
Abstract
Brassica rapa L. ssp. pekinensis, commonly known as Chinese cabbage, is a cruciferous vegetable traditionally consumed in east Asia. Although its habitual consumption could account for the low incidence of chronic vascular inflammation, the therapeutic and protective potential of phytochemicals derived from Chinese cabbage has been poorly studied. In this study, we identified the phenolic compounds, kaempferol and quercetin, from the ethanol extract of Chinese cabbage (EtCC). We show for the first time that EtCC contains effective phytochemicals that suppress tumor necrosis factor (TNF)-α-induced inflammatory response in human umbilical vein endothelial cells. The EtCC inhibited TNF-α-induced monocyte adhesion to endothelial cells in a dose-dependent manner. The antiadhesive activity of EtCC directly correlated with downregulation of expression and transcription of vascular cell adhesion molecule-1 (VCAM-1). It was caused by an Nrf-2-dependent mechanism, leading to activation of antioxidant responsive element-driven promoter. Taken together, these results suggest that EtCC inhibits the expression of TNF-α-induced adhesion molecules through the indirect transcriptional modulation of VCAM-1 in endothelial cells. In conclusion, regular consumption of vegetables containing dietary phytochemicals might be a potential therapeutic strategy to protect against various stresses, to prevent several pathological conditions, and to treat chronic vascular inflammation, such as atherosclerosis.
Introduction
C
Atherosclerosis is a chronic inflammatory disorder and a leading cause of mortality worldwide. It involves the inflammation inside the wall of large blood vessels that results in vascular endothelial dysfunctions and eventually disruption of blood flow. 11 In early arterial inflammation, the endothelium is activated by various stimuli, including pathogens, oxidized lipids, or cytokines, such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6, which subsequently induce the expression of specific adhesion receptors that enhance circulating monocyte recruitment into the plaque-forming foci. 12 The rolling and arrested monocytes on inflamed endothelium consecutively transmigrate, ensuing diapedesis. 13,14
Extensive epidemiological data, from both in vivo and in vitro atherosclerosis models, support the premise that dietary intake of natural products may improve vascular endothelial dysfunction. 15,16 Popular and edible cruciferous vegetables have been traditionally used to maintain health, and their components have been used in the prevention and treatment of chronic vascular inflammatory diseases. For example, Chinese cabbage has antioxidant properties and is used to make the traditional Korean fermented side dish, kimchi, leading to its consumption in large quantities. Polyphenols in Chinese cabbage have antioxidant and antiplatelet properties, which may be protective against cardiovascular diseases, leading to the stabilization of the atheroma plaque. 17 Isothiocyanates also possess a protective role owing to the regulation of the expression of antioxidant enzymes, such as glutathione transferase and NAD(P)H:quinone oxidoreductase. 18
Although the active compounds of the widely consumed Chinese cabbage have been identified, the molecular targets and the mechanisms underlying their vascular anti-inflammatory effects are poorly studied. In the present study, we used an ethanol extract of Chinese cabbage (EtCC) to gain insights into the physiological mechanisms of its vascular anti-inflammatory activity. We observed that EtCC has an anti-inflammatory effect on TNF-α-stimulated human umbilical vein cells (HUVECs) due to its suppression of the expression of adhesion molecules.
Materials and Methods
Materials
HUVECs and monocyte U937 cells were obtained from Clonetics (Walkersville, MD, USA), and human embryonic kidney (HEK) epithelial 293T cells were obtained from the American Type Culture Collection (#CCL-2, Rockville, MD, USA). Endothelial growth medium, Dulbecco's modified Eagle's medium, fetal bovine serum, and antibiotics were purchased from Gibco (Grand Island, NY, USA). Human TNF-α was purchased from Sigma (St. Louis, MO, USA). Antibodies against vascular cell adhesion molecule (VCAM-1) and β-actin were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA) and Sigma, respectively.
Preparation of ethanol extract
A commercial Chinese cabbage cultivar (Kwonnong Carotene™; Kwonnong Seed Company, Chungju, Korea) cultivated in Fall 2015 was obtained from Kwonnong Seed Company. The Chinese cabbages were stored at −20°C before being subjected to ethanol extraction. After thawing, the samples were crushed using blenders, and the soluble fraction was removed using centrifugation (15,000 g) at 4°C for 20 min. The solid fraction was dispersed in twofold (w/w) amount of 50% ethanol (v/v) and mixed using on overhead stirrer (250 rpm [OS20-Pro, Scilogex, CT, USA]) at room temperature for 15 min. The 50% ethanol extract was obtained after removing insoluble fractions after centrifugation (15,000 g) at 4°C for 20 min. The extract was freeze-dried after being concentrated through vacuum evaporation. Before analyses, the extract was stored at −20°C.
Analysis of phenolic compounds
Phenolic acids (gallic acid, caffeic acid, coumaric acid, trans-ferulic acid, and sinapic acid) and flavonols were analyzed according to the methods described by Jiang et al. 19 with slight modification. Phenolic acids were identified using the five standards, and phenolic acids having no standard molecule were quantified using sinapic acid as the standard molecule. Quercetin and kaempferol were used as standards for flavonol quantification. Total phenolic compounds were assayed according to the method described by Vasco et al. 20
Cell viability assay
Monolayer cultures of HUVECs were maintained as described previously. 21 HUVECs at 80–90% confluence were treated with various concentrations of EtCC (1–1000 μg/mL) and then stimulated with 15 ng/mL of TNF-α. The effect of EtCC on the viability of HUVECs was analyzed using the RealTime-Glo™ MT luminescent kit (Promega, Madison, WI, USA) in an opaque-walled assay plate, according to the manufacturer's instructions. Briefly, the HUVECs were incubated with various concentrations of EtCC (1–1000 μg/mL) for 72 h. To continuously monitor the viability of the treated HUVECs in real time, the prosubstrate and luciferase were added at the same time as EtCC treatment. Luminescence intensity, observable only in metabolically active cells due to reduction of the luciferase substrate, was measured as a marker of viability, on a GloMax luminometer (Promega, Madison, WI, USA). Each sample was analyzed at least in triplicate.
Monocyte–endothelial cell adhesion assay
The monocyte–endothelial cell adhesion assay was performed using suspension cultures of U937 cells, as described previously. 21 Briefly, HUVECs (3 × 104 cells) were seeded in 96-well plates to reach confluence and treated with various concentrations of EtCC, followed by stimulation with 15 ng/mL TNF-α for 18 h. For the quantitative adhesion assay, fluorescent-labeled U937 monocytes were added to the TNF-α-stimulated HUVECs and incubated for a further 2 h. After washing off the unbound U937 three times, monocyte adhesion was measured using a GloMax fluorometer (ex: 485 nm; em: 538 nm). Wells containing HUVECs alone were used as blanks. The bound U937 cells were visualized using a fluorescence microscope (Axiophot; Carl Zeiss, Germany).
Luciferase assay
The effect of EtCC treatment on regulation of VCAM-1 transcription was determined by luciferase reporter assay. HEK293T cells stably having VCAM-1 promoter fused to the firefly luciferase reporter gene (HEK 293T/VCAM-1-luc) were used. 21 The HEK 293T/VCAM-1-luc cells were incubated with various concentrations of EtCC in an opaque-walled 96-well plate (4 × 104 cells/well) for 1 h, followed by TNF-α stimulation and further incubation for 6 h. Subsequently, the cells were treated with luciferin for 30 min at 37°C and the resulting biophotonic reaction was measured using the IVIS® Lumina XRMS imaging system (PerkinElmer, Inc., Waltham, MA, USA) and analyzed using Living Image Software 4.4 (PerkinElmer).
HUVECs in a six-well plate (2 × 105 cells/well) were transfected with antioxidant responsive element (ARE)-luciferase reporter plasmid and pCMV-pRL internal control vector (Promega) at a ratio 40:1 (reporter construct: control vector) using Effectene transfection reagent (Qiagen). The cells were treated with EtCC at the desired concentration for 6 h, followed by stimulation with TNF-α. Then, the cells were washed with ice-cold phosphate-buffered saline (PBS) and harvested in reporter lysis buffer. After centrifugation, 20 μL of supernatant fraction was used for measurement of dual-luciferase activity (Promega) using a luminometer. The luciferase activity was normalized against protein concentration and expressed as relative luciferase activity.
Real-time polymerase chain reaction analysis of adhesion molecule, VCAM-1
TNF-α-stimulated HUVECs were treated with EtCC, and then, RNA was extracted using the Qiagen RNeasy kit (Valencia, CA, USA). cDNA was synthesized using a Maxime RT PreMix kit containing oligo (dT) primers from Intron Biotechnology (Seoul, Korea). The cDNA level was quantified with real-time reverse transcription-polymerase chain reaction (PCR) using an AriaMx Real-time PCR system (Agilent Technologies), and the fluorescence was monitored at each cycle. Cycling parameters were 95°C for 12 min to activate Taq polymerase, followed by amplification for 40 cycles at 95°C for 10 sec, 60°C for 20 sec, and 72°C for 30 sec. A standard curve was constructed using different amounts of cDNAs, and the ΔCt values were calculated. The human primer sequences for VCAM-1 were as follows: forward, 5′-GTTGAAGGATGCGGGAGTAT-3′ and reverse, 5′-TTCATGTTGGCTTTTCTTGC-3′.
Immunoblotting
Cells were treated with the specified concentrations of EtCC and lysed, as previously described. 21 The cell lysate was cleared by centrifugation at 12,000 g for 20 min, and the supernatant was used for immunoblotting. Proteins were resolved on SDS-PAGE and transferred onto a polyvinylidene fluoride membrane. Immunoblotting was performed using anti-VCAM-1 antibody and densitometric analysis was performed by normalization to the β-actin intensity.
Immunocytochemistry
HUVECs were cultured on coverslips, followed by treatment with 300 μg/mL EtCC and stimulation with TNF-α for 3 h. After washing with PBS, the cells were fixed with 4% paraformaldehyde, permeabilized with 0.1% Triton X-100, and then incubated with 5% bovine serum albumin (BSA) in PBS for 45 min. Subsequently, the cells were treated with the anti-Nrf-2 antibody (1:200) at 4°C for 18 h, washed with BSA buffer (0.5% BSA and 0.15% glycine in PBS), and incubated with Alexa Fluor 568-conjugated secondary antibody (1:1000; Molecular Probes) for 1 h. Then, cells were counterstained with Hoechst 33342 for 3 min and visualized under a fluorescence microscope (Axiophot, Carl Zeiss, Germany).
Statistical analysis
Differences in the measured variables between the control and EtCC-treated groups were determined using a one-way analysis of variance followed by Bonferroni's test for multiple comparisons. A P-value of <.05 was considered significant.
Results
Phenolic compounds in the EtCC
The content of phenolic acids in EtCC was ∼100-fold greater than that of flavonols (Table 1). Among flavonols, kaempferol was the major compound identified. The amount of total phenolic compounds was also determined to evaluate the total reducing power of the extract. The content is expressed as gallic acid equivalents and was estimated to be ∼20% (w/w) of the extract (Table 1).
Effect of the EtCC on HUVEC viability
To investigate the effects of EtCC on cell cytotoxicity, we performed a bioluminescent assay. The viability of HUVECs was unchanged after treatment with various concentrations of EtCC, although a slight, nonsignificant decrease was detected on treatment with 1000 μg/mL EtCC (Fig. 1). The effect of EtCC on HUVEC viability was also monitored at 24, 48, and 72 h and showed more than 92% viability on treatment with 1000 μg/mL EtCC at 72 h. These results indicate that treatment of EtCC did not cause nonspecific cytotoxicity or changes in HUVEC morphology. The EtCC treatment preserved HUVEC viability and concentrations of less than 1000 μg/mL can be regarded as safe; therefore, the EtCC was used at concentrations of ≤500 μg/mL in further studies.
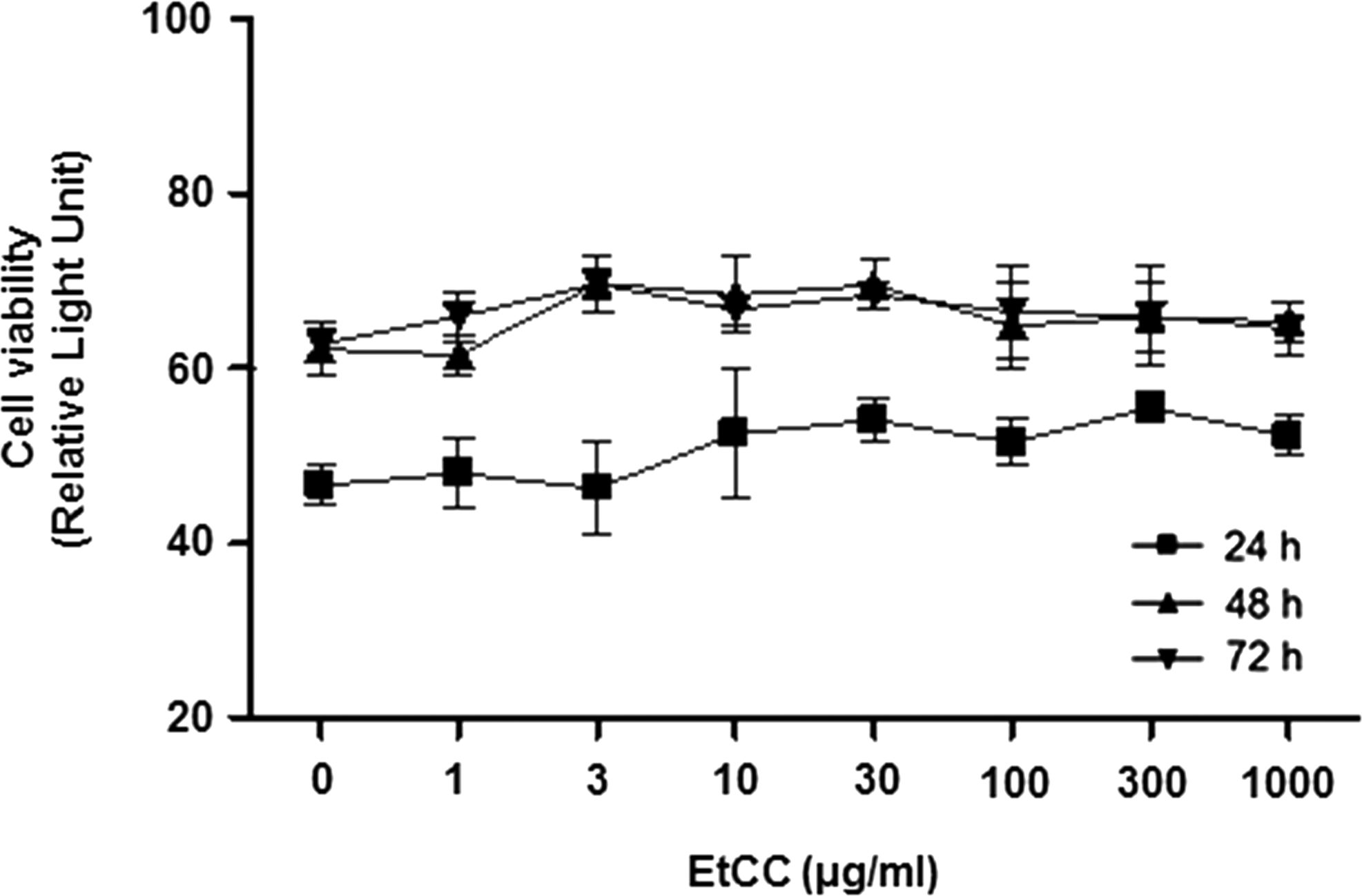
EtCC treatment (1–1000 μg/mL) did not cause decreased viability in HUVECs. The effects of the EtCC on HUVEC viability were determined using a bioluminescent assay and compared after 24, 48, and 72 h of treatment at the indicated concentrations. Data are presented as mean ± SEM (n = 3). Similar results were observed in replicate experiments. EtCC, ethanol extract of Chinese cabbage; HUVEC, human umbilical vein endothelial cells; SEM, standard error of the mean.
EtCC treatment caused a suppression of VCAM-1 expression via transcriptional regulation
The adhesion molecule, VCAM-1, plays an important role in the formation of atherosclerotic lesions by binding to circulating monocytes and increasing their adherence to inflamed surfaces of endothelial cells. We determined the effect of EtCC treatment on VCAM-1 levels to gain insights into the mechanism of EtCC-mediated suppression of inflammation in TNF-α-stimulated HUVECs. As shown in Figure 2A, exposure of HUVECs to TNF-α significantly induced VCAM-1 expression (left panel). However, HUVECs treated with the EtCC before incubation with TNF-α for 18 h showed a marked VCAM-1 suppression, in an EtCC-concentration-dependent manner. For instance, when TNF-α-stimulated HUVECs were treated with 300 μg/mL EtCC for 18 h, the expression level of VCAM-1 was decreased by 50% (Fig. 2A right panel).
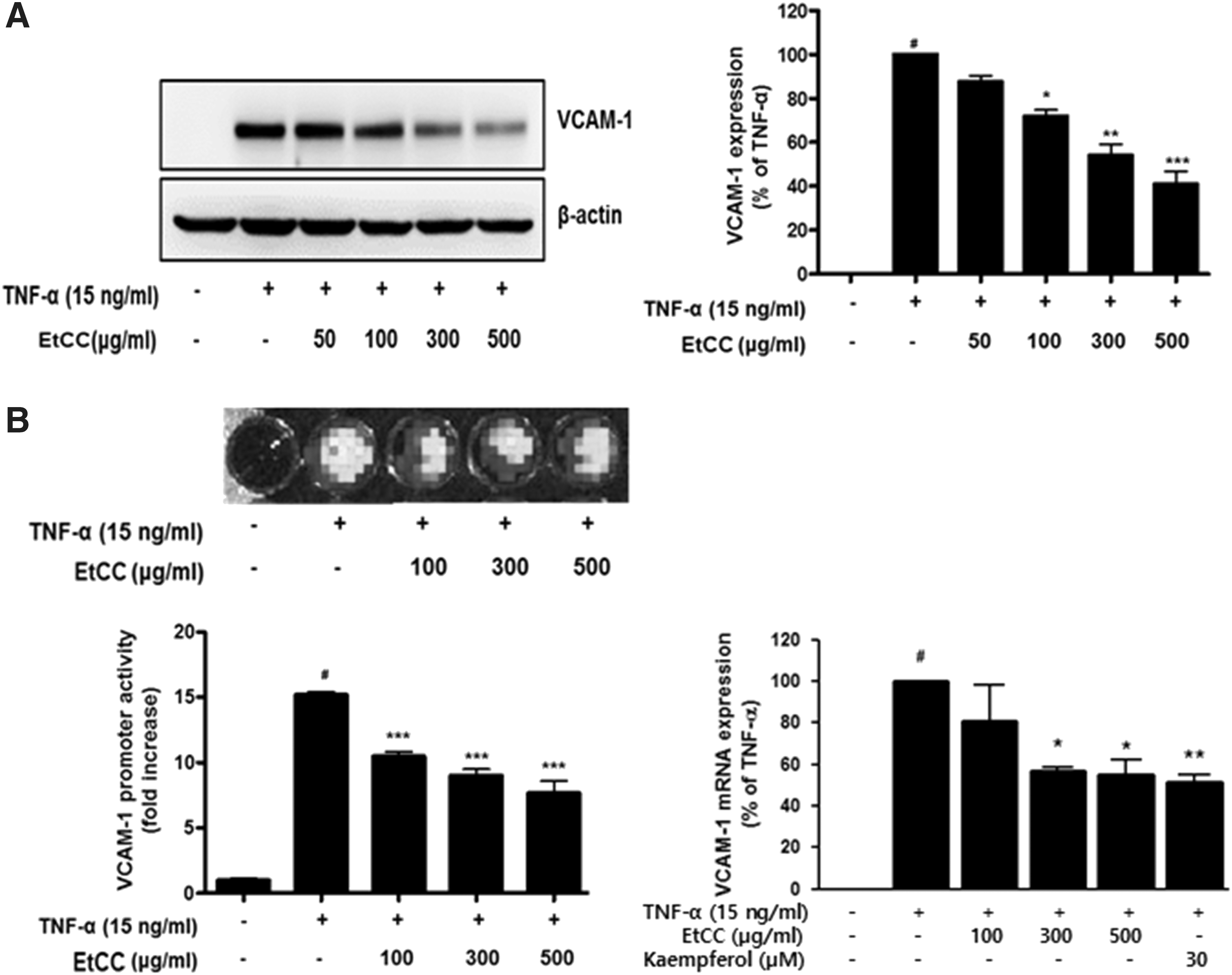
EtCC treatment altered protein levels and transcription of vascular cell adhesion molecule-1 (VCAM-1) in TNF-α-stimulated HUVECs.
Furthermore, to confirm whether the EtCC treatment caused the downregulation of VCAM-1 expression, we analyzed the regulation of transcription of VCAM-1 by using a luciferase reporter assay in HEK 293T/VCAM-1-luc cells, 21 and the transcriptional level of VCAM-1 by real-time PCR. The results are summarized in Figure 2B. Treatment with TNF-α caused an increase in the transcriptional activity of VCAM-1 in HEK293T/VCAM-1-luc cells and the level of VCAM-1 transcript in HUVECs. A remarkable decrease in both transcriptional activity and mRNA level of VCAM-1 was observed in EtCC-treated TNF-α-stimulated cells, in a concentration-dependent manner, similar to the results observed in immunoblot analyses (Fig. 2A). Treatment with 300 μg/mL EtCC caused 40% inhibition in TNF-α-induced VCAM-1 transcriptional activity as shown in relative luciferase activities for VCAM-1 in TNF-α-stimulated HEK293T/VCAM-1-luc cells with representative luminescence images (Fig. 2B left panel). In addition, the VCAM-1 transcript levels in 300 μg/mL EtCC-treated cells decreased by 50%, compared with that in the TNF-α-treated control cells (Fig. 2B right panel). These results indicate that the EtCC-mediated decrease in VCAM-1 expression level was caused by the transcriptional regulation of VCAM-1.
EtCC treatment inhibits monocyte adhesion to TNF-α-stimulated HUVECs
Since the effect of EtCC was most pronounced on VCAM-1 expression in TNF-α-stimulated HUVECs (Fig. 2), we analyzed whether EtCC treatment functionally affected monocyte adhesion to TNF-α-stimulated endothelial cells through the monocyte adhesion assay. HUVECs pretreated with the EtCC were incubated with TNF-α for 18 h, followed by reaction with fluorescent probe-labeled monocytes. TNF-α-induced monocyte adhesion was inhibited by the EtCC in a concentration-dependent manner (Fig. 3). For example, exposure to 300 μg/mL EtCC resulted in a 21.7% decrease in monocyte adhesion compared to that in cells treated with TNF-α alone. The inhibitory action of the EtCC was shown by the fact that the extract itself did not affect monocyte adhesion (data not shown). Fluorescent probe-labeled monocyte adhesion to TNF-α-stimulated endothelial cells was observed using fluorescence microscopy (Fig. 3A upper panel). EtCC treatment induced a significant decrease in the fluorescent intensity in a concentration-dependent manner. The effects of EtCC treatment on monocyte adhesion were well correlated with its effects on VCAM-1 expression. Collectively, the results indicate that EtCC might inhibit the adherence of circulating monocytes to damaged endothelial cell surfaces.
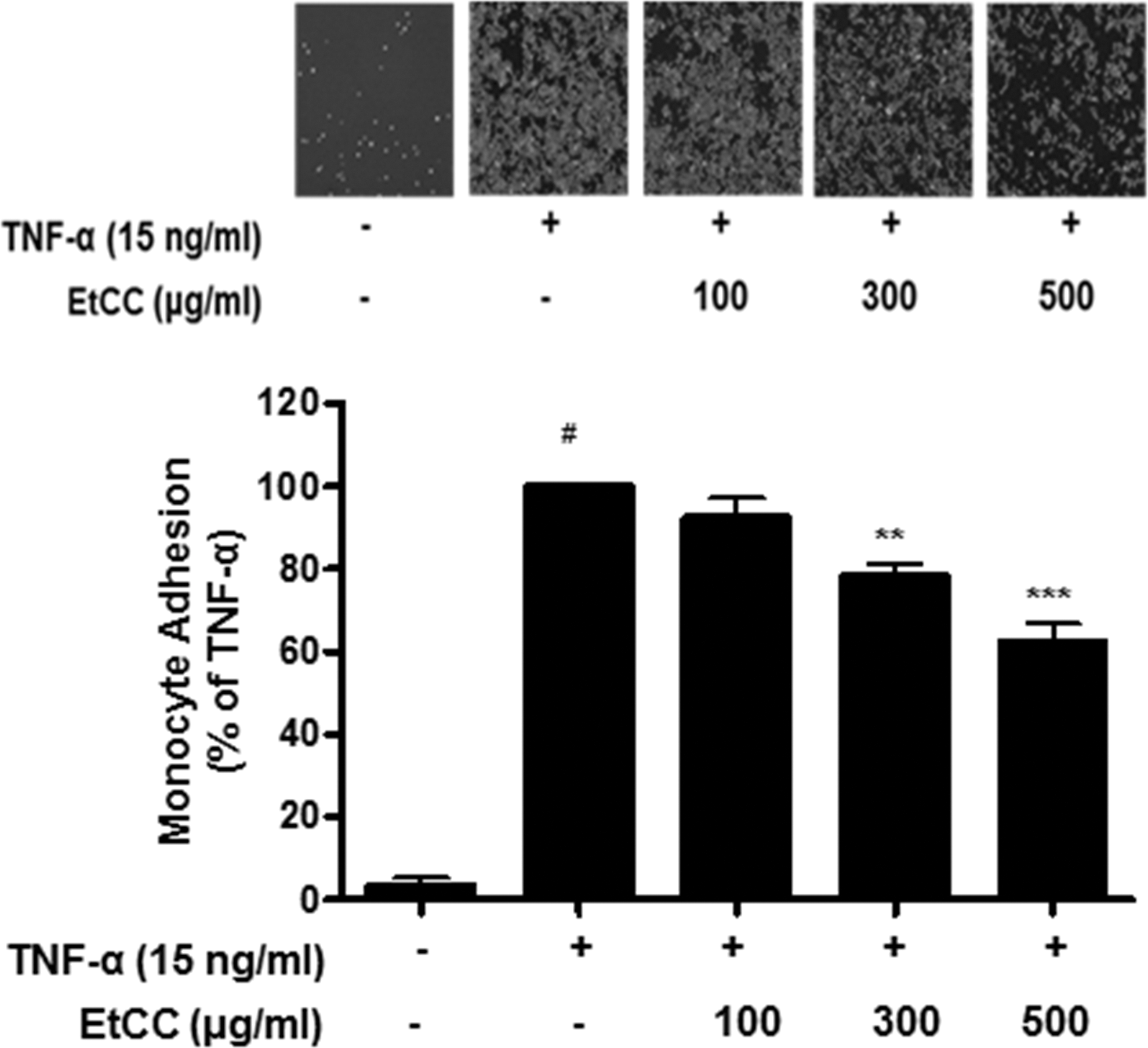
Quantitative adhesion assay of U937 monocytes to TNF-α-stimulated HUVECs. HUVECs were pretreated with EtCC for 1 h, followed by treatment with TNF-α for 18 h. Fluorescence probe-labeled U937 monocytes were added onto TNF-α-stimulated HUVECs and then incubated for 90 min. Adherent U937 cells observed using a fluorescence microscope at × 200 magnification (upper panel). The number of U937 cells adhering to the HUVECs was determined using a fluorometer (lower panel). Data are representative of three independent experiments. Results are presented as mean ± SEM (n = 3). # P < .01 compared with untreated cells and **P < .01, ***P < .001 compared with TNF-α-treated group as determined by one-way ANOVA followed by Bonferroni's multiple comparison tests.
EtCC treatment resulted in suppression of TNF-α-induced VCAM-1 expression through a nuclear factor erythroid 2-related factor 2-dependent mechanism with activation of ARE-driven promoter
Phytochemicals are known as phase II detoxification enzyme inducers and increase ARE-mediated transcriptional activity. EtCC was shown to have potent amounts of phytochemicals such as kaempferol as shown in the HPLC analysis (Table 1). Next, we determined whether ARE activation resulted in VCAM-1 suppression after nuclear factor erythroid 2-related factor 2 (Nrf2) translocation into the nucleus in EtCC-treated HUVECs. Nuclear translocation of Nrf2 was observed in TNF-α-stimulated HUVECs treated with 300 μg/mL EtCC for 3 h. Figure 4A shows that EtCC treatment caused a comparable increase in the nuclear level of Nrf2 in kaempferol-treated cells as a positive control. Interestingly, Nrf2 translocation was observed in response to only TNF-α, resulting in the generation of intracellular reactive oxygen species. This observation may be attributable to the emerging regulatory role of Nrf2 in cellular resistance to oxidants.
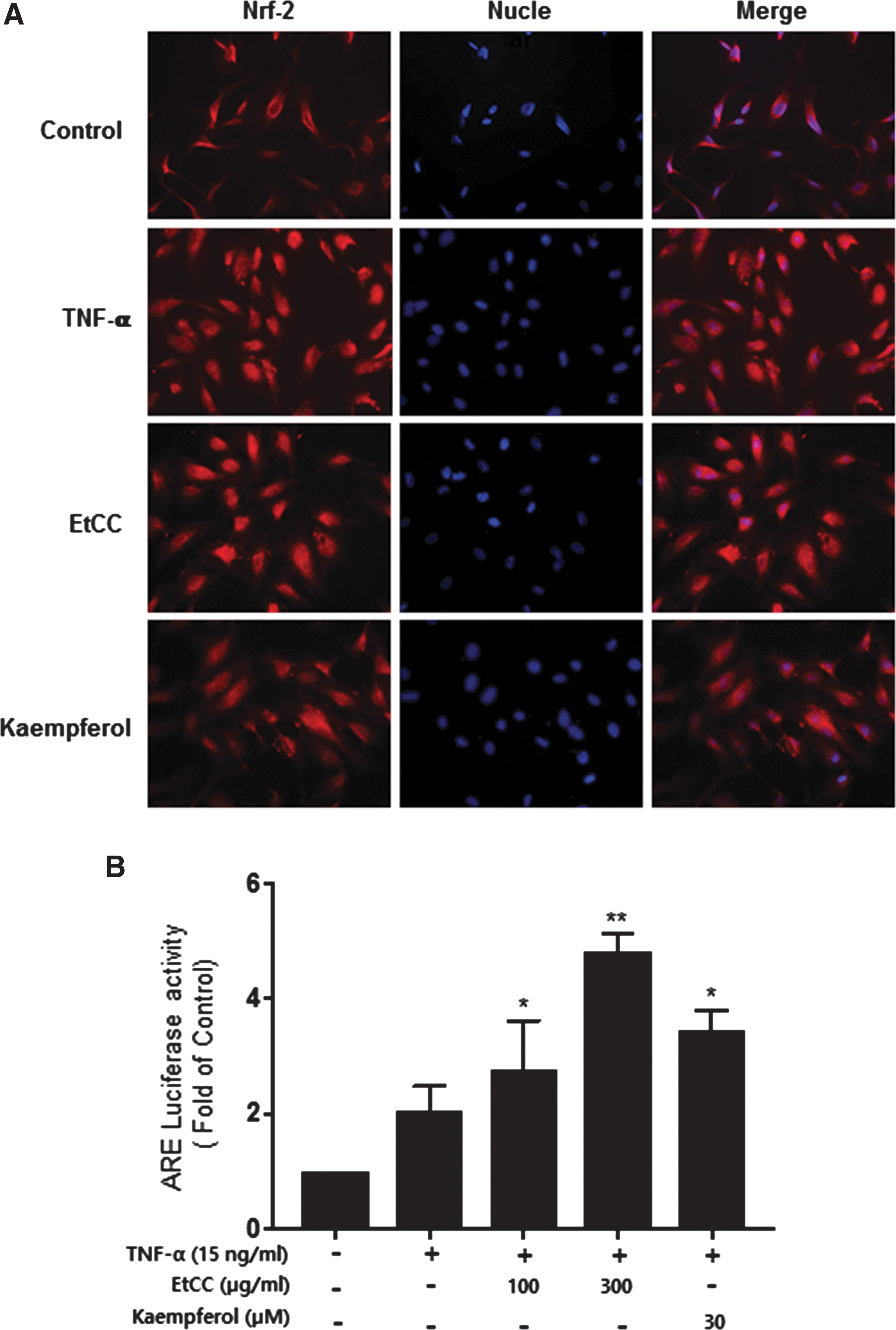
EtCC treatment caused translocation of Nrf-2 from the cytosol to the nucleus in TNF-α-stimulated HUVECs, leading to activation of the ARE-driven promoter.
The responses to EtCC by ARE-driven promoter activity in TNF-α-stimulated endothelial cells, which were transiently transfected with ARE-luc, were analyzed, as a measure of EtCC-induced inhibition of vascular inflammation. Indeed, a remarkable increase in ARE transcriptional activity was observed in EtCC-treated HUVECs in a concentration-dependent manner. The EtCC elicited an increase in ARE transcriptional activity, such that TNF-α-induced initial activation was followed by an ∼2.5-fold increase after a 4-h treatment with 300 μg/mL EtCC, excluding the possibility that the effect was due to the extract itself. Taken together, these data suggest that the Nrf-2/ARE pathway is involved in EtCC-mediated inhibition of vascular inflammation, leading to suppression of VCAM-1.
Discussion
Cruciferous vegetables, including B. rapa spp., are known to possess therapeutic activities against chronic diseases and are increasingly considered as a source of novel and potential biological agents. 22,23 In the present study, we prepared EtCC and analyzed its compositions to identify nutraceutical ingredients from Chinese cabbage, B. rapa L. ssp. pekinensis, and observed anti-inflammatory activity against vascular inflammation. EtCC showed vascular anti-inflammatory activity in TNF-α-stimulated endothelial cells based on the following observations: (a) EtCC plays an effective role with IC50 of 300–500 μg/mL in TNF-α-stimulated HUVECs. (b) EtCC treatment decreased VCAM-1 expression in a concentration-dependent manner due to its transcriptional regulation, leading to suppression of monocyte adhesion to TNF-α-treated HUVECs. (c) EtCC treatment also showed a remarkable increase in ARE-driven promoter activity through an Nrf-2-dependent mechanism. (d) This activity correlated with the known effects of the nutraceutical ingredients present in EtCC: phenolic acid, gallic acid, and two flavonoids, kaempferol and quercetin.
Consumption of vegetables containing nutraceutical ingredients is associated with a lowered risk of cardiovascular disease (CVD), including atherosclerosis. 24 In Korea, a lower prevalence of chronic disease is observed, which may be attributed to the incorporation of various B. rapa spp. in food. Since the 1970s, the incidence rate of CVD has been increasing with changes in socioenvironmental and lifestyle factors, including dietary patterns such as decreases in the intake of plant foods rich in traditional Korean diets and increases in the intake of animal foods. 25 Recent epidemiological studies have also shown that westernized lifestyle and nutritional imbalance are associated with chronic diseases, including obesity, diabetes, cancer, and CVD. 26,27 Many reports describe the biological activities of dietary polyphenols in modulating gene expression for promoting anti-inflammatory mediators. For example, the polyphenols from cruciferous vegetables, resveratrol, attenuate the overexpression of adhesion molecules (VCAM-1 and ICAM-1) through inhibition of the NF-κB pathway in endothelial cells. 28 The anti-inflammatory properties of quercetin were also observed in lipopolysaccharide-stimulated RAW264.7 macrophages owing to NF-κB regulation, thereby decreasing mRNA and protein levels of cytokines and adhesion molecules. 29 In this study, EtCC significantly inhibited monocyte adhesion in TNF-α-stimulated endothelial cells at a dose ≤500 μg/mL, which was within the nontoxic range. These results would explain the relationship between frequent use of cruciferous vegetables and low cardiovascular disorder incidence in Korea.
In this study, we identified the major polyphenols, gallic acid, kaempferol, and quercetin, from EtCC owing to their volatile nature as well as relatively lower proportions and solubility, although B. rapa spp. contains various other phenolic compounds. 1,2,5 Since the adherence of circulating monocytes to the adhesion molecules expressed on the surface of endothelial cells is a critical step in the formation of atherosclerotic lesions, the regulation of the expression of adhesion molecules is a potential pharmacological target for CVD treatments. In this study, we also found that both protein levels and transcription of VCAM-1 were remarkably attenuated by 300 μg/mL EtCC treatment in TNF-α-stimulated HUVECs, causing moderate inhibition of monocyte adhesion. These results indicated that the vascular anti-inflammatory effects of polyphenols from EtCC are, at least in part, mediated by the downregulation of VCAM-1. Many studies also suggest that the anti-inflammatory effect of polyphenols results from direct inhibitory activity on NF-κB, the ubiquitous transcription factor, leading to regulation of cytokines and adhesive molecules. 23,30 However, NF-κB, which regulates proinflammatory genes, was not inhibited by EtCC treatment in TNF-α-stimulated HUVECs (data not shown). We observed no inhibition of p65-NF-κB transcriptional activity in response to EtCC treatment in a reporter gene assay. Thus, the anti-inflammatory signaling of EtCC does not involve the classical NF-κB activation-IκB degradation and nuclear translocation of p65 for suppression of VCAM-1. 31 It may be due to the discrepancies in the phytochemical concentrations used in this observation although none showed cytotoxicity.
Previous reports have indicated that phytochemicals from cruciferous vegetables activate the redox-sensitive transcription factor, Nrf2, which regulates ARE-mediated genes, including anti-inflammatory response proteins. 32,33 Kaempferol, a major flavonol in EtCC, is one of the most potent activators of the Nrf2/ARE pathway and it exerts indirect antioxidant effects. Accordingly, our data showed that EtCC treatment increased ARE-driven transcriptional activity, which affected the EtCC-mediated increase in Nrf-2 translocation within 3 h. This effect could be attributable to the fact that the anti-inflammatory activity of EtCC is mediated more by acute actions than it is by the subsequent activation of Nrf2-dependent genes following longer exposure to EtCC.
In conclusion, these results indicate that the polyphenols from B. rapa spp. regulate TNF-α-induced inflammation, mediated by the suppression of VCAM-1. This study also suggests that frequent consumption of B. rapa spp. is useful for lowering the incidence of chronic inflammatory diseases, such as CVD. Furthermore, cultivation of improved B. rapa spp. with anti-inflammatory activities would be a step toward the development of novel functional foods.
Footnotes
Acknowledgments
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Golden Seed Project, funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA) (No. 213002044SBX20). In addition, it was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2014R1A6A1029617).
Author Disclosure Statement
The authors report no conflicts of interest.