
Abstract
Plant sterols in their free forms are known to inhibit colon cancer. Whether these activities persist when compounds are incorporated into processed food is not reported yet. This study aimed to test the ability of plant sterol esters (PSE) incorporated into a nonpuffed extruded food (NPE) model to inhibit colon carcinogenesis. PSE was added into NPE at four concentrations (0.0%, 0.7%, 1.4%, and 2.1%). PSE-NPE activity was tested in azoxymethane/dextran sodium sulfate-induced Balb/c mice. The groups given PSE-NPE did not show any colon tumor formation. Immunohistochemistry results revealed that the group fed with 1.4% PSE had the lowest histoscore for cyclooxygenase-2 expression and the highest histoscore for cleaved caspase-3, cleaved caspase-8, and cleaved caspase-9expressions. The results of this study indicated that even after incorporation into a food system, which is processed using high pressure and temperature, PSE retained its chemopreventive activity. The proposed mechanisms are by suppressing inflammation and inducing apoptosis.
Introduction
C
Nutrition is an important factor that is widely known to have a strong link with the disease. 3
One of bioactive compounds reported to have a role in multiple pathways of cancer prevention is plant sterols. Plant sterols are plant-derived dietary compounds with similar structure and basic functions as cholesterol in animals, except for substitutions on the sterol side chain at the C24 position. 4 Plant sterols are found naturally in plant foods as a mixture of β-sitosterol (65%), campesterol (32%), and stigmasterol (3%). 5 Plant sterols were reported to lower blood cholesterol levels by competing with cholesterol loading to micelles. They have a more hydrophobic moiety than cholesterol, and are thus easier to absorb. 6
Plant sterols were also reported to inhibit several types of cancer, including lung, stomach, ovary, breast, and colon cancer. 7 –11 The anticancer mechanisms of plant sterols were proposed to be mediated through decreased carcinogen production, suppression of cell growth and multiplication, inhibition of angiogenesis and metastasis, and were also reported to be associated with an increased number of cancer cells that undergo apoptosis. 12
In the case of colon cancer, the proposed mechanisms were through cancer cell proliferative inhibition and apoptosis stimulation. Plant sterols were reported to suppress the expression of some proliferative markers (β-catenin and PCNA) in colon carcinogenesis. 13 Plant sterols also increased the activity of essential enzymes for apoptosis (caspase-3, caspase-8, and caspase-9). 14,15 The apoptosis effect was also due to the increment of proapoptotic protein Bax, the release of cytochrome c from mitochondria, and DNA fragmentation. 13,14,16
Apoptosis could be inhibited by higher expression of cyclooxygenase-2 (COX-2). 17 COX-2 is one of the enzymes involved in catalyzing arachidonic acid into products of prostaglandin, prostacyclin, and thromboxane. 18 COX-2 has a key role in the generation of inflammatory response and in proliferative diseases, such as cancer. 19,20 COX-2 expression levels were reportedly increased by 85% in the case of adenocarcinoma. 17
Data on the anticancer activity of plant sterol esters (PSE) when it is incorporated into complex food matrices are still lacking. Most previous studies reported the colon cancer chemoprevention activity of pure plant sterol(s) in its free form, without being incorporated into food products. In this study, PSE were incorporated into nonpuffed extruded food (NPE) made by a hot extrusion method to investigate the activity of PSE in food model after processing. Effect of heating and pressure involved during the process, and also interaction between PSE and other components during processing could possibly affect PSE bioactivity. Chemopreventive activities of plant sterol ester-incorporated nonpuffed extrudates (PSE-NPE) were therefore tested in vivo using azoxymethane/dextran sodium sulfate (AOM/DSS)-induced Balb/c mice. AOM/DSS combination was used to develop colitis-related colon carcinogenesis in rodents. AOM is a colonic genotoxic carcinogen that is widely used for colon carcinogenesis investigation, whereas DSS is a synthetic sulfate polysaccharide (nongenotoxic carcinogen) that is extensively used to produce colitis. 21 The expression of several enzymes responsible for inflammation and apoptosis, including COX-2, cleaved caspase-3, cleaved caspase-8, and cleaved caspase-9, was observed by immunohistochemistry (IHC) staining techniques.
Materials and Methods
Materials
The PSE used in this study was Vegapure® 95 FF (BASF, Germany) (min. PSE content 91%). It is a mixture of brassicasterol, campesterol, campestanol, stigmasterol, β-sitosterol, β-sitostanol, D5-avenasterol, D7-stigmastenol, and D7-avenasterol. Sorghum (Kawali cultivar) was kindly provided by PTPN XII Indonesia. Red palm oil was generously provided by PT Sinar Mas Indonesia. Soybean was purchased from a local market in Bogor. Rice bran was prepared in our laboratory from paddy (Ciherang cultivar), which was bought from local farmers in Bogor.
Plant sterol esters incorporation into nonpuffed extrudates
Four nonpuffed extrudates were developed with PSE content of 0%, 0.7%, 1.4%, and 2.1%, respectively (Fig. 1). These concentration ranges were based on the report of cell proliferative inhibition in murine colon by plant sterol. 22 Sorghum, soybean, rice bran, and red palm oil were reported as rich sources of plant sterols. 23 –26 PSE and all base ingredients were mixed in a pilot scale mixer for 3 min, fed through a twin screw extruder (Berto BEX-DS-2256) at 95°C, and the resulting extrudates were dried in a drying oven (80–85°C, 1.0–1.5 h).

Pictures of nonpuffed extruded food (NPE) with plant sterol esters (PSE) enrichment of 0%
Animal and experimental design
Forty-eight male Balb/c mice (6–8 weeks), body weight 21.7 ± 0.3 g approximately, were individually caged and were acclimatized for 10 days before the experiment. All mice were divided into six groups, C− (negative control group), C+ (positive control group), and four PSE-NPE groups. Experimental diets were prepared based on AIN-93 M semipurified rodent diet with modification to contain an 8% fat content. 27 Cornstarch was used as the carbohydrate source of control groups, whereas each of PSE-NPEs was used for each PSE-NPE groups.
Colon cancer was induced in all groups (except C−) by a single intraperitoneal injection of AOM (A5486; Sigma Aldrich) with the dose of 10 mg/kg body weight, whereas C− group was injected by 0.9% NaCl solution. One week after injection, all groups, except C− were given 1% DSS (40,000 MW, J63606, Alfa Aesar) in drinking water for 4 days. After 24 weeks, all mice were sacrificed by cervical dislocation. All animal experimental procedures had been reviewed and been accepted by the Animal Ethics Committee of Veterinary Faculty, Bogor Agricultural University, Indonesia (006/KEH/SKE/VI/2014).
Analysis of COX-2 and cleaved caspase-3,-8,-9 expressions
The colon was Hematoxylin and Eosin (H&E) stained for macroscopic lesion observation before COX-2 and cleaved caspase-3,-8,-9 analyses. The severity of mouse colon changes was determined by scoring: 0 (normal colonic mucosa without inflammation); 1 (villi shortened and slight inflammation); 2 (hyperplasia); 3 (mild-grade dysplasia); 4 (high-grade dysplasia); and 5 (adenocarcinoma).
Analysis of COX-2 and cleaved caspase-3,-8,-9 expressions was conducted using IHC technique, 28 by using monoclonal antibody of rabbit anti-human COX-2 (12282; Cell Signaling Technology), rabbit anti-human cleaved caspase-3 (9579; Cell Signaling Technology), rabbit anti-human cleaved caspase-8 (9429; Cell Signaling Technology), and rabbit anti-human cleaved caspase-9monoclonal antibody (7237; Cell Signaling Technology). Evaluation of expression levels was conducted by scoring the brown color formation and distribution. 29 Histoscore was obtained by multiplying the scores of brown color intensity and color distribution.
Analysis of phytochemical composition of plant sterol ester-incorporated nonpuffed extrudates
Plant sterol
Analysis was conducted by using high-performance liquid chromatography (HPLC) (Agilent Technologies 1200 series) with ultraviolet (UV) detector and silica column. 30 Hexane:isopropanol (9:1) were used as mobile phase and the flow rate was 1.0 mL/min. Sample was extracted by Soxtec extraction and was prepared for saponification before analysis. 31
Soluble fiber
Both soluble and insoluble dietary fiber content was analyzed by the gravimetric enzymatic method. 32 Total dietary fiber was calculated by totaling the soluble and insoluble dietary fiber.
α-Tocopherol
α-tocopherol was determined by using HPLC (Agilent Technologies 1200 series) with C-18 column and UV detector. 33 A mixture of isopropanol:hexane (98:2) was used as mobile phase and the flow rate was 1.0 mL/min.
γ-Oryzanol
Analysis of γ-Oryzanol was determined by using HPLC (Agilent Technologies 1200 series) with C-18 column and UV detector. 34 A mixture of methanol:acetonitrile:dichloromethane:acetic acid (50:44:3:3) was used as mobile phase and the flow rate was 1.0 mL/min.
Total flavonoid and total phenolic content
Total flavonoid and total phenolic content were performed using aluminium chloride method 35 and Folin–Ciocalteu method, 36 respectively.
Statistical analysis
Differences among group means were determined by one-way analysis of variance and post hoc Duncan's multiple range test (P < .05).
Results and Discussion
Effect of plant sterol ester-incorporated nonpuffed extrudates supplementation to the mice health
PSE-NPE addition into mouse diets did not affect the average of food intake, % body weight increment, and relative weight of liver (Table 1). The PSE-NPE groups' kidney relative weight were similar to C+ group, which was higher than C− group (Table 1). The kidney weight alterations of C+ and PSE-NPE groups might reflect the renal toxicity caused by AOM and DSS induction. Based on histopathology profiles, the representative PSE-NPE group showed better improvements in liver and kidney profiles as compared with C+ group (Fig. 2). It had lower liver inflammation and necrosis, fatty degeneration, as well as lower kidney inflammation and hemorrhagic level as compared with C+ group. More detailed data for all groups are presented in Supplementary Figure S1 (Supplementary Data are available online at

Histopathology of liver and kidney of mice group (H&E staining, 1000 × magnification).
C−, negative control group; C+, positive control group; R, group with 0% PSE-NPE; R1, group with 0.7% PSE-NPE; R2, group with 1.4% PSE-NPE; R3, group with 2.1% PSE-NPE. Data are expressed as mean ± SD. Same superscript in the same row indicates no statistical difference (P < .05). Data are average of 6 (1), 10 (2), and 9 (3) replications. Data are shown as 103 (+).
All of the PSE-NPE groups had significantly lower colon relative weights than C− and C+ groups (Table 1). Visual appearance of the PSE-NPE group's colon seemed normal when compared with that of C− group, whereas any tumor or suspected mass was found in C+ group's colon (Supplementary Fig. S2). H&E staining revealed that the PSE-NPE groups had similar colon structure without any tumor formation as compared with C− group (Fig. 3A).
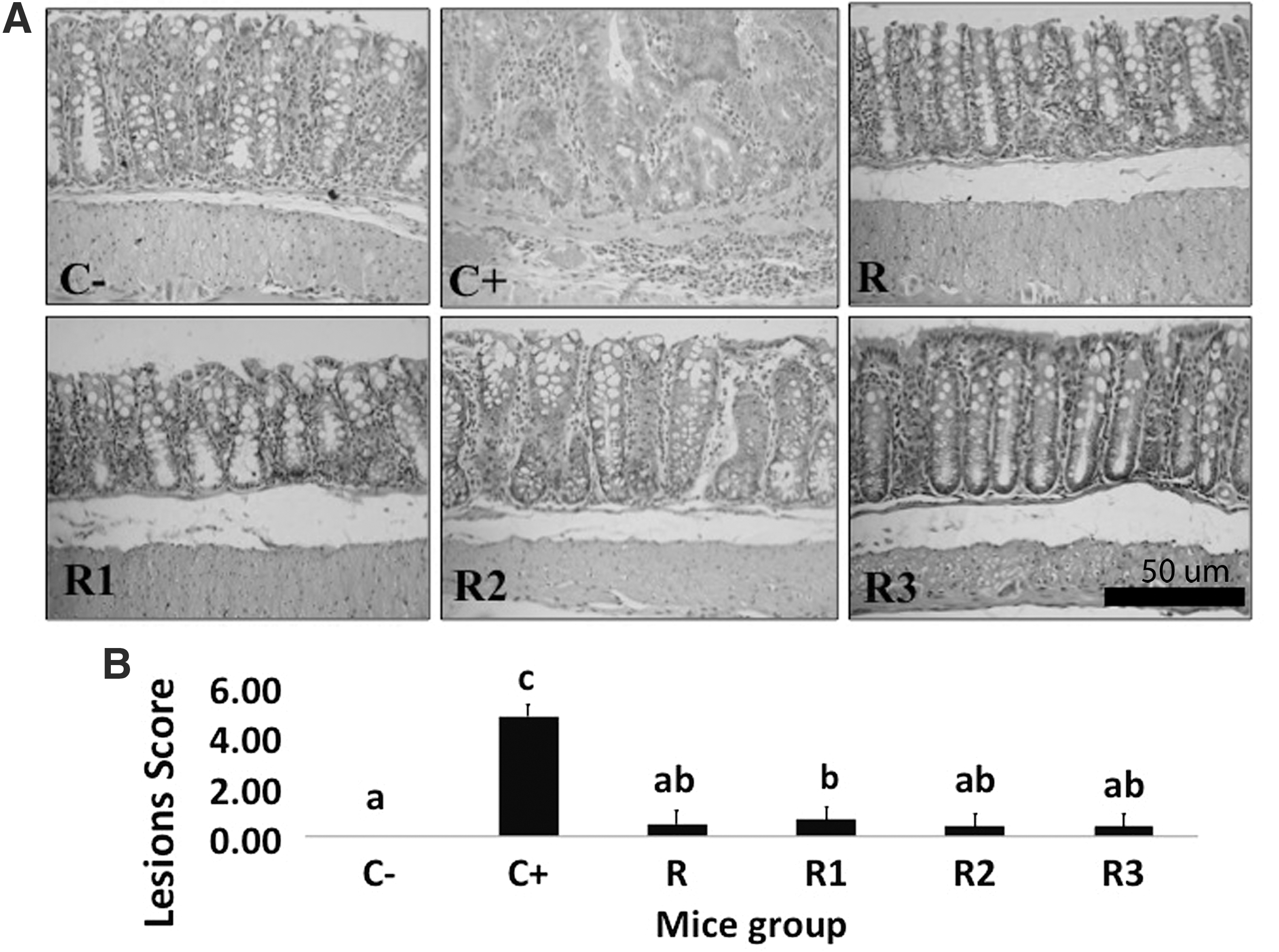
Figure 3B showed the colon macroscopic lesion score, which indicated the carcinogenesis step (severity) of mouse groups' colon. Lesion score of the C− group represented the normal colon. C+ group had the highest lesion score since tumor colon formation was found in this group. Interestingly, all PSE-NPE groups were shown to have significantly lower lesion scores than that of the C+ group. This result indicated that PSE-NPE addition into mouse diets was able to inhibit colon carcinogenesis in AOM-DSS-induced Balb/c mice.
Effects of plant sterol ester-incorporated nonpuffed extrudate supplementation on inflammation and apoptosis in AOM/DSS-induced Balb/c mice
By using IHC staining techniques, COX-2 and cleaved-caspase expressions in mouse colons were observed. Representative figures for COX-2 and cleaved caspase-3,-8,-9 expressions are presented in Figure 4, whereas their histoscores (brown color intensity and distribution) are presented in Figure 5. More detailed data for all groups are presented in Supplementary Figure S3.
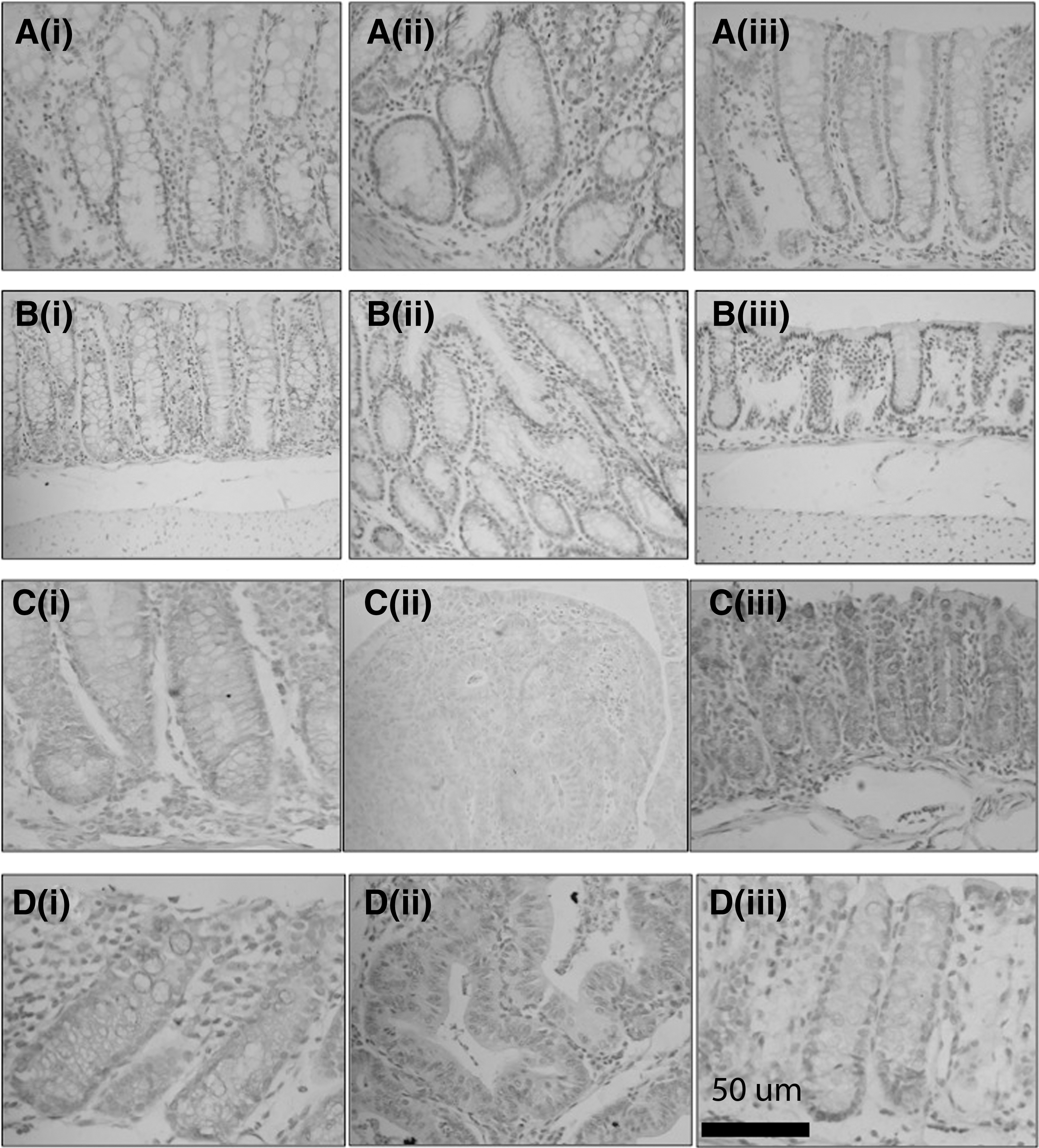
Representative figures of COX-2 and cleaved caspase expression in the colon of mouse groups (IHC staining, 400 × magnification). COX-2 expression of C− group,
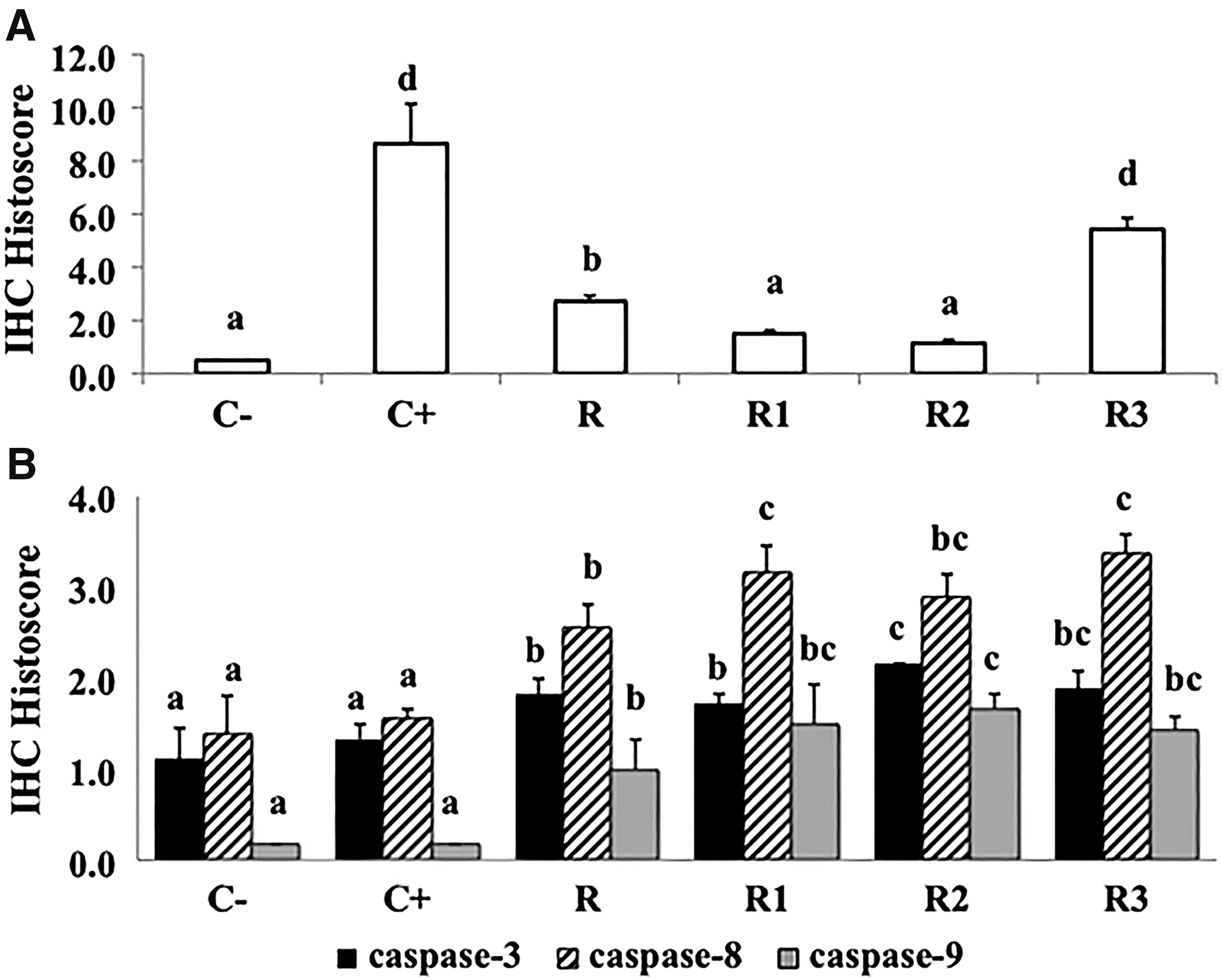
Colon IHC histoscore of COX-2
An intense expression of COX-2 and high COX-2 histoscore were found in C+, indicating that this group had more severe inflammation than the others (Figs. 4Aii and 5A). In contrast, group C− had very low expressions of this enzyme (Figs. 4Ai and 5A). The level of COX-2 expression in PSE-NPE-treated groups was significantly lower as compared with C+ groups. The COX-2 histoscore of R1 and R2 groups was not statistically different from that of C− group, but the score of R and R3 groups was higher than C− group (P < .05) (Fig. 5A). This result indicated that PSE incorporated into extruded product was able to suppress inflammation in AOM/DSS-induced Balb/c mice.
In our in vitro preliminary study (data not shown), plant sterols in the hexane extracts of PSE-NPE at the dose of 1350 mg/L exerted different inhibitory effects on the growth of different human colon adenocarcinoma cells; WiDr (overexpressing COX-2) and HCT-116 cell lines (not expressing COX-2). The inhibitory effects of the sterols against WiDr cells were stronger (>90%) than its inhibitory effect against HCT-116 cells (±20%), which indicated that plant sterols probably specifically inhibit the growth of colon cancer lines that are overexpressing COX-2 (in vitro). Colon cancer cell lines, which do not express COX-2, were reported to be more insensitive to the growth inhibitory and proapoptotic effects of carotenoids. 37
There are no reports yet that indicate that plant sterol esters suppress COX-2 expression in cancer cells. However, a decrease in prostaglandin release in the cultured lipopolysaccharide-activated macrophage cells treated with plant sterols was reported. 38 This decrement was independent from the alteration of COX-2 expression, but it was thought to occur as a result of the COX-2 activity increment. Overexpression of COX-2 in cancer cells was reported to be associated with apoptosis inhibition. 39 Consistent with our previous in vitro study (unpublished data), the present study using Balb/c mice showed that suppression of COX-2 expression was accompanied by an increased expression of caspases.
There are two pathways of caspase activation: the intrinsic pathway, which is closely regulated by a group of Bcl-2 family, cytochrome c, and caspase-9, 40 and extrinsic pathway, which begins when the respective ligands BIND to a death receptor resulting in the formation of death-inducing signaling complex, then activates caspase-8. 41 Both intrinsic and extrinsic pathways converge to caspase-3, which contributes to the typical morphological changes in apoptosis. 42 However, caspase activation requires proteolytic processing of its inactive zymogen into the activated form through the cleavage at an aspartate residue. Detection of cleaved caspase is appropriate to indicate the presence of active caspase. 43
Figure 4B showed cleaved caspase-3expression in representative groups. The colons of all PSE-NPE groups had significantly higher levels of cleaved caspase-3 expression than C− and C+ groups (P < .05) (Fig. 5B). It can be seen from Figure 4C that representative PSE-NPE groups showed the higher intensity of cleaved caspase-8 than that of C− and C+ groups. Cleaved caspase-8 histoscore of R, R1, R2, and R3 groups were higher than that of C− and C+ groups (Fig. 5B). These results were followed by an increase in cleaved caspase-9 expression in the representative groups (Fig. 4D). The colon of PSE-NPE groups had a significantly higher level of cleaved caspase-9 expression than that of C− and C+ groups (P < .05) (Fig. 5B). These results indicated that PSE-NPE chemopreventive activity in AOM/DSS-induced Balb/c mice occurred through apoptosis stimulation through extrinsic and intrinsic pathways.
Several studies have reported the ability of plant sterols to activate both intrinsic and extrinsic apoptosis pathways. β-sitosterol supplementation (16 μM, 3 days) induced the activity of caspase-8 and −9 in human breast cancer cells MDA-MB-231 by 39% and 80%, respectively. 10 Administration of stigmasterol isolated from Navicula incerta was shown to upregulate caspase-8, 9, Bax, and p53 expression levels in human hepatocarcinoma cells HepG2. 44
Different growth inhibitory effects of PSE-NPE hexane extracts in our previous in vitro study (unpublished data) were also suggested, due to the differences in p53 expression among cancer cells. The p53 wild-type expressing cells (HCT-116) were found to be insensitive to the growth inhibitory effects of plant sterols in PSE-NPE hexane extracts than that of p53 mutant-expressing cells (WiDr). Tumor suppressor gene p53 can induce cell cycle progression, DNA repair, and apoptosis through intrinsic and extrinsic pathways. 45 This may indicate that p53 mutations inhibit cell cycle and DNA repairs, thus WiDr cells showed higher inhibition against PSE-NPE hexane extracts. We suggested that PSE-NPE addition into mouse diet could protect p53 gene against mutation arising from carcinogen induction by activating both intrinsic and extrinsic apoptosis pathways.
An increase of caspase activity after β-sitosterol administration was thought to be the result of alterations in the cancer cell membrane structures and functional alterations after β-sitosterol incorporation. 46 Lipid rafts of cell membrane are the regions in which cholesterol is highly concentrated and tend to form characteristic domains with sphingolipids or phospholipids. This an important region for signaling process, including apoptosis pathway. 47 Phospholipids had stronger interactions with cholesterol than plant sterols. 48 Lipid raft structure alteration as a result of plant sterol incorporation might have a beneficial effect on apoptosis signal transduction.
In this study, the expressions of cleaved caspase-3, −8, −9 were not statistically different between C− and C+ groups (Fig. 5B). The higher fat content in mice diet was reportedly correlated with higher serum leptin and decreased expression level of total PARP, PARP-89, PARP-24, Bax, Bcl-2, Bcl-xL, and caspase-3 activity. 49 An increased expression of cleaved caspases in PSE-NPE groups were presumed to be due to the presence of plant sterol ester as a fat component and phytochemical compounds from NPE base ingredients.
PSE addition (0.7%, 1.4%, and 2.1%) into NPE formulation showed higher chemopreventive activity as compared with NPE without PSE addition (Fig. 5), which indicated that PSE retained its colon cancer inhibitory activity after being subjected to high pressure and high temperature during the extrusion process. PSE was reported to have high stability to heat exposure due to steric hindrance obtained from double bond in the B-ring structure. 50 In this study, group R2, which received NPE with 1.4% PSE, showed the optimum inhibitory effect. However, group 3, which received higher levels of PSE addition (2.1% PSE), showed no significant cleaved caspase expression levels when compared with R2 group, although its COX-2 expression was the highest.
The role of phytochemical content of NPE in colon cancer inhibition
Besides plant sterols, NPE developed in this study also contained other active compounds that might contribute to its chemopreventive activity. PSE content of NPE was in the range of 203.84–1617.49 μg/g. NPE also contained dietary fiber (56,000 μg/g), γ-oryzanol (6.80 μg/g), α-tocopherol (2.06 μg/g), flavonoids (206.51 μg/g), and phenolic compounds (223.18 μg/g) (Table 2). Many studies reported the role of those active compounds in preventing colon carcinogenesis. Fermentation of soluble dietary fiber in the colon produces short-chain fatty acids, leading to a decrease in colon pH, which is associated with a reduced incidence of colon cancer. 51,52 γ-Oryzanol, a unique component of rice bran, inhibited tumor growth in CT-26-transplanted Balb/c mice by inducing the activity of natural killer cells and macrophages. 53 α-Tocopherol and γ-tocopherol showed colon cancer inhibition in Fe-induced rat by decreasing the formation of mutagens arising from the free radical oxidation of fecal lipids. 54 Quercetin is one of the most abundant flavonoids present in fruits and vegetables. 55 Quercetin's chemopreventive activity in AOM-treated mice and human colon cancer lines (Caco-2 and HuTu-80) has been reported. 56,57 Gallic acid is a strong phenolic antioxidant. It exerted significant chemopreventive effects on DMH-induced rat and induced apoptosis in HCT-15 colon cancer cells. 58,59 Apart from PSE, these phytochemicals might also contribute to the chemopreventive activity of NPE developed in this study.
Data are mean of three replications. Data are expressed as mean ± SD. Total flavonoid content is expressed as quercetin equivalent (QE). Total phenolic content is expressed as gallic acid equivalent (GAE).
Conclusion
PSE-NPE significantly inhibited colon carcinogenesis in AOM-DSS-induced Balb/c mice. Based on the macroscopic lesion scores and IHC histoscore of COX-2 and cleaved caspase-3,-8,-9 expressions, the optimal dose was 1.4% PSE-NPE. Our results indicated that PSE retained its colon cancer chemopreventive activity after being incorporated into a food system, which was processed using high pressure and temperature. The expressions of cleaved caspase-3, −8, and −9 were increased in PSE-NPE treated mice, whereas COX-2 was decreased. Thus, the PSE-NPE chemopreventive activity is believed to be primarily due to suppressing inflammation and activation of both intrinsic and extrinsic apoptosis pathway.
Footnotes
Acknowledgments
This study was funded by 2013 BASF Newtrition™ Asia Research Grant and 2015 ITSF (Indonesia Toray Science Foundation) Research Grant. The authors would like to acknowledge Abdullah Muzi Marpaung for reading this article carefully.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.