoholic fatty-liver disease (NAFLD), are primary risk factors for life-threatening conditions such as diabetes, heart attack, stroke, and hepatic cancer. Extracts from the tropical tree Moringa oleifera show antidiabetic, antioxidant, anti-inflammatory, and anticancer effects. Fermentation can further improve the safety and nutritional value of certain foods. We investigated the efficacy of fermented M. oleifera extract (FM) against high-fat diet (HFD)-induced glucose intolerance and hepatic lipid accumulation and investigated the underlying mechanisms by analyzing expression of proteins and genes involved in glucose and lipid regulation. C57BL/6 mice were fed with normal chow diet (ND) or HFD supplemented with distilled water (DW, control), nonfermented M. oleifera extract (NFM), or FM for 10 weeks. Although body weights were similar among HFD-fed treatment groups, liver weight was decreased, and glucose tolerance test (GTT) results improved in the FM group compared with DW and NFM groups. Hepatic lipid accumulation was also lower in the FM group, and expressions of genes involved in liver lipid metabolism were upregulated. In addition, HFD-induced endoplasmic reticulum (ER) stress, oxidative stress, and lipotoxicity in quadriceps muscles were decreased by FM. Finally, proinflammatory cytokine mRNA expression was decreased by FM in the liver, epididymal adipose tissue, and quadriceps of HFD-fed mice. FMs may decrease glucose intolerance and NAFLD under HFD-induced obesity by decreasing ER stress, oxidative stress, and inflammation.
Introduction
M
The tropical tree Moringa oleifera is a well-known traditional medicine source. 3 There have been many reports on the medicinal benefits of M. oleifera, including anti-inflammatory, 4 antioxidant, 5 and anticancer effects. 6 Recent studies have also reported that M. oleifera extracts protect against diabetes in animal models. For instance, Olayaki et al. reported significant improvements in glucose tolerance and lipid metabolism in alloxan-induced diabetic rats treated with a methanolic extract of M. oleifera leaves, 7 whereas treatment of streptozotocin-induced diabetic rats with M. oleifera seed extract significantly decreased hemoglobinA1c. 8 In addition, treatment of high-fat diet (HFD)-induced obese (DIO) mice with M. oleifera leaf extract significantly improved insulin sensitivity and hepatic gluconeogenesis. 9 However, the underlying molecular mechanisms remain unknown.
Fermentation confers additional benefits to many edibles by improving safety (through preservation), palatability, and quality. 10 Indeed, many fermented foods are widely accepted for their benefits to human health. 11 Microbial food cultures such as lactic acid bacteria (LAB) are widely used for food fermentation, and LAB starter cultures adapted to plant matrix fermentation are also available. 12
In this study, we selected three strains of LAB, Lactobacillus sakei, Lactobacillus plantarum, and Lactobacillus brevis, isolated from the Korean fermented food kimchi and used them for fermentation of a methanolic M. oleifera extract. We then examined the beneficial effects of fermented M. oleifera extract (FM) against hyperglycemia and hepatic steatosis in HFD-induced obese mice compared with nonfermented M. oleifera extract (NFM) or vehicle. Our data suggest that supplementation of FM could be an effective treatment for glucose intolerance and fatty liver associated with obesity.
Materials and Methods
Animals
The animal experimental protocol was approved by the Institutional Animal Care and Use Committee on animal experiments in Handong Global University (No. Handong Animal Ethics 20151022-005). Four-week-old male C57BL/6J mice were purchased from Hyochang Bioscience (Daegu, Korea). Baseline metabolic conditions were normalized by administering a normal chow diet (ND) (D10001; Research Diets, Inc., New Brunswick, NJ, USA) for 1 week under a 12-h light/dark cycle at 22°C ± 1°C and 45% ± 10% humidity. Mice were then maintained under the same environmental conditions on ND or switched to a 60% kcal high-fat diet (D12492; Research Diets, Inc.) for 10 weeks. HFD mice were divided into three groups of 6–10 each, in which HFD was supplemented with distilled water (DW, 150 μl), FM (250 mg/kg body weight in 150 μL DW), or an equal amount of NFM by daily oral administration.
Mice were fasted for 4 h before sacrifice. Epididymal fat, liver, and quadriceps muscle tissues were rapidly excised, snap-frozen in liquid nitrogen, and stored at −75°C in a deep freezer until processed for analysis.
M. oleifera fermentation and extraction
LAB were isolated from cabbage kimchi obtained from Korean traditional markets. After incubation at 37°C for 24 h on Man, Rogosa, and Sharpe (MRS) agar (Difco Co., NJ, USA) plates, representative colonies were selected from plates yielding >106 colony-forming units. Three selected strains selected as starter cultures were identified by bioMerieux™ API™ ZYM, 50 CH Microbial Identification Strip (Fisher Scientific, PA, USA), and 16S rRNA sequencing. These strains were subcultured twice at 37°C in MRS broth before use.
For preparation of fermented extract, M. oleifera leaf powder was mixed 1:3 (w/v) in DW. Three such mixtures were inoculated with 2.5% culture media of each strain and incubated at 37°C. pH was measured using a Thermo Scientific™ Orion™ 2 Star pH Meter (Fisher Scientific). After fermentation, each sample was mixed with one volume of methanol using a Stomacher 400 blender (Seward Co., London, United Kingdom) and incubated at 37°C for 24 h. Samples were centrifuged at 4°C and 12,000 g for 15 min. The supernatants were filtered using 0.45-μM pore size Sartorius Minisart® syringe filters (Sartorius Corporate Administration GmbH, Gottingen, Germany), followed by evaporation and lyophilization.
Glucose tolerance test
After 8 weeks of M. oleifera extract or control treatment, mice were fasted for 16 h and injected intraperitoneally with glucose (2 g/kg). Blood glucose levels were measured by Accu-Check Go (Roche Diagnostics GmbH, Basel, Switzerland) from tail blood samples at 0, 15, 30, 60, 90, and 120 min after injection.
Liver histology
Liver tissues were isolated immediately after sacrifice. For hematoxylin and eosin (H&E) staining, tissues were fixed in 10% formalin and embedded in paraffin. Images were obtained under a microscope at × 200 magnification.
Western blotting
Tissue proteins were extracted using the PRO-PREP protein extraction solution (iNtRON Biotechnology, Seongnam, Korea), centrifuged, boiled, separated by 10% SDS-PAGE, and transferred to PVDF membranes. Membranes were blocked with 5% skim milk, washed in TBS-T, and incubated with primary antibodies against 5′-AMP-activated protein kinase (AMPK), phosphorylated AMPK (pAMPK), binding immunoglobulin protein (BiP), protein disulfide isomerase (PDI) (Cell Signaling Technology, Beverly, MA, USA), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Bioss Antibodies, Woburn, MA, USA). Membranes were then washed and incubated with anti-rabbit IgG conjugated to horseradish peroxidase (Cell Signaling Technology). Signals were detected by enhanced chemiluminescence and analyzed by an AlphaImager 2200 (Protein Simple, Santa Clara, CA, USA).
Quantitative real-time polymerase chain reaction
Total RNA was isolated with TRIzol® reagent (Thermo Fisher Scientific, Waltham, MA) and reverse-transcribed with the ImProm-II™ reverse transcription system (Promega, Madison, WI). Quantitative polymerase chain reaction (PCR) was performed with VeriQuest™ SYBR® Green qPCR Master Mix (Affymetrix, Inc., Santa Clara, CA) on an ABI 7500 Real-Time PCR system (Applied Biosystems, Foster City, CA). The following mRNA expression levels were detected using gene-specific primers (sequences available on request): acetyl-CoA carboxylase (ACC), adipose triglyceride lipase (ATGL), BiP, carnitine palmitoyltransferase 1 (CPT1), CCAAT/enhancer-binding protein α (C/EBPα), CCAAT-enhancer-binding protein homologous protein (CHOP), cluster of differentiation molecule 36 (CD36), fatty-acid synthase (FAS), hormone-sensitive lipase (HSL), interleukin-1β (IL-1β), IL-6, IL-12, lipoprotein lipase (LPL), monocyte chemoattractant protein 1 (MCP1), monoglyceride lipase, peroxisomal acyl-CoA oxidase 1 (ACOX1), peroxisome proliferator-activated receptor α (PPARα), PPARγ, PDI, sterol regulatory element-binding protein 1c (SREBP1c), tribbles homolog 3 (TRB3), tumor necrosis factor α (TNFα), uncoupling protein 2 (UCP2), and UCP3. Results are presented as mean ± standard deviation (S.D.) normalized to GAPDH expression using the ΔΔCt method.
Enzyme-linked immunosorbent assay
The mouse TNFα and IL-1β enzyme-linked immunosorbent assay set were purchased from BioLegend® (San Diego, CA). All experiments were performed according to the manufacturer's instructions.
Statistical analysis
All data are expressed as mean ± S.D. Comparisons among different groups were performed by analysis of variance. Significant difference between controls and experimental groups were assessed by Tukey's multiple comparison test using the software GraphPad Prism version 7.00 for Windows. P values <.05 were considered to be significant.
Results
Improved glucose tolerance in DIO mice by FM
We examined the impact of M. oleifera extract supplementation on weight gain and glucose tolerance in HFD-induced obese mice. After 10 weeks of HFD feeding, body weight was significantly higher in HFD-fed control (HFD+DW) mice relative to ND+DW mice (week 1–9; P < .001, week 2; P < .01), and neither FM nor NFM altered HFD-induced adiposity (Fig. 1A). Although there were no significant differences in epididymal adipose tissue (EAT) and quadriceps muscle weights among HFD-fed mouse groups (DW, NFM, and FM), liver weight was markedly lower in FM-treated HFD mice (HFD+FM) than in HFD+NFM and HFD+DW mice (Fig. 1B).

Effect of FM treatment on weight gain, tissue weight, and glucose intolerance.
To examine the effect of M. oleifera extract on glucose tolerance, glucose tolerance test (GTT) results were compared between ND-fed and HFD-fed mouse groups. After 8 weeks of treatment, HFD+FM mice exhibited significantly better glucose tolerance than HFD+DW (60 min; P < .05, 90 min; P < .001, 120 min; P < .01) and HFD+NFM mice (60 min; P < .05, 90 min; P < .01, 120 min; P < .05, Fig. 1C).
Suppression of hepatic adiposity and lipid production by FM
To assess the effect of FM on hepatic lipid accumulation, liver sections were visualized by H&E staining. The HFD-fed control (HFD+DW) mice showed a remarkable increase in liver fat content compared with ND+DW mice, whereas HFD-induced lipid accumulation was almost completely reversed by FM but only partially by NFM (Fig. 2A). These staining results were verified by measuring hepatic fat content, which revealed strongly decreased hepatic adiposity by FM compared with control (P < .001) and NFM (P < .001) and partial reduction by NFM (P < .001, Fig. 2B). To elucidate the molecular mechanism underlying the suppression of liver fat accumulation by extract treatment, expressions of hepatic genes related to lipid metabolism were analyzed by quantitative real-time polymerase chain reaction (qRT-PCR). The expressions of the lipogenic genes ACC (P < .01), FAS (P < .001), and SREBP1c (P < .01) were significantly downregulated in FM-treated and LPL (P < .01) was markedly decreased only in NFM-treated HFD mice compared with HFD+DW mice (Fig. 2C). There were no significant gene expression changes for C/EBPα and PPARγ among HFD groups. In contrast, expressions of genes related to lipid uptake, oxidation, and lipolytic activity, including CD36 (P < .001), ACOX1 (P = .07), ATGL (P < .05), and HSL (P = .07), were markedly increased only in HFD+FM mice relative to HFD+DW control (Fig. 2D). There were no differences in CPT1 and PPARα expression among HFD groups. Hepatic protein expression levels of AMPK and pAMPK were significantly increased in both NFM- and FM-treated HFD mice, with higher pAMPK expression in FM-treated mice (P < .01, Fig. 2E).
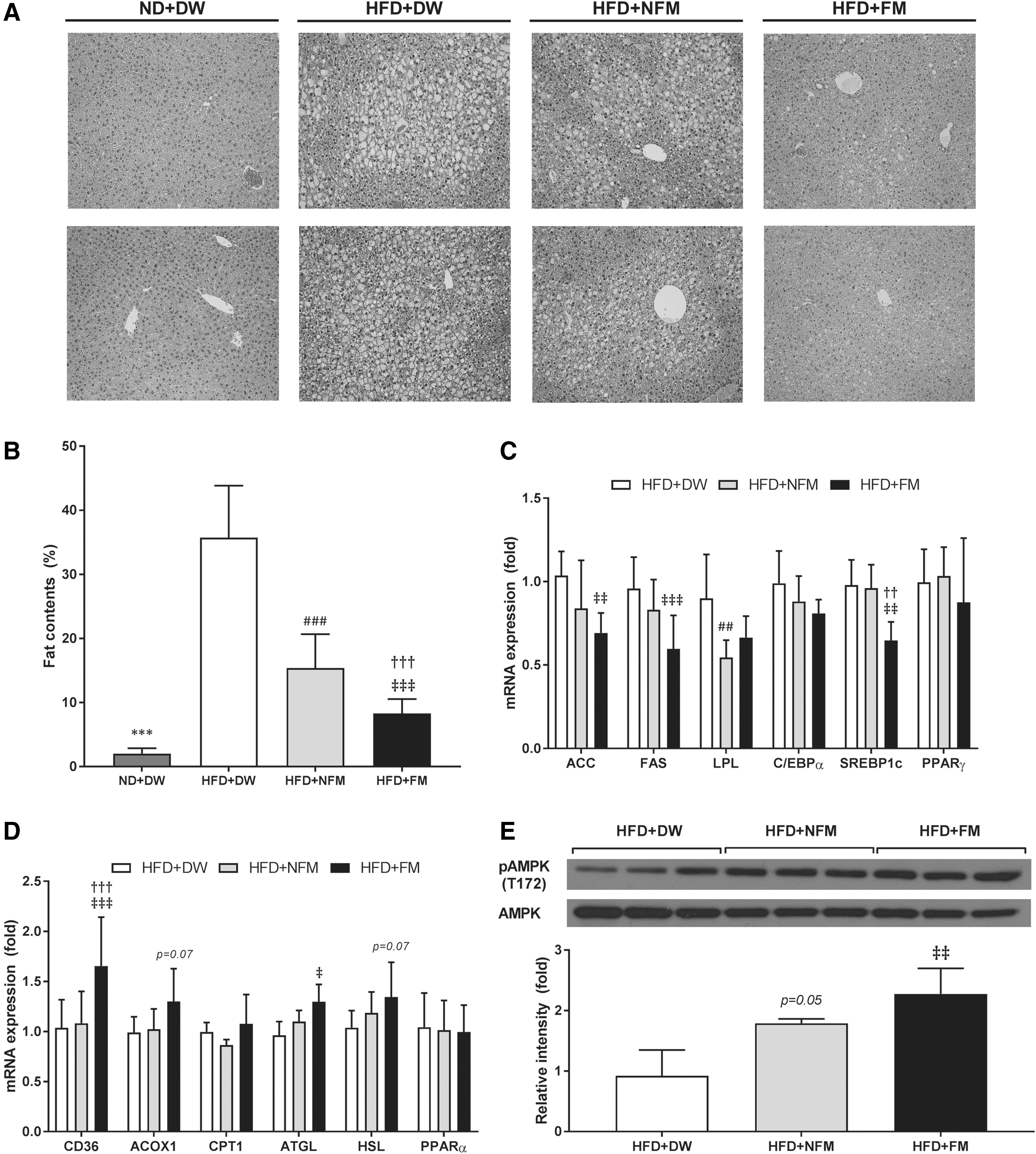
Effect of FM treatment on hepatic lipid accumulation and expression of genes involved in hepatic lipid metabolism.
Reduction of endoplasmic reticulum stress in skeletal muscle through modulation of lipid metabolism by FM
Endoplasmic reticulum (ER) stress has emerged as a key factor in the onset of IR. 13 To examine whether the enhancement of glucose tolerance by FM is attributable to decreased ER stress in skeletal muscle, expression of genes related to ER stress was measured, including BiP, PDI, CHOP, and TRB3. BiP and CHOP gene expressions were decreased in FM-treated mice (P < .01), and PDI mRNA expression was substantially decreased by both NFM (P < .05) and FM treatment of HFD mice (Fig. 3A). TRB3 level was not changed by either extract. BiP and PDI protein levels were also significantly decreased by NFM and FM compared with HFD+DW (P < .01, Fig. 3B).
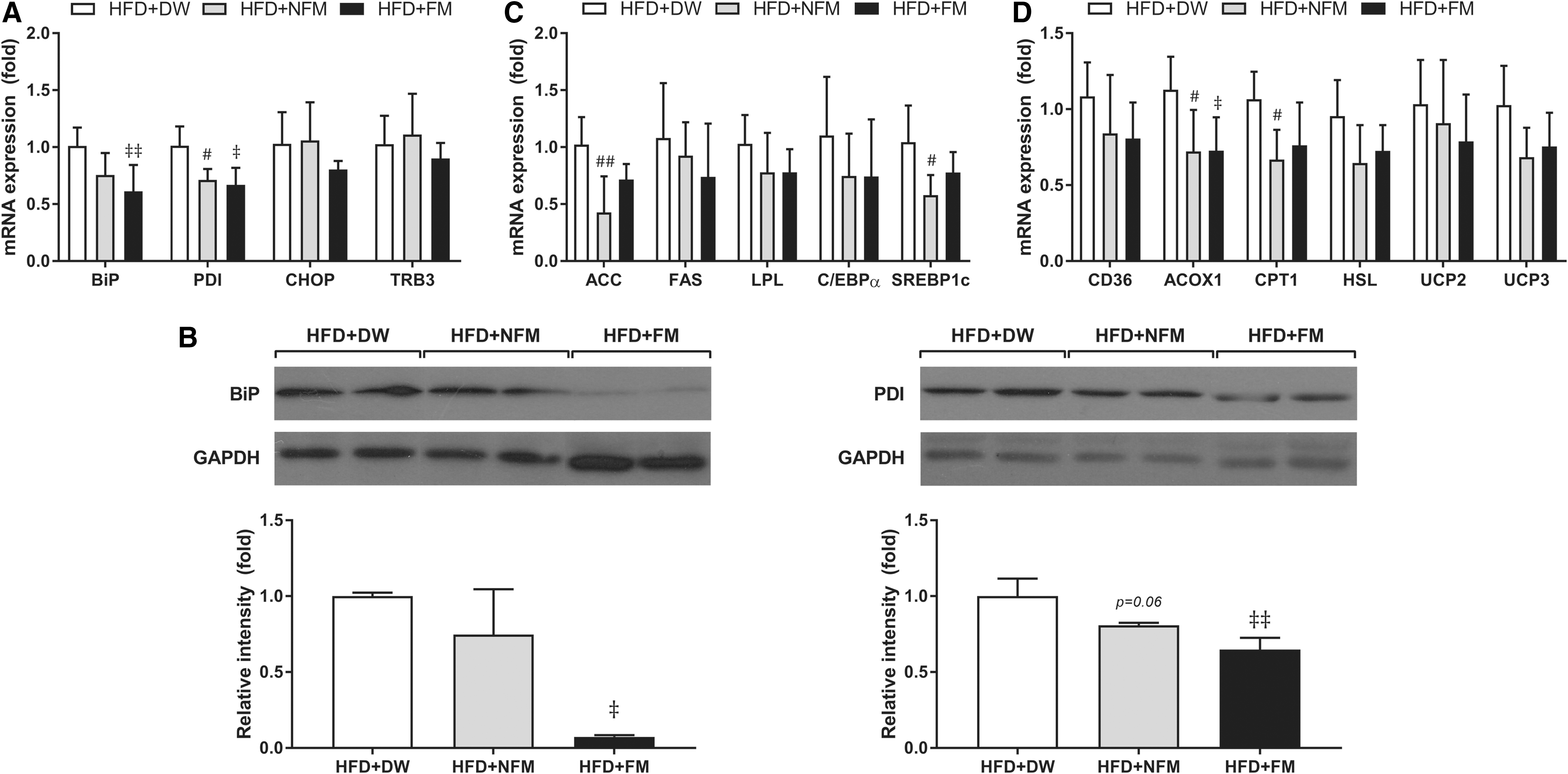
Effect of FM treatment on genes related to ER stress and lipid metabolism in skeletal muscle.
Several studies have reported that ER stress is involved in the association between saturated nonesterified fatty-acid-induced inflammation and IR. 14 To investigate a possible mechanism underlying ER-stress reduction in HFD+FM mice, expression of genes involved in lipid synthesis and oxidation was analyzed in skeletal muscle. The expression of the lipogenic genes ACC (NFM; P < .01), FAS, LPL, C/EBPα, and SREBP1c (NFM; P < .05) was decreased in both NFM-fed and FM-treated HFD mice compared with HFD+DW controls (Fig. 3C). Similarly, expressions of the lipid oxidative genes CD36, ACOX1, CPT1 (NFM), and HSL and of the oxidative stress genes UCP2 and UCP3 were decreased by NFM and FM (Fig. 3D).
Inhibition of chronic low-grade inflammation by FM
Chronic low-grade inflammation is another major contributor to obesity-induced glucose intolerance. 15 We measured the expression of proinflammatory cytokines in the liver, EAT, and quadriceps muscle. mRNA expression of the proinflammatory cytokines TNFα, IL-6, and IL-12 in the liver was markedly decreased by FM (Fig. 4A) and TNFα (P < .01), IL-1β and IL-6 (NFM and FM) in the quadriceps was also decreased by FM (Fig. 4C). In the adipose tissue, expressions of all these proinflammatory genes plus MCP1 (NFM; P < .05, FM; P < .01) were downregulated by FM (Fig. 4B). In addition, the protein expression level of TNFα (NFM; P < .01 and FM; P < .05) in EAT and quadriceps was decreased by FM and NFM, but the expression level of IL-1β was decreased only in EAT (Fig. 4D).
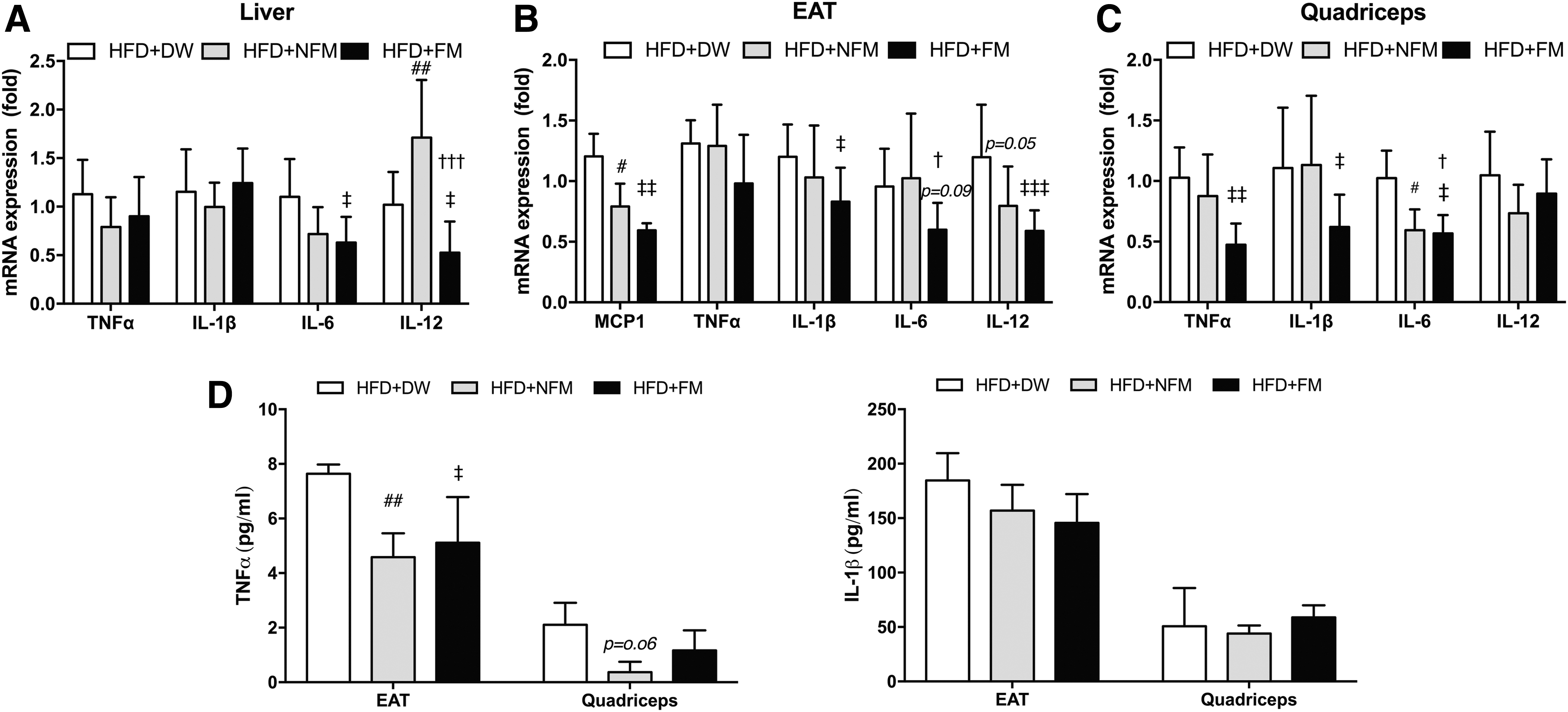
Effect of FM treatment on proinflammatory cytokines.
Discussion
Obesity, glucose intolerance, and nonalcoholic fatty-liver disease (NAFLD) are increasing in frequency, and they collectively constitute a major healthcare burden. Although many drugs for metabolic syndromes have already been developed, side effects such as weight increase, liver toxicity, and cardiac insufficiency limit their applicability and efficacy. 2 M. oleifera is one of the most widely grown trees in tropical and subtropical regions, and many parts of M. oleifera are edible. Especially, leaves, rich in vitamins, minerals, and protein, are the most nutritious part; therefore, M. olefera is used as a conventional nutritional supplement and also medicine. 16 Extracts of M. oleifera have demonstrated antidiabetic, anti-inflammatory, antioxidant, and even antitumor effects and may ameliorate the metabolic sequela of obesity. 4 –9 We prepared a methanolic extract from M. oleifera leaves and fermented it with three strains of Lactobacillus. Although supplementation of a HFD with FM did not decrease body weight gain, it did reverse the increase in liver mass and fat content, and partially reversed glucose intolerance in these DIO mice. The decrease in HFD-induced liver fat accumulation was greater in FM-treated mice than NFM-treated mice, suggesting additional benefits of fermentation. Expression of multiple lipid synthetic genes was decreased, whereas expression of genes related to lipid oxidation was upregulated by FM. pAMPK expression was increased by FM compared with control HFD mice. In addition, expression of proteins and genes involved in ER and oxidative stresses was decreased in the quadriceps of FM-treated mice, and expression of proinflammatory genes was decreased in the liver, EAT, and quadriceps by FM.
NAFLD is initiated and exacerbated by hepatic fat accumulation. 17 It is often associated with IR and other metabolic disorders. 18 In the present study, HFD-fed mice exhibited significantly increased body weight compared with ND-fed mice, and neither NFM nor FM had any effect on weight gain (Fig. 1A), likely because neither supplement had any effect on food intake (data not shown). However, liver weight (but not EAT or quadriceps weight) was decreased by FM (Fig. 1B). We then assessed whether decreased liver weight was because of changes in lipid synthesis and/or lipid oxidation. Hepatic lipid accumulation associated with HFD was markedly decreased by FM (Fig. 2A). Moreover, expression of the lipogenic genes ACC and FAS, as well as their transcriptional factors SREBP1c 19 and C/EBPα, 20 was decreased, as was LPL expression, the major source of nonesterified fatty acids. 21 Conversely, expression of genes related to fatty-acid uptake (CD36), 22 fatty-acid β-oxidation (ACOX1), 23 and lipolysis (ATGL and HSL) 24 was upregulated in the liver by FM (Fig. 2C, D). AMPK 25,26 acts as an energy sensor and has been suggested as a therapeutic target in NAFLD and related metabolic diseases. 27 AMPK phosphorylation was elevated in FM-treated mice (Fig. 2E). Collectively, decreased lipogenesis, increased lipolysis, and greater AMPK activation by FM may contribute to the decreased hepatic lipid accumulation in FM-treated HFD-fed mice. Thus, FM supplementation may slow the progression of NAFLD.
Glucose tolerance is another major characteristic of metabolic syndrome. 28 Treatment with FM improved glucose regulation in the HFD-fed mice after 8 weeks (Fig. 1C). In contrast to a previous study assessing the antidiabetic effects of M. oleifera, 9 we found no effects of FM and NFM on body weight. Several drugs for treatment of type 2 diabetes are associated with weight gain, 2 whereas supplementation with FM had no such effects in mice. Several studies have reported that the improvement of glucose intolerance is coincident with enhanced activation of insulin signaling pathways. 29 –31 To monitor the activation of insulin signaling by FM treatment, we measured the Akt phosphorylation level in the liver, EAT, and quadriceps muscle. Unexpectedly, Akt phosphorylation was unchanged by FM and also by CM in HFD-fed mice (data not shown).
In addition, recent studies have implicated ER stress in the development of glucose intolerance. Fu demonstrated that ER stress acts as a negative modulator of insulin sensitivity 32 ; thus, decreased ER stress may enhance glucose tolerance. 33,34 We speculated that decreased ER stress contributes to the improved GTT results observed in FM-treated HFD-fed mice. Indeed, expression of the ER protein chaperon BiP and the disulfide bond regulator PDI was decreased in NFM-fed and FM-treated mice, whereas the transcriptional factor CHOP and apoptosis-associated TRB3 were decreased only in FM-treated mice (Fig. 3A). 13,32,35 Decreased BiP and PDI expression in NFM-fed and FM-treated mice (Fig. 3B) was consistent with decreased ER stress. Several studies have demonstrated that palmitate-induced IR is mediated by ER stress in skeletal muscle. 33 In addition, Akt is inactivated by direct exposure to palmitate in myocytes. 36 We observed decreased expression of the lipogenic enzymes ACC and LPL (Fig. 3C), as well as lipid oxidation mediators CD36, ACOX1, CPT1, and HSL (Fig. 3D). Lipid intermediates, such as diacylglycerol, ceramides, and fatty acyl-CoAs, converted from fatty acids can impair the function of cells, which is known as lipotoxicity. In skeletal muscle, there is a negative correlation between lipid levels and insulin sensitivity, and increased lipid levels are regarded as indicator of IR and also as a causative factor in obesity-associated IR. 37 A few studies revealed that lipotoxicity-induced oxidative stress in skeletal muscle under diabetic condition is triggered by ER stress, 38 as evidenced by increased expression of UCP2 and UCP3, which attenuate reactive oxygen species production by mitocondira. 39,40 In addition, increased oxidative stress activates the expression of proinflammatory genes by activating transcriptional factors. 40 Consistent with this notion, we found decreased UCP2 and UCP3 mRNA expression levels in skeletal muscle in FM-treated mice. Therefore, treatment with NFM and FM could improve glucose tolerance by alleviating ER stress, lipotoxicity, and oxidative stress.
It has been reported that chronic low-grade inflammation in peripheral tissues leads to metabolic impairment and aggravates metabolic disease. 15,41 We found that multiple proinflammatory cytokines were elevated in the liver, EAT, and quadriceps of HFD-fed obese mice. In all these tissues, however, elevations in TNFα and the proinflammatory cytokines IL-1β, IL-6, and IL-12 were significantly reversed by FM (Fig. 4A–C). Obese patients exhibit increased infiltration of macrophages in adipose tissue compared with lean subjects. Macrophage attractant MCP-1 42 expression was markedly decreased in the adipose tissue of FM-treated mice. It thus appears that FM decreased chronic low-grade inflammation throughout the body.
In conclusion, we demonstrated that FM treatment of DIO mice decreases hepatic lipid accumulation and improves glucose tolerance by enhancing lipid metabolism, decreasing ER and oxidative stress, and suppressing inflammation. These findings support previous studies showing antidiabetic effects of M. oleifera extracts, and also demonstrate the added benefits of M. oleifera fermentation by three strains of Lactobacillus. FM may be an effective dietary supplement for the treatment of hyperglycemia and NAFLD.
Footnotes
Acknowledgment
This research was supported by No. 20140168 of Handong Global University Research Grants.
Author Disclosure Statement
No competing financial interests exist.