
Abstract
The methanol extract of Mammea longifolia Planch. and Triana (M. longifolia) fruit was studied for anticancer and apoptotic effects in the SW480 colon cancer cell line. The apoptotic and necrotic effects of M. longifolia were detected by 3-(4,5-dimethyl-2-thiazolyl)-2, 5-diphenyl-2H tetrazolium bromide (MTT) and lactate dehydrogenase assays, respectively. One hundred μg/mL of the extract killed ∼82.4% of the cells; however, 2% of the death was related to necrosis. The morphological changes in M. longifolia-stimulated SW480 cells were observed directly by light microscopy. DNA fragmentation assay was employed to analyze the apoptosis induction. M. longifolia-treated SW480 cells promoted the expression of Bax, Bad, cleaved-poly-ADP-ribose polymerase (PARP), and p53 proteins and decreased the protein expression of pro-caspases Bcl-2 and Bcl-XL. The ratios of Bax/Bcl-2 and cleaved-PARP/PARP, predictive markers of apoptotic stimuli in cancer, increased and may play an important role in regulating the progression of apoptosis. The results suggested that M. longifolia induces cell death via mitochondrial-related apoptosis in SW480 cells.
Introduction
T
For treating early detected colon cancer, doctors usually use surgery, chemotherapy, and radiation therapy. 6 Targeted drugs, such as bevacizumab, cetuximab, panitumumab, and regorafenib, have also been used for treating colon cancer. 7 Targeted therapies can deliver a high concentration of medication directly to the lesions. In cancer cells, the drugs block essential biochemical pathways or target mutant proteins and inhibit cancer cell growth and survival. 8 However, due to the high cost, these treatments are not available for all cancer patients. Thus, natural anticancer drugs, which are less expensive, are the subject of great interest.
Mammea longifolia is a common tree in India. Its flower buds are extensively used in cooking, especially in spice blends. Due to its stimulating, carminative, and astringent effects, the buds are used in treating dyspepsia and hemorrhoids. 9 Physical and chemical studies report that 13 bioactive compounds can be isolated from Mammeas flowers and these have benefits such as radical scavenging, antitumor, anti-HIV, antifungal, and antibacterial activities. 10,11 However, there is relatively little research on the biological activities of M. longifolia fruit.
In our previous research, the methanol extract from M. longifolia fruit exhibited considerable anticancer effects on human colon cancer cells. 12 Thus, we investigated the potential apoptotic effects and the related molecular mechanisms of M. longifolia on the human colon cancer cell line, SW480.
Materials and Methods
Samples and chemicals
The methanol extract from the fruit of M. longifolia Planch. and Triana (M. longifolia) was purchased from the Plant Extract Bank (Seoul, Korea). The dried extract was dissolved in dimethyl sulfoxide (DMSO) to a final concentration of 0.1% (v/v). Control groups consisted of medium containing 0.1% DMSO without M. longifolia. DMSO, Triton X-100, Pierce bicinchoninic acid (BCA), tris-base, NaCl, ammonium persulfate, and Tween 20 were purchased from Sigma Co. (St. Louis, MO). Roswell Park Memorial Institute medium 1640 (RPMI 1640), fetal bovine serum (FBS), penicillin-streptomycin, trypsin-EDTA, RNase, and proteinase K were purchased from HyClone Laboratories, Inc. (Logan, UT); 3-(4,5-dimethyl-2-thiazolyl)-2, 5-diphenyl-2H tetrazolium bromide (MTT), 20 × phosphate buffered saline (PBS), 50 × Tris-acetate-EDTA (TAE), DNA lysis buffer, NP-40 lysis buffer, 1.5 M Tri-HCl (pH 8.8), and 1 M Tri-HCl (pH 6.8) were purchased from Biosesang, Inc. (Gyonggi, Korea). Lactate dehydrogenase (LDH) kit was purchased from Takara Bio Co. (Otsu, Japan). One hundred base pairs DNA Ladder was purchased from Bioneer, Inc. (Daejeon, Korea). N, N, N′, N′-tetramethylethylenediamine was purchased from Bio-Rad Laboratories, Inc. (Hercules, CA). Antibodies to β-actin, Caspase-3, −8, and −9, poly-ADP-ribose polymerase (PARP), p53, Bax, Bad, Bcl-2, and Bcl-XL were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA).
Cell line and cell culture
Embryonic kidney cell (HEK293) line was purchased from the Korean Cell Line Bank (Seoul, Korea) and grown in Dulbecco's modified Eagle's medium; and the human colon cancer SW480 cell line (ATCC CCL-228) was purchased from the American Type Culture Collection and maintained in RPMI 1640 medium. Both cell lines were supplemented with 5% (v/v) FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C in 95% humidified air containing 5% carbon dioxide (CO2).
MTT assay
The SW480 and HEK293 cells were plated at a density of 2 × 104 cells/well in a 96-well plate and incubated at 37°C in a humidified atmosphere with 5% CO2 for 24 h. Then, the cells were treated with 25, 50, 100, and 200 μg/mL of M. longifolia for another 24 h. After incubation, the supernatant of each well was discarded. A 200 μL aliquot of 1X MTT solution (0.5 mg/mL) was added to each well and incubated for 4 h. In addition, the supernatant was removed, and 200 μL/well DMSO was added to dissolve the formazan crystals. The absorbance was measured at 550 nm by using an enzyme-linked immunosorbent assay (ELISA) plate reader (Bio-Tek, Winooski, VT). The percent of cell viability was determined with respect to the control group.
LDH assay
The SW480 cells were plated in a 96-well plate (2 × 104 cells/well) for 24 h. After treatment with/without different concentrations of M. longifolia (25, 50, 100, and 200 μg/mL) for 24 h, 4 μL of triton X-100 (2%, v/v) was added to the negative control group 1 h before the measurement. The supernatants (100 μL) were mixed with a 100 μL prepared detection kit reagent and incubated for 30 min. Next, the absorbance was measured at 490 nm by an ELISA plate reader. The percent of LDH was determined with respect to the triton X-100 group.
Light microscopic examination
Ten thousand SW480 cells/well were seeded in six-well plates and incubated for 24 h. After treatment with/without 100 μg/mL of M. longifolia for 24 h, the cells were observed under a light microscope (Olympus, Tokyo, Japan).
DNA fragmentation assay
SW480 cells were seeded in a 35-mm cell culture dish (1 × 106 cells/well) for 24 h. Then, the cells were treated with 25, 50, and 100 μg/mL of M. longifolia for 24 h. The harvested cells were washed with PBS and lysed in a 100 μL DNA lysis buffer. After centrifugation, the supernatant was incubated with RNase (10 mg/mL) and proteinase K (2 mg/mL) at 37°C for 1 h. The DNA samples were analyzed in 1% agarose gels in TAE buffer, and DNA bands were visualized by ethidium bromide staining and a Mini BIS image analysis system (DNR Bio-Imaging Systems Ltd., Jerusalem, Israel).
Western blot analysis
SW480 cells were plated in six-well plates at ∼1 × 106 cells/well, and media were replaced with 5% FBS/RPMI for 24 h. After the cells were treated with 25, 50, and 100 μg of M. longifolia for 24 h, the cells were harvested and washed twice with PBS. The prepared cells were lysed with cell lysis buffer (150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate [SDS], 50 mM Tris-HCl, and 2 mM EDTA) for 30 min at 4°C. Homogenates were centrifuged for 10 min at 13,000 g. The protein concentration in the supernatant was determined by using the BCA protein assay.
Thirty μg of the isolated protein was separated on 10% SDS-polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride (Bio-Rad Laboratories, Inc.) membrane. The membrane was blocked with 5% BSA in Tris-buffered saline-Tween (TBST; 20 mM Tris, 500 mM NaCl, pH 7.5, and 0.1% Tween 20) at room temperature for 1 h. Subsequently, the membrane was incubated with a primary antibody against each target protein in 5% BSA for 1 h. Then, the membrane was washed three times with TBST for 5 min. The membrane was then incubated with horseradish peroxidase-conjugated secondary antibody at room temperature for another 1 h. The protein bands were visualized by using an enhanced chemiluminescence detection system. Blots were developed with a Mini BIS image analysis system.
Statistical analysis
All tests were carried out independently in triplicate. Data are expressed as mean ± standard derivation. One-way analysis of variance followed by Tukey's post hoc test using SPSS 19.0 software (IBM, Armonk, NY) was performed to compare the difference between the means. Values of P < .05 were considered statistically significant. Means with the same letter in each figure are not significantly different.
Results
M. longifolia reduces the cell viability in SW480 cells
In this study, the MTT assay was performed to measure the cell viabilities of M. longifolia-treated SW480 cells (Fig. 1A) and HEK293 cells (Supplementary Fig. S1; Supplementary Data are available online at

The apoptotic and necrotic effects of M. longifolia on human colon cancer cell line, SW480.
M. longifolia-induced necrotic cell death was tested by an LDH assay. As shown in Figure 1B, the LDH content in the prepared medium was low when the dosage of the M. longifolia was lower than 200 μg/mL.
The morphological changes in M. longifolia-stimulated SW480 cells were observed directly by light microscopy (Fig. 1C). The untreated cells were spindle shaped and formed cohesive groups. In contrast, cells treated with 100 μg/mL of M. longifolia were round shaped and observed in wider intercellular spaces.
M. longifolia induced DNA fragmentation
A DNA fragmentation assay was employed to analyze the apoptotic induction activity of M. longifolia on SW480 cells. A laddering pattern post-agarose gel electrophoresis (Fig. 2) indicated that the DNA of the M. longifolia-stimulated cells was degraded into giant DNA fragments. In addition, increased DNA laddering was observed with higher doses of M. longifolia. In contrast, the chromosomal DNA of the untreated cells remained intact.
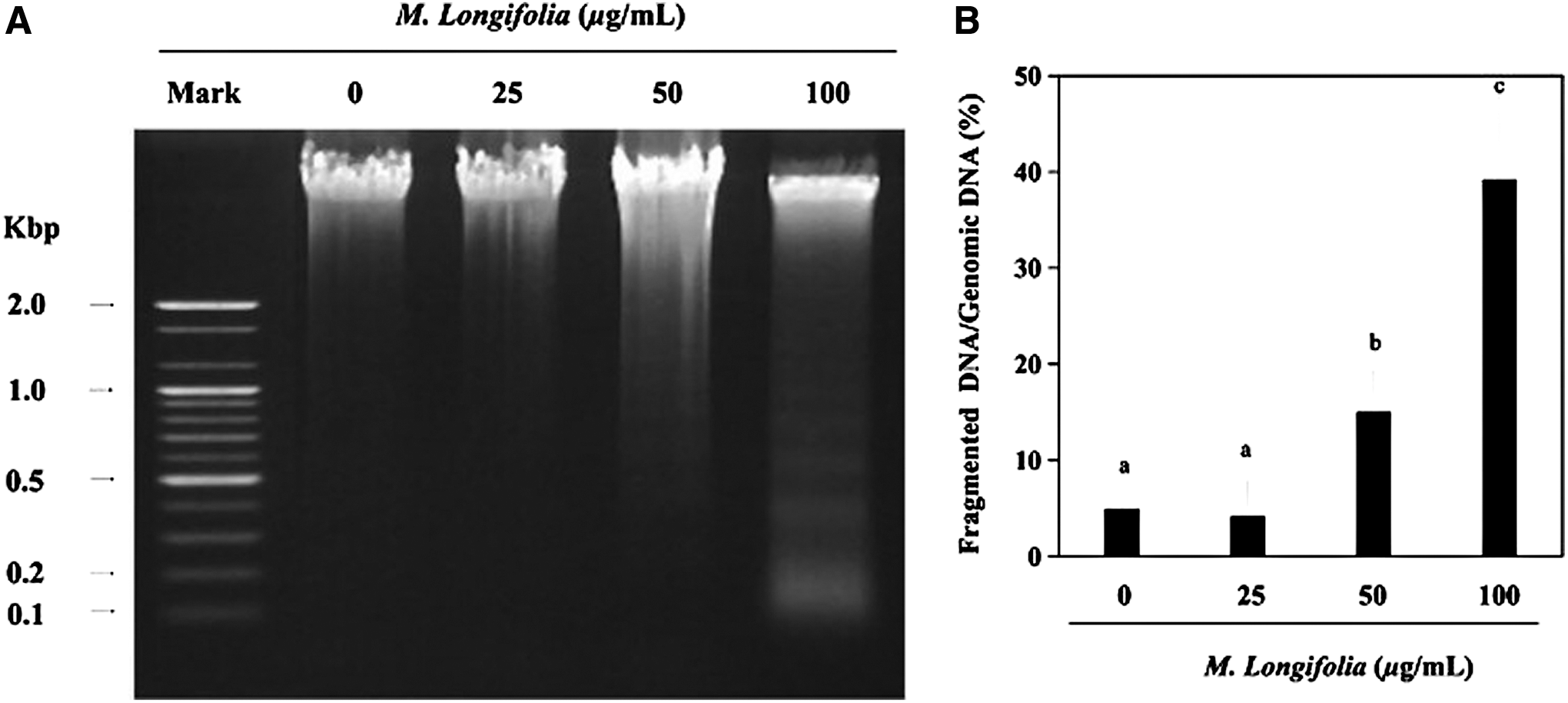
Effect of M. longifolia on chromosomal DNA fragmentation. Samples were prepared from SW480 cells that were incubated with 25, 50, and 100 μg/mL of M. longifolia extract for 24 h.
M. longifolia weakened pro-caspase expression
After stimulation with M. longifolia, proteins of pro-caspase 3, pro-caspase 8, and pro-caspase 9 were gradually decreased in SW480 cells (Fig. 3A). Moreover, the indistinct cleaved-pro-caspase 3 (20 kD) appeared. The main substrate, PARP, was cleaved into two fragments when apoptosis started. The cleaved PARP/PARP ratio increased in a dose-dependent manner after cells were incubated with M. longifolia for 24 h (Fig. 3B).
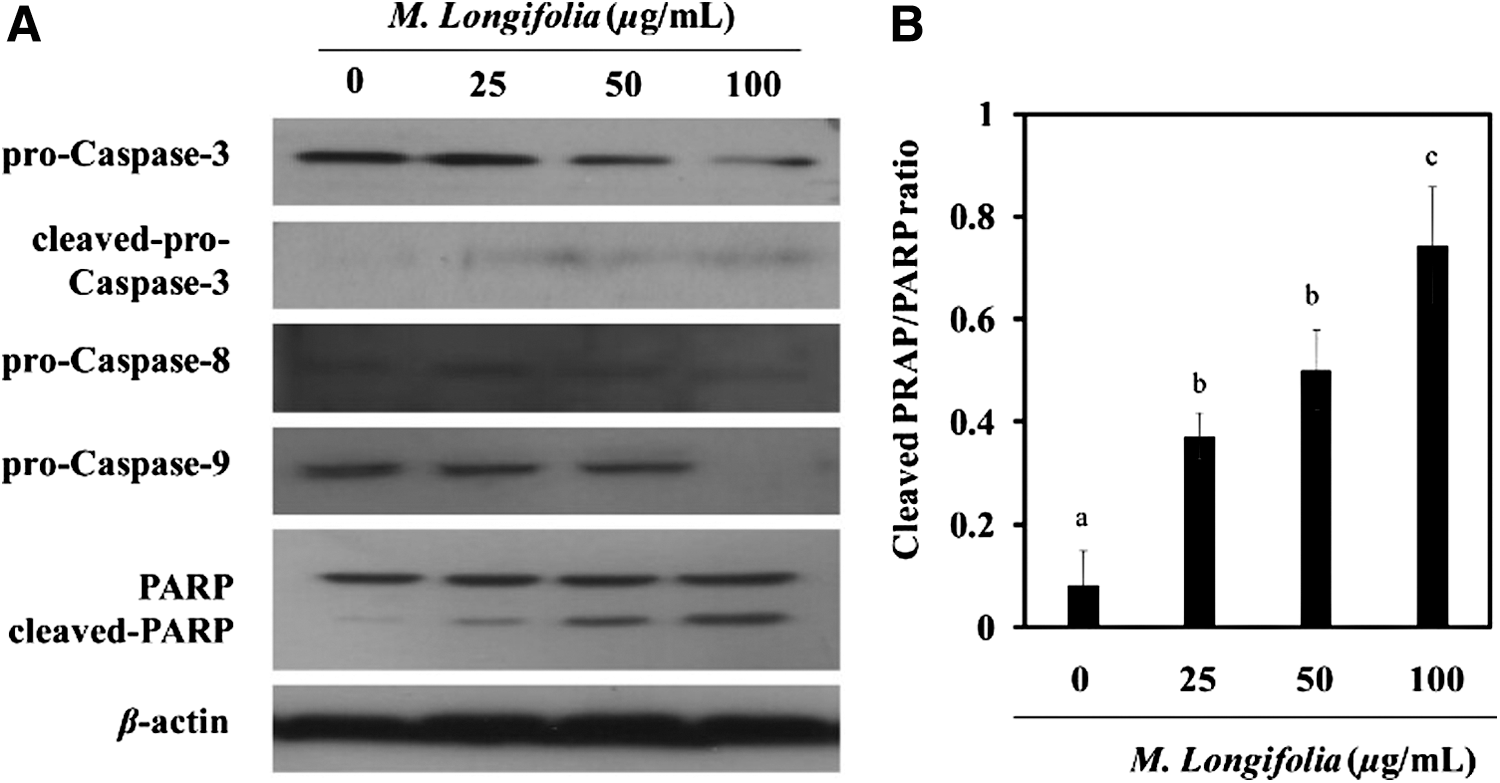
Effects of M. longifolia on pro-caspase expression.
M. longifolia enhanced the p53 expression
Western blot analysis was used to screen for inducible p53 protein expression. Analysis of protein expression in SW480 cells shows the minimum levels of p53. M. longifolia stimulation led to a remarkable increase of p53 levels in SW480 cells (Fig. 4). The treatment increased the relative intensity of the band from 0.27 to 0.85.
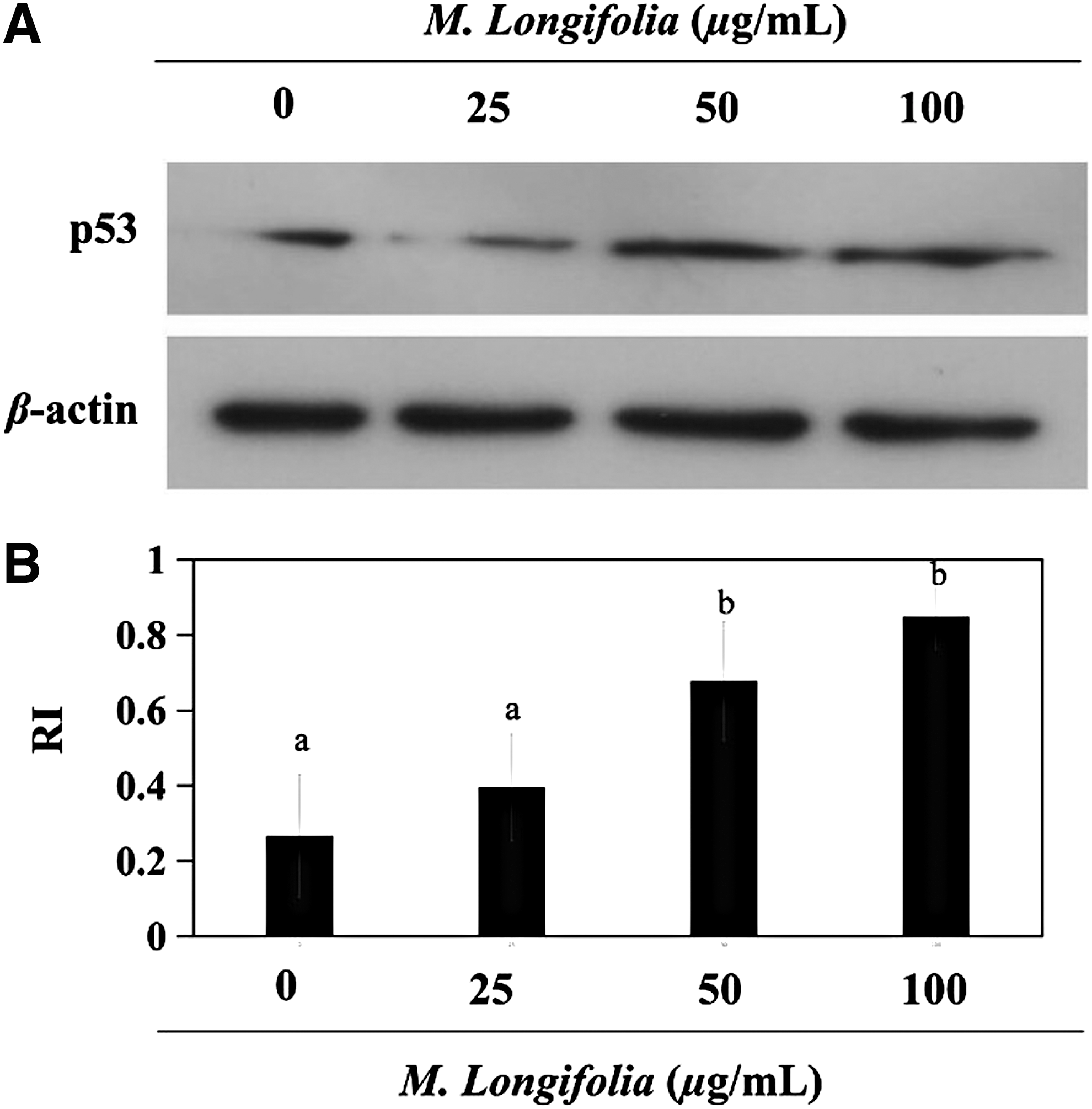
Effects of M. longifolia on the expression p53.
M. longifolia affected the Bcl-2 family
To investigate the effects of M. longifolia on the Bcl-2 family, which belongs to the mitochondrial pathway, proteins of Bax, Bad, Bcl-2, and Bcl-XL expression were measured by the Western blot assay. M. longifolia stimulation upregulated Bax and Bad expression; however, it downregulated Bcl-2 expression (Fig. 5A). The Bax/Bcl-2 ratio increased from 0.59 to 3.79 (Fig. 5B). These results revealed clear Bax overexpression, and an increase in the Bax/Bcl-2 ratio accelerated apoptosis that was induced by the M. longifolia in the SW480 cell line.

Effects of M. longifolia on the expression of mitochondria-related proteins.
Discussion
Cell death typically occurs by one of the following two distinct processes: apoptosis or necrosis. Apoptosis is an active and programmed process of cell death. 13 It is proposed to be related to cardiomyocyte loss during myocardial hypoxia, ischemia, infarction, mechanical stress, aortic constriction, and heart failure. It has also been described as nontraumatic cell death. 14,15 Necrosis is death that is due to unexpected or accidental cell damage, resulting from toxic chemical or physical perturbations such as toxins, radiation, heat, trauma, and oxygen-poor blood flow. 14,16
To determine the types of M. longifolia-induced cell death, we used LDH and MTT assays. LDH is the assay originally used to measure cell death occurring via necrosis. 17 A high dosage of M. longifolia (200 μg/mL) induced the loss of SW480 cell membrane integrity. MTT is a common method that is used to determine the cell viability. When the results from MTT and LDH assay are compared, M. longifolia dosages <200 μg/mL induced slight necrosis. The majority of cell death was due to the M. longifolia-induced apoptosis. Thus, to measure the possible cell death, a concentration ≤100 μg/mL should be used in future research.
DNA fragmentation, regarded as a critical process in apoptosis, is the most recognizable hallmark of apoptosis. 18 Chromosomal DNA is cleaved into 50- to 200-kb segments in the early stage of apoptosis and later cleaved into nucleosomal units. 19 In this study, 100 μg/mL of M. longifolia fragmented the total DNA into 100-bp small segments. M. longifolia stimulation of SW480 cells causes DNA fragmentation, thereby inducing the process of apoptosis.
Some reports indicated that activated caspases are required for DNA fragmentation. When apoptotic stimuli attack the cells, caspases, specifically caspase 3, activate an endonuclease caspase-activated DNase (CAD) that is responsible for DNA fragmentation. Caspase 3 then cleaves the inhibitor of CAD to cause DNA degradation. 19 –21
Caspases are synthesized in the cell as inactive precursors, or procaspases. Ordinarily, caspases are selective cleavage of the precursor. Pro-caspases can be triggered by the activation of death receptors on the cell surface, resulting in apoptosis induction. 22 In other words, M. longifolia inhibits the expression of pro-caspases by stimulating death receptors enabling SW480 cells to carry out apoptosis.
PARP is an important nuclear DNA repair enzyme. The cleavage of PARP is considered an early marker for DNA fragmentation and apoptosis. 23 –25 Cleaved PARP appeared in the Western blot. The increased ratio of cleaved-PARP/PARP confirms the DNA fragmentation and apoptosis induction effects of M. longifolia on SW480 cells.
p53 is involved in cellular responses to DNA damage during apoptosis activation. p53 regulates the cell cycle, works as a tumor suppressor, controls proliferative signals in apoptosis, and controls DNA. 26 –28 When M. longifolia induces DNA damage and apoptosis in SW480 cells, the level of p53 protein expression increases. p53-dependent apoptosis is related to the Apaf-1/caspase-9 pathway. 29 The enhanced p53 levels correspond to the attenuated pro-caspase-9 expression observed in Figure 3A.
On activation, p53 increases the transcription of many genes encoding mitochondrial proteins, including cytosolic Bax. 30 Bax is a pro-apoptotic member (Bax, Bad, Bak, and Bok) of the Bcl-2 family that usually interacts with anti-apoptotic Bcl-2 family members (Bcl-2, Bcl-XL, and Mcl-w). In M. longifolia-treated SW480 cells, the levels of Bax and Bad protein expression were increased. In contrast, the level of Bcl-2 protein was decreased in the mitochondria. The ratio between pro- and anti-apoptotic proteins in the Bcl-2 family helps to determine the susceptibility of cells to a death signal. 31 Particularly, the Bax/Bcl-2 ratio plays an important role in mitochondria-related apoptosis and is considered a rheostat to identify apoptotic cells. 32 The Bax/Bcl-2 ratio significantly increased in this study and may contribute toward activating caspase-3 downstream under the apoptosis-inducing effects of M. longifolia.
In conclusion, the present work provides the first evidence for the anticancer effects of M. longifolia in human colon cancer cells. M. longifolia induced apoptotic cell death in SW480 cells through DNA fragmentation, p53 activation, and mitochondria-related apoptosis. M. longifolia upregulated pro-apoptotic Bax and Bad protein expression, downregulated anti-apoptotic Bcl-2 protein expression, inhibited pro-caspase proteins, enhanced the level of caspase-3, and cleaved PARP protein. Moreover, M. longifolia induced apoptosis by increasing the Bax/Bcl-2 ratio. Taken together, the results suggest that M. longifolia may help prevent or treat human colon cancer.
Footnotes
Acknowledgments
This work was supported by a grant (714001-07) from the Research Center for Industrialization of Natural Nutraceuticals through the Agriculture, Food and Rural Affairs Research Center Support Program, Ministry of Agriculture, Food and Rural Affairs.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.