
Abstract
Omega-3 and omega-6 polyunsaturated fatty acids (PUFAs), such as α-linolenic and linoleic acids, are essential fatty acids in mammals, because they cannot be synthesized de novo. However, fat-1 transgenic mice can synthesize omega-3 PUFAs from omega-6 PUFAs without dietary supplementation of omega-3, leading to abundant omega-3 PUFA accumulation in various tissues. In this study, we used fat-1 transgenic mice to investigate the role of omega-3 PUFAs in response to inflammatory pain. A high omega-3 PUFA tissue content attenuated formalin-induced pain sensitivity, microglial activation, inducible nitric oxide synthase expression, and the phosphorylation of NR2B, a subunit of the N-methyl-
Introduction
T
Omega-3 and omega-6 fatty acids are polyunsaturated fatty acids (PUFAs) containing multiple double bonds. They include the omega-6 PUFA linoleic acid and the omega-3 PUFA α-linolenic acid (ALA), both of which serve as precursors in the synthesis of long-chain PUFAs, such as arachidonic acid (omega-6), eicosapentaenoic acid (EPA, omega-3), and docosahexaenoic acid (DHA, omega-3). 9 Among the important omega-3 fatty acids, ALA cannot be synthesized de novo by humans or other mammals and, therefore, must be supplied by the diet. 9
The role of omega-3 PUFAs, especially EPA and DHA, in the prevention and treatment of inflammatory disorders is well known. 10 Among the proposed mechanisms underlying the anti-inflammatory effects of omega-3 PUFAs are the diversion of eicosanoid synthesis to noninflammatory products, the production of anti-inflammatory lipid mediators (resolvins and protectins), the inhibition of nuclear factor κB activation, and the induction of peroxisome proliferator-activated receptor-γ synthesis in dendritic cells. 11 However, although omega-3 fatty acids have been shown to reduce inflammation, their therapeutic benefits in different types of pain have yet to be addressed. 12
The fat-1 transgenic mouse expresses Caenorhabditis elegans fat-1, a gene encoding an omega-3 fatty acid desaturase that enables the endogenous synthesis of omega-3 PUFA from omega-6. 13 These mice have a higher omega-3 PUFA tissue content and exhibit a more robust anti-inflammatory response in models of mucosal organ injury, including acute lung injury, chemically induced colitis, hepatitis, and pancreatitis. 14 The effects on inflammatory pain, however, have been less well studied. In this study, we used fat-1 transgenic mice to examine the effects of increased tissue EPA and DHA concentrations on inflammatory pain, specifically at the spinal cord level.
Materials and Methods
Animals and formalin test
All experiments were carried out with the approval of the Animal Care and Use Committee of Chungnam National University (CNU-00491) and are consistent with the ethical guidelines of the National Institutes of Health and the International Association for the Study of Pain.
Eight-week-old male fat-1 transgenic mice (kindly provided by Dr. J.X. Kang, Harvard Medical School) and C57BL/6 mice were used in this study. All mice were individually housed in cages on a standard 12:12 h light:dark cycle. Water and food were available ad libitum, except during experimentation and during transport to the laboratory ∼1 h before experiments. The presence of fat-1 was confirmed by genotyping. Two groups of mice were studied: wild-type C57BL/6 mice (n = 8) and fat-1 transgenic mice (n = 8). All mice received 10-μL injections of 2% formalin just under the skin of the plantar surface of the right hind paw, which was delivered using a microsyringe with a 26-gauge needle. Mice were then placed in study chambers where dodging, shaking, or flinching events were counted in 5 min interval for 60 min. All measurements were performed between 11 am and 3 pm in a quiet room.
Immunohistochemistry
Immunohistochemistry was performed 1 h after formalin injection. Mice were anesthetized with intraperitoneal sodium pentobarbital (50 mg/kg) and transcardially perfused with heparinized phosphate-buffered saline (PBS, pH 7.4), followed by perfusion with 4% paraformaldehyde for 15 min. Lumbar segments (L3–L5) of the spinal cord were immediately removed, immersed in the same fixative overnight, and then embedded in paraffin. Four-micrometer sections were deparaffinized and rehydrated in a graded alcohol series. The antigen was retrieved using 0.01 M citrate buffer (pH 6.0) by heating the sample in a microwave vacuum histoprocessor (RHS-1, Milestone) at a controlled final temperature of 121°C for 15 min. For immunohistochemical analyses, endogenous peroxidase activity was blocked using 0.3% hydrogen peroxide. Sections were treated with Protein Block solution (Dako) for 20 min and then incubated with specific antibodies targeting iba-1 (1:400, No. 019-19741; Wako), inducible nitric oxide synthase (iNOS) (1:500, No. n32030; Transduction Laboratories), and phospho-p44/42 mitogen-activated protein kinases (MAPKs) (Thr202/Tyr204 Erk1/2, phosphorylated extracellular signal-regulated kinase [p-ERK], 1:200, No. 9101; Cell Signaling). After washing with PBS, tissues were incubated with biotinylated antirabbit IgG and streptavidin peroxidase complex (Vector Laboratories). Immunostaining was observed with diaminobenzidine and specimens were mounted using Polymount.
Western blots
After behavioral testing, lumbar spinal dorsal horn tissue (n = 8 per group) was rapidly dissected. Tissue protein extracts were prepared and immunoblot analysis was performed using antibodies targeting: phosphorylated NR2B (p-NR2B) (1:5000), NR2B (1:10,000), total ERK (t-ERK) (1:1000), p-ERK (1:1000), and β-actin (1:5000). All antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Immunolabeled proteins were detected through chemiluminescence using a Supersignal ECL kit (Pierce Chemical). Western blots were subjected to different exposures during development, ranging from 10 sec to 10 min, and the best exposures were selected for data presentation. Densities of immunoblot bands were quantified using ImageJ software.
Statistical analyses
Statistical significance of differences between the mean values was determined using a two-way repeated measures analysis of variance with a post hoc test or a Mann–Whitney U test. The statistical software package SigmaStat (Systat, San Jose, CA) was used for all statistical analyses. All data are presented as mean ± standard error of the mean. Results were considered significant at P < .05.
Results
Effects of increased omega-3 PUFA on formalin-induced inflammatory pain
The pain responses of the mice were evaluated using formalin injections as a model of chemically induced inflammatory pain. 15 After the subcutaneous injection of diluted formalin (2%, 10 μL) into the plantar surface of the mouse right hind paw, an immediate spontaneous behavioral response was observed, characterized by licking, lifting, and flinching of the injected paw. The behavioral pain response was quantified as the number of licks/lifts/flinches per min during each phase. During the first phase (0–10 min) of this response, fat-1 and wild-type mice did not differ significantly (Fig. 1B, C), whereas during the second phase (10–60 min), the pain response was significantly less in fat-1 mice than in the wild-type control mice (Fig. 1B, C; P < .001). Since the second-phase response reflects central sensitization to pain in the spinal cord, 16 we hypothesized that endogenous omega-3 PUFAs play an important role in the sensitization to chemically induced pain.

Formalin-induced nociceptive behavior.
Effects of increased omega-3 PUFA on microglial activation in the spinal dorsal horn
The effects of increased endogenous omega-3 PUFA content on central sensitization in the spinal cord were determined by examining microglial activation in the spinal dorsal horn of fat-1 and wild-type mice, because microglial cells appear to play a vital role in the initiation of chronic pain. 17 Expression of the microglia-specific protein iba-1 was detected in the lumbar dorsal horn (L3–L5) of both groups, particularly in the superficial dorsal horn (Fig. 2). This region corresponds to areas receiving afferent inputs from the dorsal surface of the hind paw. 18
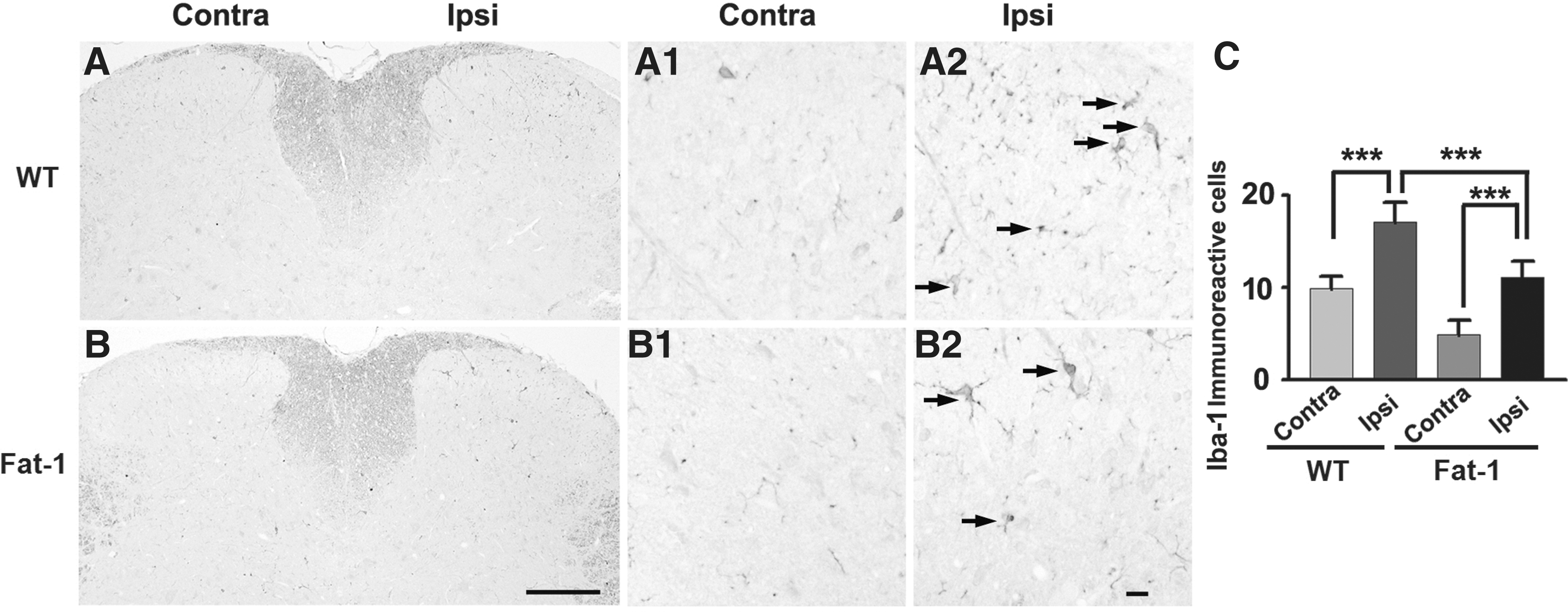
Expression of iba-1 in the lumbar spinal dorsal horn after formalin injection.
Spinal microglia in wild-type mice exhibited a morphology characteristic of activation, including an increase in cell body size and thicker processes (Fig. 2A). By contrast, the spinal microglia of fat-1 mice had thin and highly ramified processes (Fig. 2B). The number of iba-1 positive cells in the ipsilateral lumbar spinal dorsal horn was significantly higher in wild-type mice than in fat-1 mice (Fig. 2C; P < .001). In both groups, there were fewer iba-1-immunoreactive cells on the side contralateral to the formalin injection (Fig. 2C). These results show that iba-1 expression is blunted in fat-1 mice after formalin injection, suggesting that the inhibition of microglial activation alters inflammation-induced behavioral responses.
Effects of increased omega-3 PUFA on iNOS expression in the spinal dorsal horn
NOS, a key enzyme in nitric oxide (NO) production, modulates the response to inflammatory pain. 19 We thus examined the effects of increased endogenous omega-3 PUFA concentrations on iNOS expression in the spinal dorsal horn of wild-type and fat-1 mice after unilateral formalin injection. iNOS expression was detected in the lumbar spinal dorsal horn of wild-type (Fig. 3A) and fat-1 (Fig. 3B) mice, but it was downregulated in the latter compared with the controls (Fig. 3C). The number of iNOS-immunoreactive cells was significantly higher on the ipsilateral side of wild-type mice than that of fat-1 mice (Fig. 3C; P < .01). These findings suggest that an increase in omega-3 PUFA content inhibits iNOS expression, possibly leading to a decrease in formalin-induced inflammatory pain.
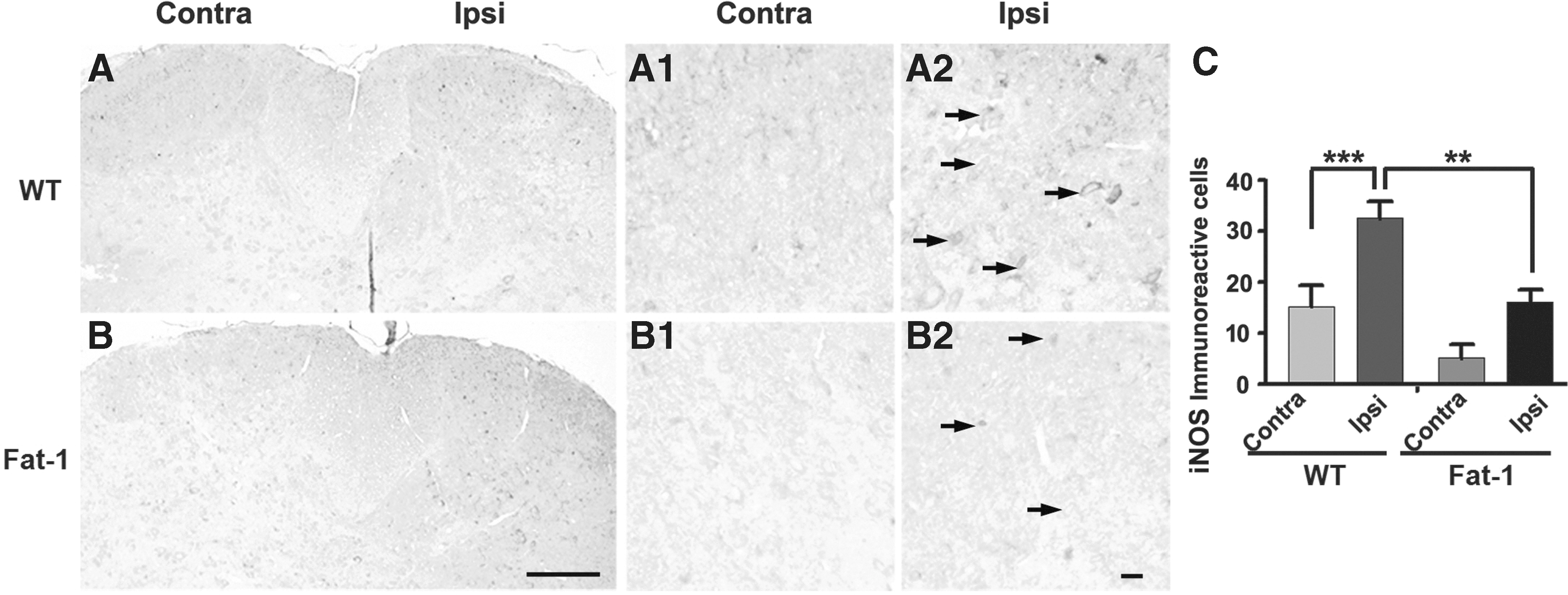
Expression of iNOS in the lumbar spinal dorsal horn after formalin injection.
Effects of increased omega-3 PUFA on ERK and NR2B expression in the spinal dorsal horn
MAPKs, such as ERK, are activated by inflammatory mediators, including those in primary and secondary sensory dorsal horn neurons.
20
These kinases participate in the generation and maintenance of inflammatory pain.
21
N-methyl-
Formalin injection did not alter the expression of t-ERK or total NR2B in fat-1 mice compared with the levels in the wild-type controls (Fig. 4A). However, the expression of p-ERK and p-NR2B in the ipsilateral dorsal horn was lower in fat-1 mice than in wild-type mice (Fig. 4A, F). The ratios of p-ERK to t-ERK (Fig. 4B) and of p-NR2B to total NR2B (Fig. 4C) on the side ipsilateral to formalin injection were significantly lower in fat-1 mice than in the controls (P < .05 for both). The p-NR2B/total NR2B ratio between the contralateral and ipsilateral sides increased significantly only in wild-type mice (Fig. 4C). The ratios of c-Fos to β-actin (Fig. 4D; P < .01) and of p-CREB to β-actin (Fig. 4E; P < .001) on the side ipsilateral to formalin injection were significantly lower in fat-1 mice than in the controls. The p-CREB/β-actin ratio between the contralateral and ipsilateral sides increased significantly only in wild-type mice (Fig. 4E; P < .001). Taken together, these findings suggest that decreased levels or the blunted expression of p-ERK and p-NR2B contributes to the amelioration of the inflammatory pain response.
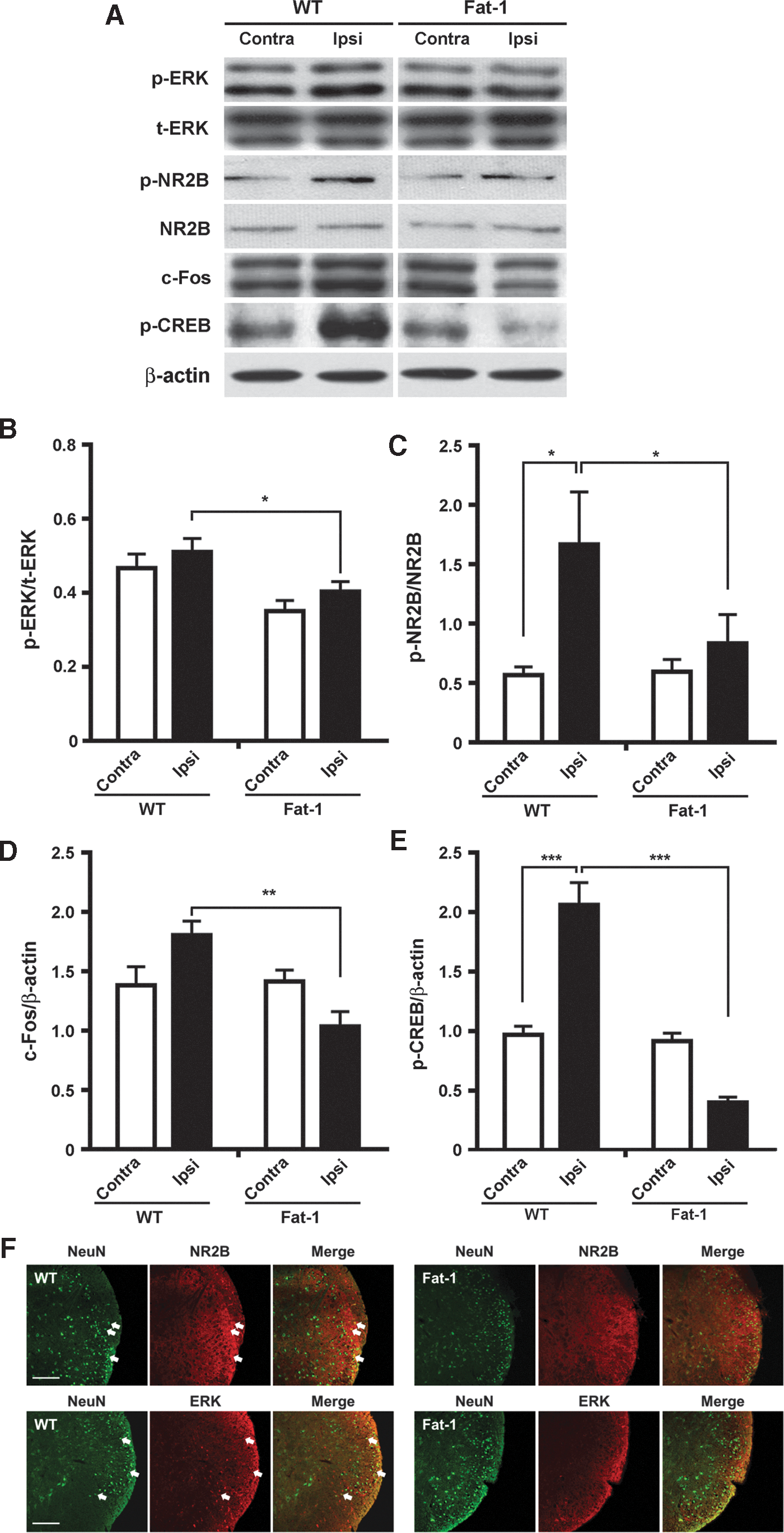
Western blots and analysis of p-ERK, t-ERK, p-NR2B, and total NR2B expression in the lumbar spinal dorsal horn after formalin injection.
Discussion
To the best of our knowledge, this study is the first to demonstrate that omega-3 PUFAs reduce formalin-induced inflammatory pain in fat-1 transgenic mice, presumably by increasing tissue DHA and EPA concentrations. Increased omega-3 PUFA content in mice blunted microglial activation and the expression of iNOS and p-NR2B in the lumbar spinal dorsal horn. Collectively, these findings suggest that increases in omega-3 PUFA content inhibit the expression of proteins in the spinal dorsal horn that are involved in inflammatory pain, leading to attenuation of the pain response.
Omega-3 PUFAs are thought to mediate antinociceptive processes, in part, by inhibiting the microglial release of matrix metalloproteinases. 23 Microglial cells play a vital role in the initiation of chronic pain 17 and glial activation can be induced by C-fiber nociceptive signaling through the sciatic nerve. Data from rodent models suggest that nociceptive glial activation is crucial in acute and inflammatory pain. 24 In lipopolysaccharide-stimulated RAW 264.7 macrophages, pretreatment of the cells with omega-3 significantly attenuated tumor necrosis factor (TNF)-α production. 25 Clinical studies have demonstrated the benefits of omega-3 PUFAs in the management of patients with neuropathic pain, 26 in which activation of spinal cord glial cells is a crucial component. 27 The activated cells are characterized by proliferation, hypertrophy, and the increased production of inflammatory cytokines, such as interleukin (IL)-1β, IL-6, and TNF-α. EPA and DHA may limit the production of these cytokines. Indeed, although neuropathic pain differs from inflammatory pain, the results of this study show that omega-3 PUFAs reduce microglial activation in mice, consistent with a role for microglia in inflammatory pain.
Inhibition of iNOS attenuates inflammatory pain, 28 but the molecular mechanisms mediating this process have yet to be defined. NO is generated in significant concentrations at sites of inflammation where multiple hyperalgesic inflammatory mediators, such as cytokines, prostaglandin E2, and 5-HT, are also produced. 29 NO may facilitate hyperalgesia induced by these mediators through the cyclic adenosine monophosphate second messenger pathway, in addition to exerting cyclic guanosine monophosphate-dependent hyperalgesic effects. In this study, the downregulation of iNOS expression in the ipsilateral lumbar spinal dorsal horn of fat-1 mice versus wild-type mice suggested that omega-3 PUFAs decrease NO production by inhibiting iNOS, potentially resulting in decreased inflammatory pain.
NR2B, a subunit of the NMDA receptor, has been implicated in modulating pain perception. Studies have shown that NMDA receptors contribute to chronic pain after nerve and tissue injury. 30 Peripheral inflammation increases the expression of NR2B-containing NMDA receptors and enhances neurotransmitter release in the anterior cingulate cortex. 31,32 Conversely, the inhibition of NR2B-containing receptors through selective antagonists, either locally in the anterior cingulate cortex or systemically, inhibits inflammation-associated allodynia. 31 Subcutaneous formalin injection produces an immediate increase in glutamate and aspartate in the spinal cord, whereas the induced pain is ameliorated in response to the administration of antagonists either peripherally or in the spine. 33 Taken together, these observations are consistent with a critical role for NMDA receptors in pain transmission. Support for this hypothesis comes from this study. Specifically, the decreased NR2B expression in fat-1 mice after formalin injection suggested that the inhibition of NR2B-containing receptors by omega-3 PUFAs alters inflammatory pain-induced behavioral responses.
Previous studies demonstrated that MAPKs, such as ERK, p38, and c-Jun N-terminal kinase (JNK), contribute to pain sensitization after tissue or nerve injury through distinct molecular and cellular mechanisms. 20 ERK mediates intracellular signaling in response to a variety of stimuli. In nociceptive neurons of the spinal dorsal horn, ERK is phosphorylated in response to axotomy, electrical stimulation of the peripheral nerve, noxious stimuli, and peripheral inflammation. 20 In this context, phosphorylation plays an important role in central pain sensitization in chronic inflammatory and neuropathic pain, by regulating glutamate receptor and potassium channel activity, and the induction of gene transcription. 20 These studies link the molecular effectors regulating ERK phosphorylation to nociception. Similarly, we showed that ERK is phosphorylated in the ipsilateral spinal dorsal horn after formalin injection, but to a much lesser extent in fat-1 mice than in wild-type mice. Our results indicate that omega-3 PUFAs inhibit inflammatory pain by regulating ERK phosphorylation in the spinal dorsal horn.
Recently, Lu et al. 34 investigated whether DHA, a predominant omega-3 PUFA in the central nervous system, could reduce inflammatory pain and inhibit neuroinflammation in the spinal cord after carrageenan injection in mice. They demonstrated that intrathecal injection of DHA prevents and reverses inflammatory pain. Furthermore, DHA treatment decreased carrageenan-induced activation of microglia, phosphorylation of p38 MAPK, and production of proinflammatory cytokines (TNF-α and IL-1β) in the L4–L5 spinal cord. Taken together, their results provided evidence that DHA has an antinociceptive effect in inflammatory pain, which may be attributed to, at least partially, the suppression of microglia-mediated inflammatory response through inhibition of p38 MAPK activation, consistent with our results suggesting a role for microglia and MAPK in inflammatory pain despite direct treatment with DHA into intrathecal space.
Glutamate-mediated activity through NMDA receptors is associated with the activation of p38, ERK, and JNK in microglia. 35 Inflammatory conditions in which proinflammatory cytokines such as IL-1, TNF, and IFN-γ are produced induce the expression of iNOS and production of NO, 36,37 enhancing phosphorylation of NMDA receptors in the spinal cord neurons through inhibition of the ERK signaling pathway. 38 Importantly, these alterations might be associated with the promotion of pain hypersensitivity, suggesting the crucial role of spinal NMDA receptors in microglia activation.
In conclusion, our study provides evidence of the ability of omega-3 PUFAs to modulate inflammatory pain transmission by regulating signal transmission at the spinal dorsal horn. Further studies are required to identify the molecular mechanisms of EPA- and DHA-mediated pain modulation.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT, and Future Planning, and by the Korea Government (MSIP) (2014R1A1A1038222, 2016R1A2B4009409, and 2016R1C1B1015811).
Author Disclosure Statement
No competing financial interests exist.