
Abstract
Histone lysine acetylation is thought to play a role in regulating the balance between energy storage and energy expenditure. However, the epigenetic mechanisms by which food phytochemicals influence metabolic processes in the liver have not been thoroughly investigated. In this study, we investigated the effect of an ethanol extract of Capsella bursa-pastoris (ECB) on histone acetyltransferase (HAT) inhibition, and whether it could thereby attenuate lipid accumulation in vitro and in vivo. We observed that ECB inhibits HAT activity as assessed by colorimetric and autoradiography assay systems. ECB also reduced oleic acid (OA)-stimulated histone acetylation at H4K5 and H4K12 and attenuated OA-mediated lipid accumulation in HepG2 cells, in the absence of observable cytotoxicity. We then investigated these effects in vivo. Mice were fed on either a normal diet (ND) or high-fat diet (HFD) in the presence or absence of ECB supplementation. In comparison with the ND controls, the HFD mice exhibited higher body weight, liver fat, adipose tissue size, and total serum cholesterol concentrations, and these effects were significantly attenuated by ECB supplementation. Taken together, these results suggest that ECB protects against the mechanisms responsible for HFD-induced hepatic steatosis, and may involve the targeting of histone H4K acetylation.
Introduction
E
Nonalcoholic fatty liver disease (NAFLD) arises from chronic lipid accumulation because of dysregulated lipid metabolism in the liver and is associated with obesity, dyslipidemia, hypertension, insulin resistance, and diabetes. 2 Epigenetic alterations associated with these chronic metabolic diseases include the abnormal regulation of hepatic lipid metabolism, insulin resistance, mitochondrial damage, oxidative stress, and the release of inflammatory cytokines, all of which have been implicated in the development and progression of NAFLD. 3 It has been postulated that certain bioactive food phytochemicals may influence the prevention and development of NAFLD, because of their effects on the epigenetic modifications responsible.
Capsella bursa-pastoris, colloquially known as Shepherd's purse, is a traditional medicinal plant that has been used for centuries in Korea. Extracts from the plant have been shown to confer a number of effects potentially beneficial for human health, including anti-inflammatory, antioxidative, anticancer, and antidiabetic properties. 4 –8 However, whether extract of Capsella bursa-pastoris (ECB) influences the epigenetic mechanisms responsible for NAFLD has not been previously investigated.
In this study, we confirmed a previous observation that the ethanolic ECB can inhibit HAT and investigated whether that inhibition can attenuate lipid accumulation in vitro and in vivo.
Materials and Methods
Cell culture
HepG2 cells were purchased from the American Type Culture Collection (Mannassas, VA, USA). Cells were cultured in a humidified atmosphere of 5% CO2 at 37°C using high-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum and antibiotics (Welgene, Daegu, Republic of Korea). Cells were incubated with 1% BSA low glucose DMEM (normal diet [ND]), 0.5 mM oleic acid (OA) in 1% bovine serum albumin (BSA) low glucose DMEM (OA), and 0.5 mM OA DMEM supplemented with 50, 100, or 200 μg/mL ECB for 24 h.
Preparation of Capsella bursa-pastoris extract
Capsella bursa-pastoris was purchased from a local market (Republic of Korea) and extracted in a 10-fold volume of 70% ethanol by shaking for 24 h at 25°C, and the precipitate was removed by centrifugation at 8000 g for 30 min. The supernatants were then freeze dried.
Cell toxicity
Cells were seeded in 24-well plates and treated with ECB at 50, 100, 200, or 400 μg/mL. After a 24 h incubation, the cells were treated with 10 μL of water-soluble tetrazolium salt (WST)-1 solution for 3 h. Supernatant (100 μL) was then transferred to a 96-well plate and absorbance was measured using ELISA.
Lipid contents
Cells were seeded in 24-well plates and treated with ECB for 24 h, before lipid content was evaluated using oil red O staining. After washing with 200 μL of phosphate-buffered saline (PBS), cells were fixed with 200 μL of 4% paraformaldehyde for 15 min. The cells were then washed three times with PBS and incubated with 200 μL of 60% isopropanol for 5 min, before staining with 200 μL of 0.1% oil red O staining solution Sigma-Aldrich (St. Louis, MO, USA) for 60 min. After additional washing with water (1 mL), the images were captured under a light microscope. For quantification, lipids were dissolved by incubating cells with isopropanol for 10 min, and absorbance was measured at 510 nm.
Immunoblot assay
After treatment, the cells were lysed with RIPA buffer (Elpis, Daejeon, Republic of Korea) containing protease and phosphatase inhibitors (Roche, Basel, Switzerland). Total protein was measured using the BCA method. The cell lysates containing equal amounts of total protein were then loaded on 10% Bis-Tris gels for electrophoresis before transfer to nitrocellulose membranes. The membranes were blocked in 5% skim milk solution for 1 h and probed with primary antibodies against histone H4K5Ac, histone H4K12Ac, total histone H4 (Cell signaling, Danvers, MA, USA), and β-actin (Enzo life sciences, Inc., Farmingdale, NY, USA) at 4°C overnight. After washing with PBS three times, membranes were incubated with the appropriate horseradish peroxidase-conjugated secondary antibodies for 1 h. After washing with PBS again, the protein bands were detected by chemiluminescence.
Histone extraction
Cells were lysed according to the manufacturer's protocol. Cells were washed with PBS and lysed with prelysis buffer to isolate the nuclear fraction. The nuclear pellet was resuspended in lysis buffer, incubated for 30 min on ice, and centrifuged at 9.425 g for 5 min at 4°C. The supernatant was transferred to a clean tube and Balance-DTT buffer was added. Samples were stored at −70°C until analysis.
HAT assay
HeLa cell nuclear extract was purchased from Biovision Biotechnology (Militas, CA, USA). Nuclear extracts were assessed for HAT activity using a commercially available kit, according to the manufacturer's protocol (Biovision Biotechnology). For in vitro HAT activity assays using autoradiography, HeLa cell nuclear extracts were incubated with HAT assay buffer (50 mM HEPES, pH 8.0; 10% glycerol; 1 mM DTT; 1 mM PMSF; 10 mM sodium butyrate), 1 μL [ 3 H] acetyl-CoA, and 5 μg of biotinylated-H4 peptide (Millipore, MA, USA) with 70% ethanol ECB at the indicated concentrations at 30°C for 1 h. The reactions were stopped by adding 5 × sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer and separated on 15% SDS-PAGE gels followed by autoradiography analysis.
Animal experiment
Male C57BL/6J mice (4 weeks old) were obtained from Chung-Ang Animal Laboratory (Seoul, Republic of Korea) and housed under a 12-h light/12-h dark cycle in a temperature- and humidity-controlled room (24°C ± 1°C at 50% relative humidity). The mice were fed freely on a 10% fat ND, a 60% kcal high-fat diet (HFD), or a 60% HFD plus 2% ECB (HFD +2% ECB) for 12 weeks. Body weight was measured every week. After 12 weeks, the mice were sacrificed. All animal experiments were approved by the Institutional Animal Care and Use Committee of the Korea Food Research Institute.
Quantitation of serum triglyceride and total cholesterol
Serum triglyceride (TG) and total cholesterol (TC) levels were measured with an enzymatic approach using a commercial kit (Asan Pharm, Seoul, Republic of Korea).
Histopathology
Liver tissue and white adipose tissue samples were fixed in 4% buffered formalin and cut into 4-μm-thick sections. The sections were stained with hematoxylin and eosin (H&E) and examined by light microscopy.
Statistical analysis
Data are expressed as mean ± standard deviation. The statistical analyses were conducted using SPSS (Ver. 20; SPSS, Inc., Chicago, IL, USA). Statistical differences between mean values were evaluated by Student's t-test or one-way analysis of variance followed by Bonferroni post hoc test. A value of P < .05 was considered to be statistically significant.
Results
ECB is a specific inhibitor of HATs
During screening of a natural substance library for anti-HAT activity, we observed that 70% ethanol ECB exhibited potent anti-HAT activity. To examine whether ECB could affect HAT enzyme-related epigenetic activity, HAT activity was measured in a cell-free system. As shown in Figure 1A, ECB significantly inhibited HAT activity in a dose-dependent manner. We then examined the inhibitory effect of ECB using radiolabeled H4 tail peptide by SDS-PAGE and autoradiography (Fig. 1B). Consistent with the results from the HAT activity assays, ECB significantly inhibited histone transferase activity. We therefore concluded that ECB is a specific inhibitor of HAT. To ensure that the inhibition of HAT activity was specific, we tested HDAC activity in vitro using the same HAT activity assay. It was observed that ECB had no effect on HDAC activity at the concentrations used (Fig. 1C).
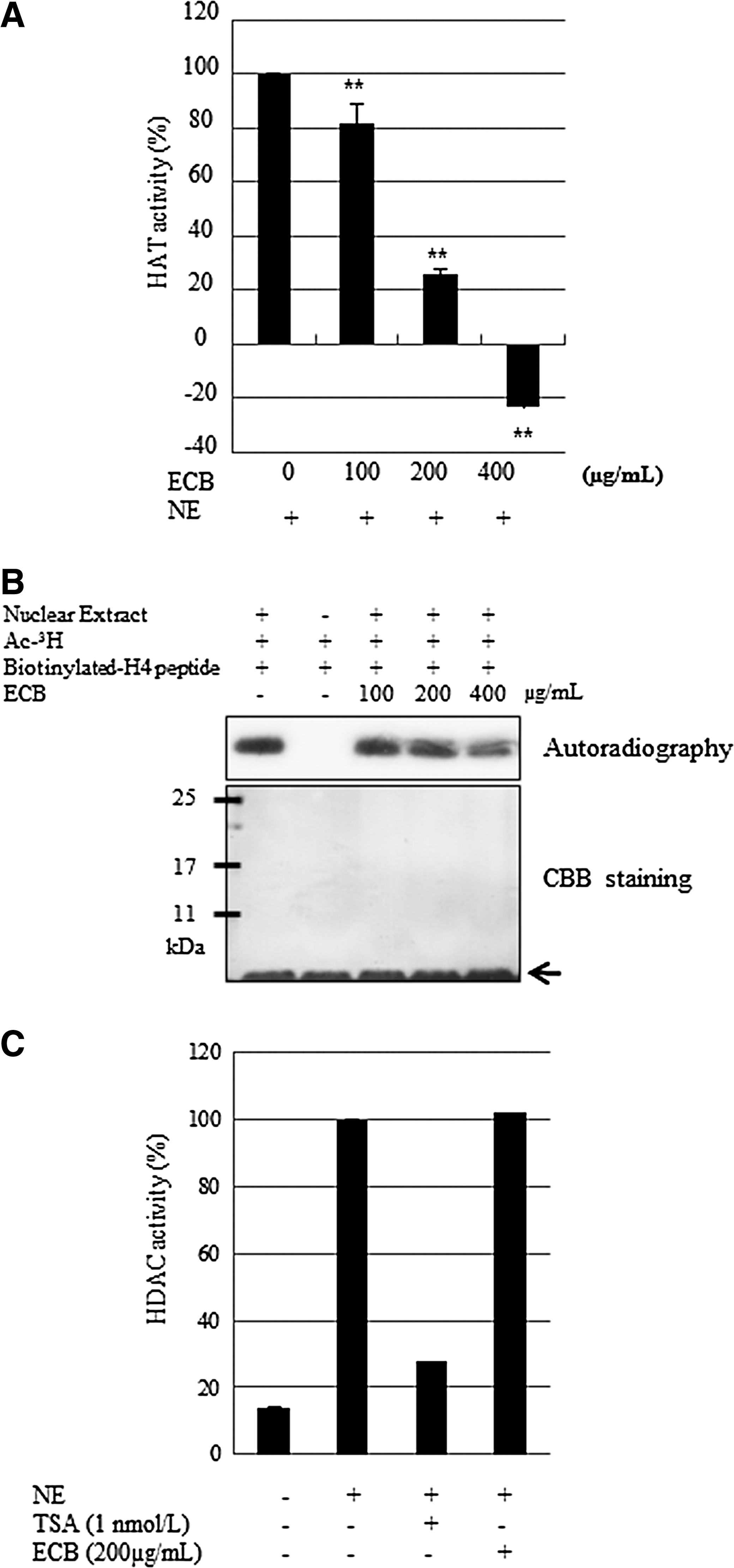
Effect of ethanol extract of Capsella bursa-pastoris (ECB) on histone acetyltransferase activity.
Next, we examined the enzyme specificity of ECB. HAT activity for p300, CREP-binding protein (CBP) and P300/CBP-associated factor (PCAF) was measured in the presence or absence of ECB. ECB was found to be a highly effective inhibitor of p300 acetyltransferase activity. Under similar conditions, ECB also inhibited CBP and PCAF activity although to a lesser extent (Fig. 2A), suggesting that ECB is a general inhibitor of HAT enzyme activity. To know whether the decreased activity of HATs influences histone acetylation, we measured the status of histone acetylation using histone extract. We found that OA treatment only stimulated the acetylation of H4K5 and H4K12, whereas 200 μg/mL ECB reduced OA-mediated hyperacetylation of both H4K5 and H4K12 (Fig. 2B). There were no changes in expression of total histone H4. Altogether, ECB attenuates histone acetylation-induced lipid accumulation through inhibition of HAT activity.
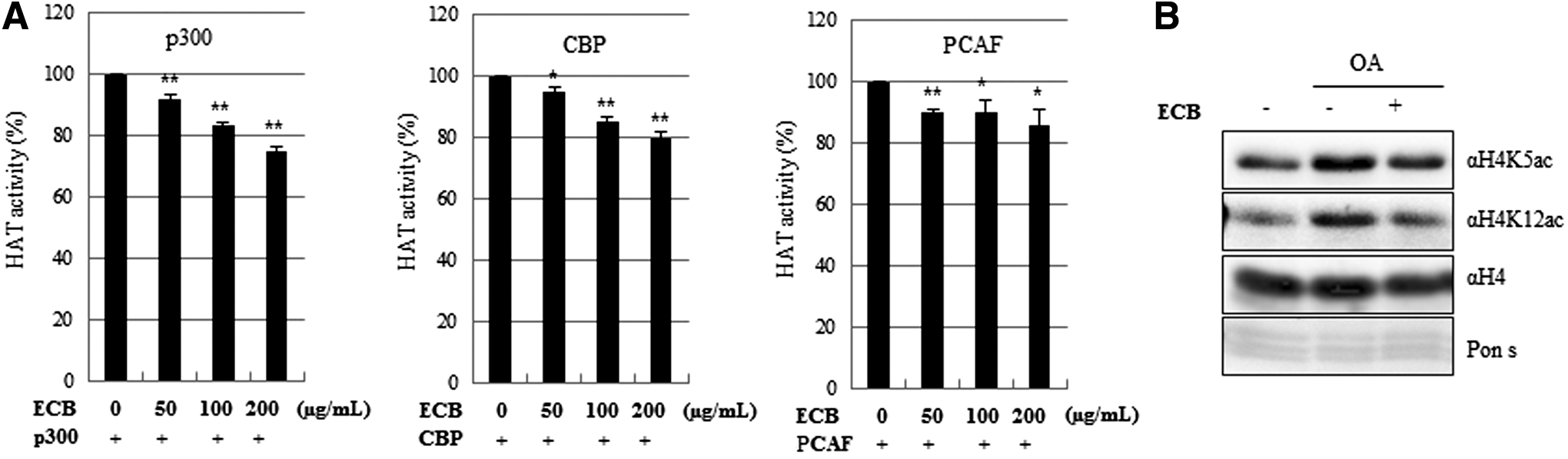
Effect of ECB on specific HAT activity and histone deacetylation.
ECB attenuates OA-Induced H4K5 and H4K12 acetylation and lipid accumulation in HepG2 cells
We next sought to examine the effects of ECB on lipid accumulation in HepG2 cells. HepG2 cells were treated with 0.5 mM OA to induce lipid accumulation, before various concentrations of ECB (100, 200, or 400 μg/mL) were added. It was found that 0.5 mM OA significantly increased lipid accumulation by ∼230%, and cotreatment with OA and ECB attenuated OA-induced lipid accumulation in HepG2 cells (Fig. 3A, 3B). It has previously been established that SREBP1-c and ACLY are regulators of fatty acid synthesis, and we next examined the effect of ECB on transcriptional activity of these genes responsible for lipid metabolism. Treatment with 200 μg/mL ECB was observed to significantly downregulate both SREBP1-c and ACLY gene expressions in OA-induced HepG2 cells (Fig. 3C). These results suggest that ECB inhibits lipid accumulation in cells through the downregulation of lipid metabolic genes by influencing their histone acetylation status.
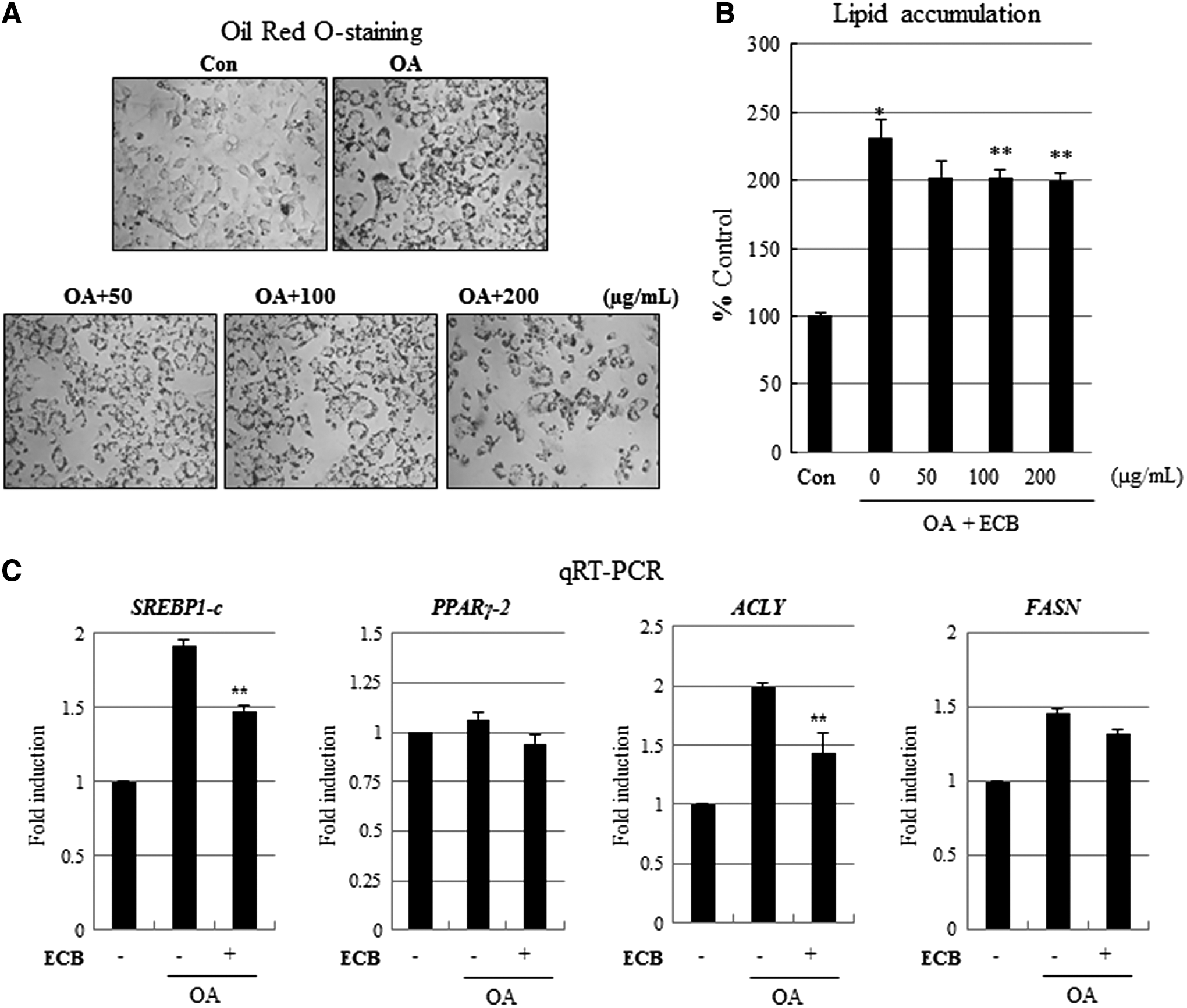
Effect of ECB on histone acetylation in OA-induced hepatic steatosis.
Assessment of ECB cytotoxicity in HepG2 liver cells
We evaluated the cytotoxicity of ECB using WST-1 assay. OA alone or ECB at 50, 100, 200, and 400 μg/mL concentrations did not produce observable cell cytotoxicity (Fig. 4A, B), indicating that ECB attenuates OA-induced lipid accumulation through a specific molecular mechanism rather than general cytotoxicity in HepG2 cells.
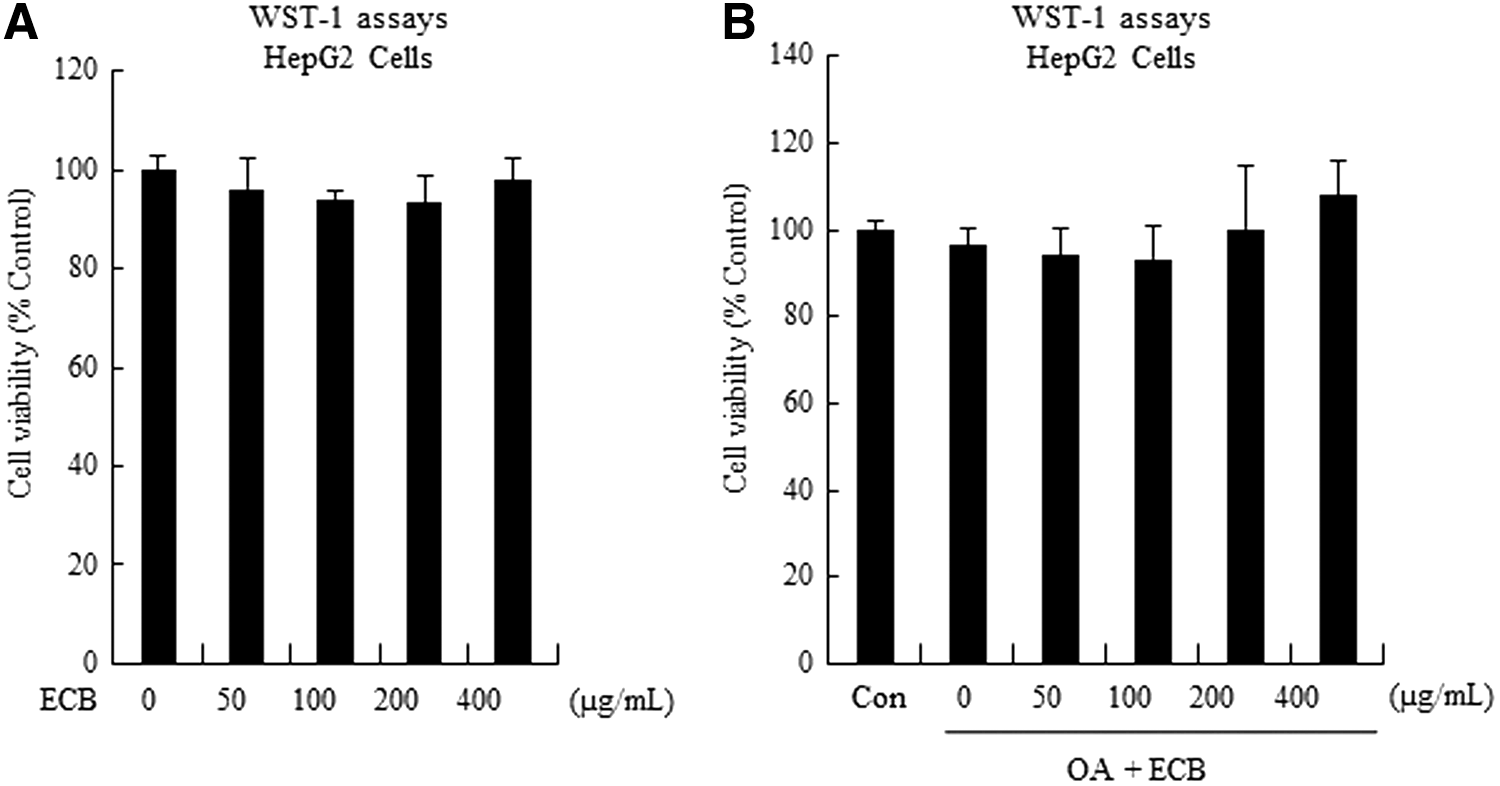
Cytotoxicity test of ECB in HepG2 cells. HepG2 cells were treated with ECB in the presence or absence of OA for 24 h
Dietary supplementation with ECB reduces subcutaneous and body fat mass, as well as total cholesterol levels elevated by a HFD in mice
To further investigate whether ECB can reduce lipid accumulation in vivo, mice were fed on a HFD supplemented with ECB. In comparison with ND controls, the HFD group had significantly increased final body weights (p < 0.001). These gains in body mass were significantly attenuated by ECB treatment (Table 1).
P<.05, ** P<.01, *** P<.001.
ECB, extract of Capsella bursa-pastoris; HFD, high-fat diet; ND, normal diet.
Histological staining revealed that ECB markedly attenuated HFD-induced fat accumulation in the liver (Fig. 5A). We also found that ECB significantly reduced HFD-mediated increases in serum total cholesterol levels, whereas triglyceride levels were slightly reduced in serum after ECB treatment (Fig. 5B). These findings suggest that ECB exhibits preventative effects against the development of diet-induced hepatic steatosis.

Effect of ECB on liver fat accumulation white adipose tissue size, serum TG, and TC. Mice were fed a normal diet (ND), high-fat diet (HFD), and high-fat diet +2% ECB. Liver fat accumulation, white adipose tissue size
Discussion
The structure of chromatin is regulated by the acetylation and deacetylation of histone residues in the histone core around which DNA is coiled. In general, the hyperacetylation of histone tails, which occurs through the inhibition of HATs or induction of HDACs, induces transcriptional activation, whereas hypoacetylation is associated with transcriptional suppression. 9 HAT and HDAC are the two primary enzymes responsible and are involved in the enhancement of acetylation and deacetylation, respectively. The stimulation of histone acetylation by HDACs is associated with the inhibition of adipogenesis in adipocytes. 10 It has been previously shown that the ability of Sprague-Dawley rats fed on a HFD for 4 weeks to restore the hepatocyte population is inversely correlated with the level of hepatic HDAC activity. 11 HDAC1 has been shown to cooperate with the liver-enriched transcription factor C/EBPα for the inhibition of hepatic proliferation in aged mice. 12,13 HAT activity has also been found to influence liver metabolism, because low HAT activity leads to reduced histone acetylation and lower levels of hepatocyte proliferation. 12,13 These findings indicate that a HFD alters the activity of HDACs and HATs, and negatively affects liver function.
HAT and HDAC are plausible targets of dietary phytochemicals that have been shown to affect epigenetic mechanisms. 1 Bioactive compounds from foods, such as genistein, phenyl isothiocyanate, curcumin, resveratrol, indole-3-carbinol, and epigallocatechin-3-gallate (EGCG), have previously been shown to regulate HDAC and HAT activity. 14 These compounds are known to affect the acetylation of histone and nonhistone chromatin proteins, with resultant changes in gene expression. Of particular note, EGCG (present in green tea) has been identified as a potent HAT inhibitor, and also influences chromatin structure by altering histone acetylation, with important implications for human health. 15 In addition, EGCG has been shown to prevent the development of obesity by stimulating fatty acid oxidation in HFD-fed mice, whereas supplementation with the compound reverses liver damage and lowers liver triglyceride levels, plasma cholesterol, and inflammatory cytokines in mice. 16,17 EGCG therefore serves as a prominent example of a food phytochemical exerting beneficial effects relevant for health in the absence of apparent cytotoxicity.
It has been shown that Capsella bursa-pastoris contains several constituents including flavonoids, polypeptides, choline, acetylcholine, histamine, tyramine, fatty acids, sterols, organic acids, amino acids, sulforaphone, trace elements, vitamins, and many other bioactive compounds. 18 In addition, a number of studies have reported that Capsella bursa-pastoris extracts possess antimicrobial, anti-inflammatory, antioxidant, anticancer, and other pharmacological properties. 4 –8 However, Capsella bursa-pastoris-mediated effects on lipid metabolism in the liver have not been investigated in detail. We report for the first time that the ethanol ECB reduces HAT activity dose dependently. ECB appears to mediate HAT enzymes including p300, CBP, and PCAF, and this is associated with reduced OA-stimulated acetylation of H4K5 and H4K12 in vitro. Our in vitro observations were also found to translate into significant metabolic benefits in vivo. Collectively, these results suggests that ECB acts as an HAT inhibitor through the regulation of histone acetylation, which likely contributes to the attenuation of lipid accumulation induced by OA treatment. Further investigation of the specific components responsible for the effects of ECB is therefore warranted, and may lead to the development of novel treatments for the prevention and treatment of NAFLD.
Footnotes
Acknowledgment
This research was supported by grants from the Korea Food Research Institute, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.