
Abstract
We reported previously that supplementation with apple pectin, a dietary fiber, reduced myocardial injury in a rat model of ischemia–reperfusion. In this study, we further investigated an arabinogalactan, one of the constituent polysaccharides of pectin, to determine which domains comprising pectin were responsible for the protection. In a rat model of 30-min ischemia followed by 3-h reperfusion, supplementation with larch arabinogalactan (LAG) over 50 mg/kg/day significantly reduced infarct size. Reverse transcription polymerase chain reaction (RT-PCR), immunohistochemistry, and immunoblot analyses showed that intake of LAG blocked the steps involved in apoptotic cascades through downregulation of gelsolin gene expression at the protein (Gelsolin) level, inhibition of p38 phosphorylation in mitogen-activated protein kinase (MAPK) pathways, decreased bax/bcl-2 ratio at the protein (Bax/Bcl-2) level, which was correlated with the ratio at the mRNA level, inhibition of the conversion of Procaspase protein to Caspase-3 protein, and consequently a decrease in apoptotic cells. In addition, the intake of LAG reduced the hif1-α gene expression at the protein (HIF1-α) level. These findings suggest that arabinogalactan is an active component of pectin for reducing myocardial injury by inhibiting apoptosis in postocclusion steps, possibly indicating that arabinogalactan can be developed as a cardioprotectant to prevent myocardial injury.
Introduction
C
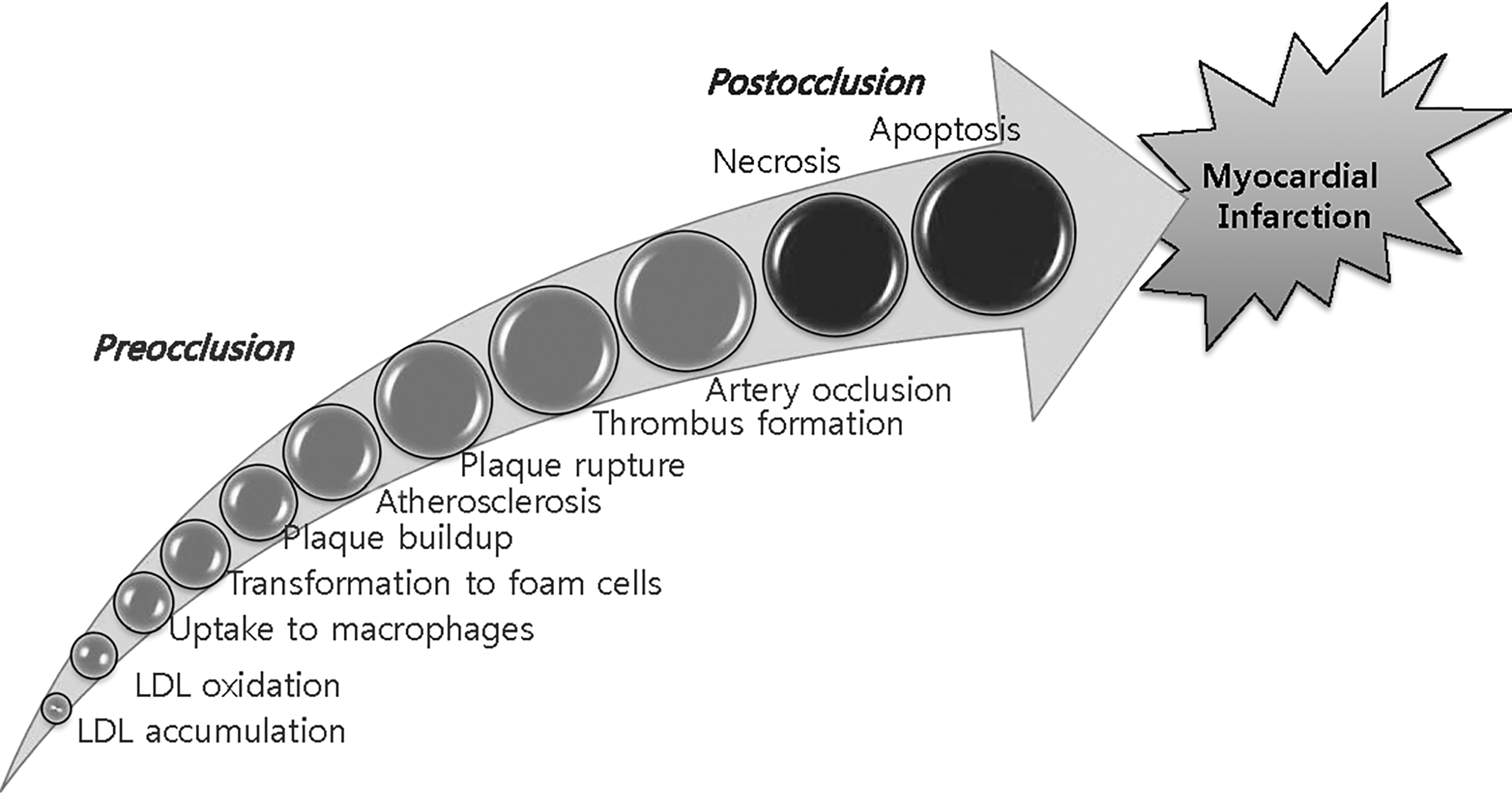
Steps leading to MI. Steps involved in MI can be divided into preocclusion and postocclusion steps. Preocclusion steps include those leading up to the occlusion of coronary arteries. Examples of preocclusion steps are accumulation and oxidation of LDL in the intima, engulfment of oxLDL into macrophages and the subsequent transformation of the macrophages into foam cells, formation of atherosclerotic plaques, rupture of the plaques, and thrombus formation in the lesions. Postocclusion steps include steps, such as artery occlusion and myocardial ischemia, which cause cell death by necrosis and apoptosis. LDL, low-density lipoprotein; MI, myocardial infarction; oxLDL, oxidized LDL.
Pectin consists of several polysaccharide domains, including arabinogalactan.
9,10
Arabinogalactan consists of
Materials and Methods
Diet preparation
Diets containing LAG (Sigma-Aldrich, St. Louis, MO, USA) were prepared as described previously. 8 In brief, to the 950 g of the LAG-free diet purchased from Unifaith, Inc. (Seoul, Korea), we added three varieties of LAG and corn starch combinations: 0.2 g of LAG with 49.8 g of corn starch, 1 g of LAG with 49 g of corn starch, and 2 g of LAG with 48 g of corn starch (Table 1). These afforded 1 kg of 10, 50, and 100 LAG diets, respectively. For the basal diet, 50 g of corn starch was added to 950 g of the LAG-free diet.
LAG-free diet was purchased in a premixed form.
One kilogram of basal diet was prepared by adding 50 g of corn starch to 950 g of LAG-free diet.
Ten, 50, and 100 mg/kg/day LAG diets refer to the corresponding dose of LAG given per kg of rat per day.
Dose of each constituent was calculated, based on data that a 300 g rat consumed 15 g of LAG diet/day.
LAG, larch arabinogalactan.
Animals
Eight-week-old male Sprague Dawley (SD) rats were purchased from Samtaco, Inc. (Osan, Korea). Experiments were carried out according to the guidelines for animal care and the experimental protocols were approved by the Institutional Animal Care and Research Advisory Committee of Catholic University, Daegu, Korea (Approval No. DCIAFCR-151215-20). Animals were housed with food and water available ad libitum under diurnal lighting conditions and in a temperature-controlled environment until the start of the experiment.
LAG and cycloheximide administration
For LAG administration, rats were randomly assigned to sham, control, and LAG-treated groups (10, 50, and 100 mg/kg/day). The rats in each group were acclimatized for 3 days. Then, in the LAG-treated groups, the rats (∼300 g) were fed 15 g/day of 10, 50, and 100 LAG diets for 3 days before occlusion at doses 10, 50, and 100 mg LAG/kg/day, respectively. Once the rats consumed all the LAG diet, more basal diet was provided thereafter. In the sham and control groups, the rats received only the basal diet. Rats were administered cycloheximide (CHX) once through intraperitoneal injection (2 mg/kg) at 1 h before ligation.
MI model
MI was induced by ligating the left anterior descending (LAD) coronary artery in male SD rats (∼300 g) and releasing the ligation afterward as described previously. 3 In brief, after anesthetization with ketamine and xylazine, rats were intubated and ventilated with air. The hearts were then exposed through a left thoracic incision. To induce ischemia, we ligated the LAD of the rats in the control and LAG-treated group for 30 min. Subsequently, the hearts were reperfused for 3 h via release of the ligation. The rats in the sham group underwent the same experimental procedure without the ligation.
Infarct size assessment
The infarct size (IS) was assessed by 2,3,5-triphenyltetrazolium chloride (TTC) staining as described previously. 3,12 In brief, Evans blue dye was infused into the heart following the LAD religation after the ischemia–reperfusion procedure. Next, the heart slices were stained with TTC after euthanization. The area at risk (AAR) was defined as the area that did not display infiltration of Evans blue dye. AAR was further divided into infarct area (IA; the area not stained with TTC) and border zone (BZ; the area in AAR excluding IA). The AAR, IA, BZ, and left ventricular area (LVA) were determined by computerized planimetry using ImageJ software (ImageJ version 1.43r; NIH). The areas were then used to evaluate the IS and risk size (RS), which are defined as the ratio of IA to AAR and AAR to LVA, respectively.
TUNEL staining
Terminal deoxynucleotidyltransferase-mediated dUTP nick-end labeling (TUNEL) staining was performed as described previously. 3,12 In brief, sections prepared from the harvested hearts were incubated with terminal deoxynucleotidyltransferase (TdT) and 5-bromo-2′-deoxyuridine-5′-triphosphate (Br-dUTP). They were also stained with methyl green to identify the nuclei. Micrographs were taken using an Olympus microscope. The nuclei were counted using ImageJ software (ImageJ version 1.43r; NIH) at 400 × magnification. The percentage of apoptotic cells was calculated as the ratio of TUNEL-positive nuclei to methyl green-stained nuclei.
Immunohistochemical staining
Immunohistochemical staining for Caspase-3, Bcl-2, and Bax were performed as described previously. 3,12 In brief, the paraffin sections were cut at 5 μm, deparaffinized, and rehydrated. And then the antigens in the heart sections were retrieved by boiling them in a microwave with 10 mM citric buffer. After endogenous peroxidase blocking step using 3% H2O2, the sections were incubated with blocking solution containing normal goat serum. Then, the sections were incubated with anti-Caspase-3 primary antibody (1:50 dilution; Cell Signaling, Beverly, MA, USA), rabbit polyclonal anti-Bcl-2, and anti-Bax (1:50 dilution; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). Finally, the sections were stained using Vectastain Elite ABC Kit (Vector Laboratories, Burlington, ON, USA), in which 3,3′-diaminobenzidine tetrahydrochloride (DAB; Roche, Mannheim, Germany) was used for color development. The color intensity was measured using ImageJ software. The sections were analyzed at 200 × magnification. The Caspase-3, Bax, and Bcl-2 levels were assessed by calculating the relative color intensity of its image to that of the control group with the latter set as 1.
Western blot analysis
Protein levels of Bax, Bcl-2, Gelsolin, and HIF1-α, and phosphorylation levels of mitogen-activated protein kinase (MAPK) (p38, JNK, and ERK), were assessed using Western blotting techniques as described previously. 11,13 The heart tissues harvested were homogenized in RIPA buffer (Cell Signaling, Beverly, MA, USA), using the Precellys 24 homogenizer (Bertin Technologies, France). The proteins obtained after centrifugation were separated on SDS polyacrylamide gels and transferred electrophoretically to nitrocellulose membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The membranes were incubated overnight with primary antibody. The primary antibodies used were those against Bcl-2 (1:1000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), Bax (1:1000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), p38 MAPK (1:1000; Cell Signaling, Beverly, MA, USA), phospho-p38 MAPK (1:1000; Cell Signaling, Beverly, MA, USA), JNK1/3 (1:1000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), phospho-SAPK/JNK (1:1000; Cell Signaling, Beverly, MA, USA), p44/42 MAPK (Erk1/2) (1:1000; Cell Signaling, Beverly, MA, USA), phospho-p44/42 MAPK (Erk1/2) (1:2000; Cell Signaling, Beverly, MA, USA), Erk1 (1:1000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), Gelsolin (1:1000; Abcam, Cambridge, USA), and HIF1-α (1:500; Novus Biologicals, Littleton, CO, USA). After washing, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (1:2000; Enzo, Farmingdale, NY, USA). Finally, they were developed with ECL substrate solution (GE Healthcare, Little Chalfont, United Kingdom) using ChemiDoc XRS Gel Imager (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The intensities of the protein bands were quantified using ImageJ software.
Caspase-3 activity assay
Caspase-3 activity was measured using a Caspase-3/CPP32 Colorimetric Assay Kit by BioVision (Milpitas, CA, USA). Heart tissue was homogenized in lysis buffer and incubated on ice for 10 min. Following centrifugation, the supernatant was transferred to a fresh tube, and the protein was quantified using the Bradford assay. Caspase-3 activity assay was performed according to manufacturer's instructions. The sample was incubated with 200 μM DEVD-pNA substrate at 37°C for 1 h. And then, the absorbance of pNA was measured at 405 nm using a microplate reader.
Evaluation of mRNA level
After the MI experiment, the heart was harvested and frozen in liquid nitrogen and stored in a deep freezer at −80°C. Total RNA of the heart tissue was extracted with Ambion TRIzol Reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) according to the manufacturer's instructions. The first strand of cDNA was synthesized from the template RNA (1 μg) with a ReverTra Ace-α Kit (Toyobo, Osaka, Japan). And then, synthesis of the second strand was performed using TaKaRa Taq (Takara Bio, Inc., Shiga, Japan). The polymerase chain reaction (PCR) was carried out using different combinations of temperature and reaction time period for each gene using DNA Engine Thermal Cycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The primers used were as follows. 5′-ATCGCTCTGTGGATGACTGAGTAC-3′ and 5′-AGAGACAGCCAGGAGAAATCAAAC-3′ for bcl-2 gene; 5′-CTGCAGAGGATGA-TTGCTGA-3′ and 5′-GATCAGCTCGGGCACTTTAG-3′ for bax gene; 5′-GAAGTTTGA-CCTGGTGCCTG-3′ and 5′-GCTGCACCGTCTTTAGGATG-3′ for gelsolin gene; 5′-CAAGATCAGCCAGCAAGTCCTTCTGATG-3′ and 5′-AGGTTTCTGTAACTGGGTCTG-CTGGAATC-3′ for hif1-α gene; and 5′-AAGTTCAACGGCACAGTCAAGG-3′ and 5′-CATACTCAGCACCAGCATCACC-3′ for glyceraldehydes 3-phosphate dehydrogenase (gapdh) gene.
Statistical analyses
Values are expressed as means ± SEM. Student's t-test was used for single comparison analysis. For multiple comparisons, one-way ANOVA with post hoc Tukey analysis using SPSS software (IBM SPSS Statistics, version 19, Armonk, NY, USA) was used. The statistical significance was considered to be P < .05.
Results
Effect of LAG supplementation on IS
To determine the effective dose range for protecting against ischemia–reperfusion injury, we examined the effect of three different doses of LAG (10, 50, and 100 mg/kg/day) on the reduction of IS by feeding. After being fed with LAG for 3 days, the rats were subjected to ischemia and subsequent reperfusion. Afterward, the hearts were stained with Evans blue dye and TTC (Fig. 2A). AAR and IA represent the area where blood did not circulate during ischemia and the area of cardiomyocyte death, respectively (Fig. 2B). To compensate for the difference in LVA of each rat, RS, calculated as the ratio of AAR to LVA, [RS(AAR/LVA)], was selected for the assessment of AAR (Fig. 2C). The RS(AAR/LVA) of the LAG-treated group (10, 50, and 100 mg/kg/day) was not significantly different from the control group (P > .05), suggesting that the surgical procedures used to ligate the LAD were reproducible for each group (Fig. 2C). To compensate for the difference in the AAR of each rat, IS, calculated as the ratio of IA to AAR, [IS(IA/AAR)], was selected for the assessment of myocardial injury (Fig. 2C). LAG supplementation at 50 and 100 mg/kg/day significantly attenuated IS(IA/AAR) compared with the control group (41.3% ± 5.08%, 39.5% ± 3.27%, respectively, vs. 56.9% ± 2.7%). As a positive control, we chose cycloheximide, a protein synthesis inhibitor, because it improved heart function through apoptosis inhibition in an ex vivo rat ischemia–reperfusion model. 14 Cycloheximide administration through intraperitoneal injection (2 mg/kg) at 1 h before ligation significantly attenuated IS compared with the control group (39.1% ± 3.6% vs. 56.9% ± 2.7%). The findings indicate that LAG intake over 50 mg/kg/day protects against ischemia–reperfusion injury.
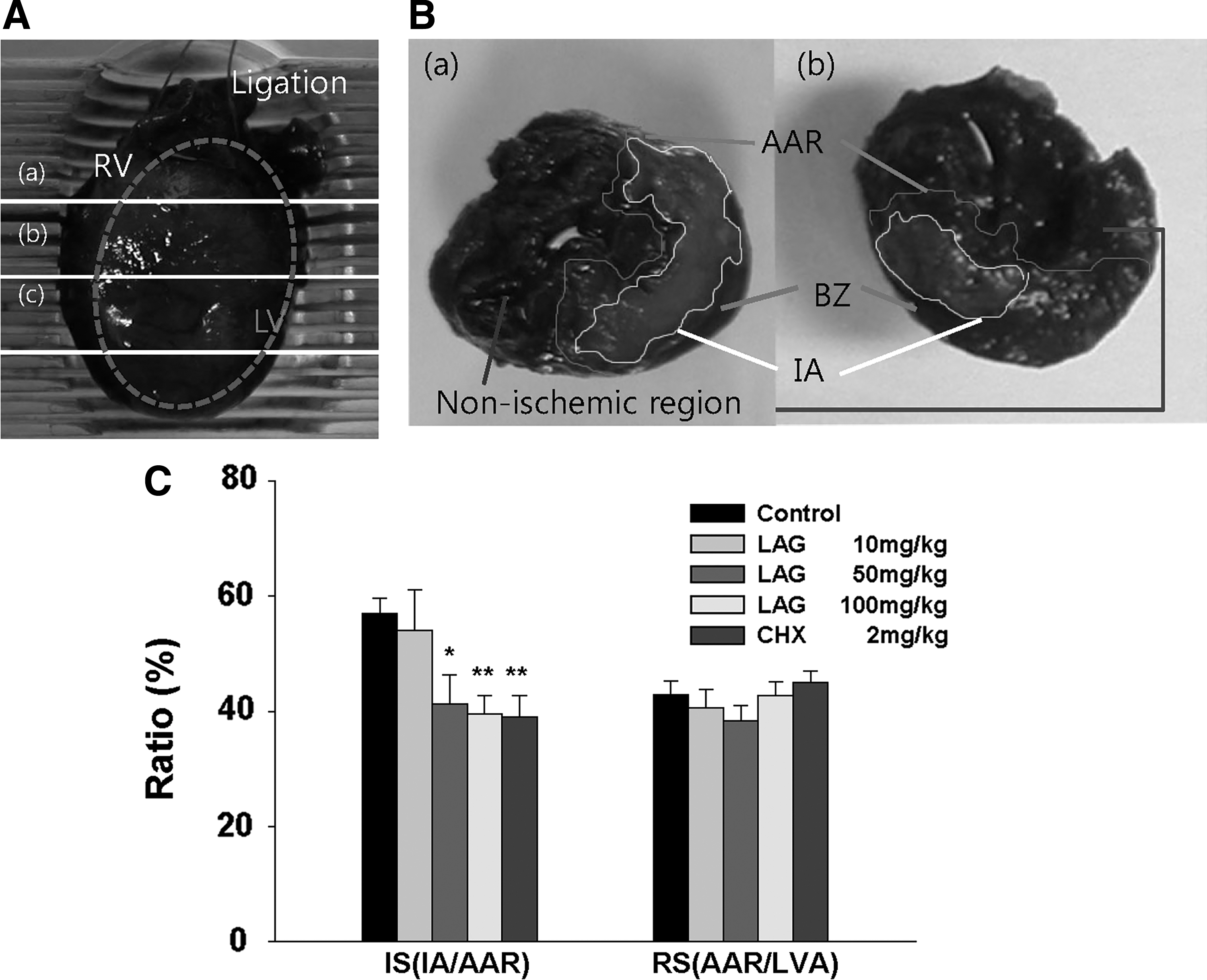
Effect of LAG supplementation on IS in rats.
Effect of LAG supplementation on apoptosis
To test whether LAG supplementation can reduce IS(IA/AAR) through antiapoptotic effects, we chose TUNEL staining because the method has served as a biomarker in detecting apoptotic cells in rat models of ischemia–reperfusion. 15 –17 To evaluate the antiapoptotic effect of LAG, we counted apoptotic cells in AAR separately in IA and BZ, since the efficacy may differ between the two regions. 18 Representative sections from IA and BZ of the LAG-treated group exhibited decreased numbers of apoptotic cells both in IA and BZ compared with the control group (Fig. 3A). To quantify the extent of apoptosis, we assessed the ratio of the number of apoptotic cells to that of total cells (Fig. 3B). In BZ, the ratio was significantly lower in the LAG-treated group at 100 mg/kg/day compared with the control group (7.3% ± 1.7% vs. 17.2% ± 3.2%). In IA, the ratio also tended to be lower for the LAG-treated group at 100 mg/kg/day (15.3% ± 1.2% vs. 24.5% ± 3.3%, P > .05). These results indicate that the protective effect of LAG against ischemia–reperfusion injury in BZ results from antiapoptotic effects of LAG intake.
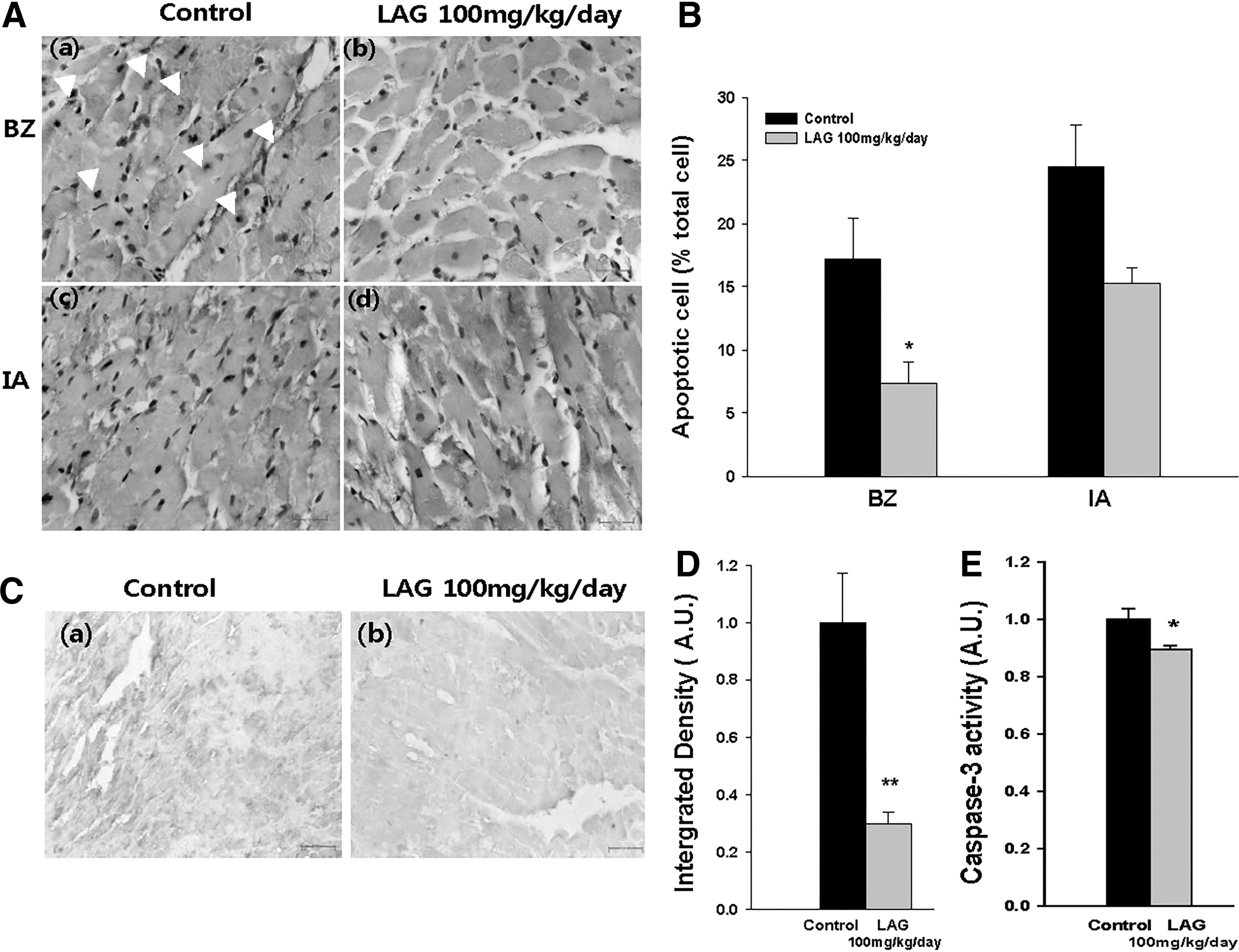
Effect of LAG supplementation on apoptosis and the activation of Procaspase-3 into Caspase-3.
Effect of LAG supplementation on the activation of Procaspase-3 to Caspase-3 (active)
To examine the LAG effect on the apoptotic cascade located further upstream of DNA nicking, we first assessed the caspase-3 protein expression through immunohistochemical staining, in which the color intensity reflects the Caspase-3 level. 19,20 A representative section in BZ of the LAG-treated group (100 mg/kg/day) showed a decreased Caspase-3 level compared with the control group (Fig. 3C). This conclusion was also confirmed by a quantitative assessment (Fig. 3D). The Caspase-3 level was significantly less in the LAG-treated group (100 mg/kg/day) compared with the control group (0.30 ± 0.04 vs. 1.00 ± 0.18). In addition, we also measured Caspase-3 activity (Fig. 3E). The Caspase-3 activity was significantly suppressed in the LAG-treated group (100 mg/kg/day) compared with the control group (0.89 ± 0.02 vs. 1.00 ± 0.04). The results indicate that antiapoptotic effect of LAG intake is mediated by inhibition of Caspase-3 production from Procaspase-3.
Effect of LAG supplementation on bax and bcl-2 genes expression
Expression levels of mRNA (bax and bcl-2) and protein (Bax and Bcl-2) in BZ that are located upstream of caspase-3 were assessed through reverse transcription polymerase chain reaction (RT-PCR), Western blotting, and immunohistochemical staining (Fig. 4A, B). At the mRNA levels, bax/bcl-2 ratio was significantly lower in the LAG-treated group (100 mg/kg/day) compared with the control group (0.53 ± 0.01 vs. 1.00 ± 0.16) (Fig. 4C). At the protein levels, Bax/Bcl-2 ratio, a determinant of the fate of the cells, 21 was also significantly decreased in the LAG-treated group (100 mg/kg/day) compared with the control group, which was assessed through Western blotting (0.39 ± 0.21 vs. 1.00 ± 0.12) and immunohistochemical staining (0.74 ± 0.06 vs. 1.00 ± 0.05) (Fig. 4C).
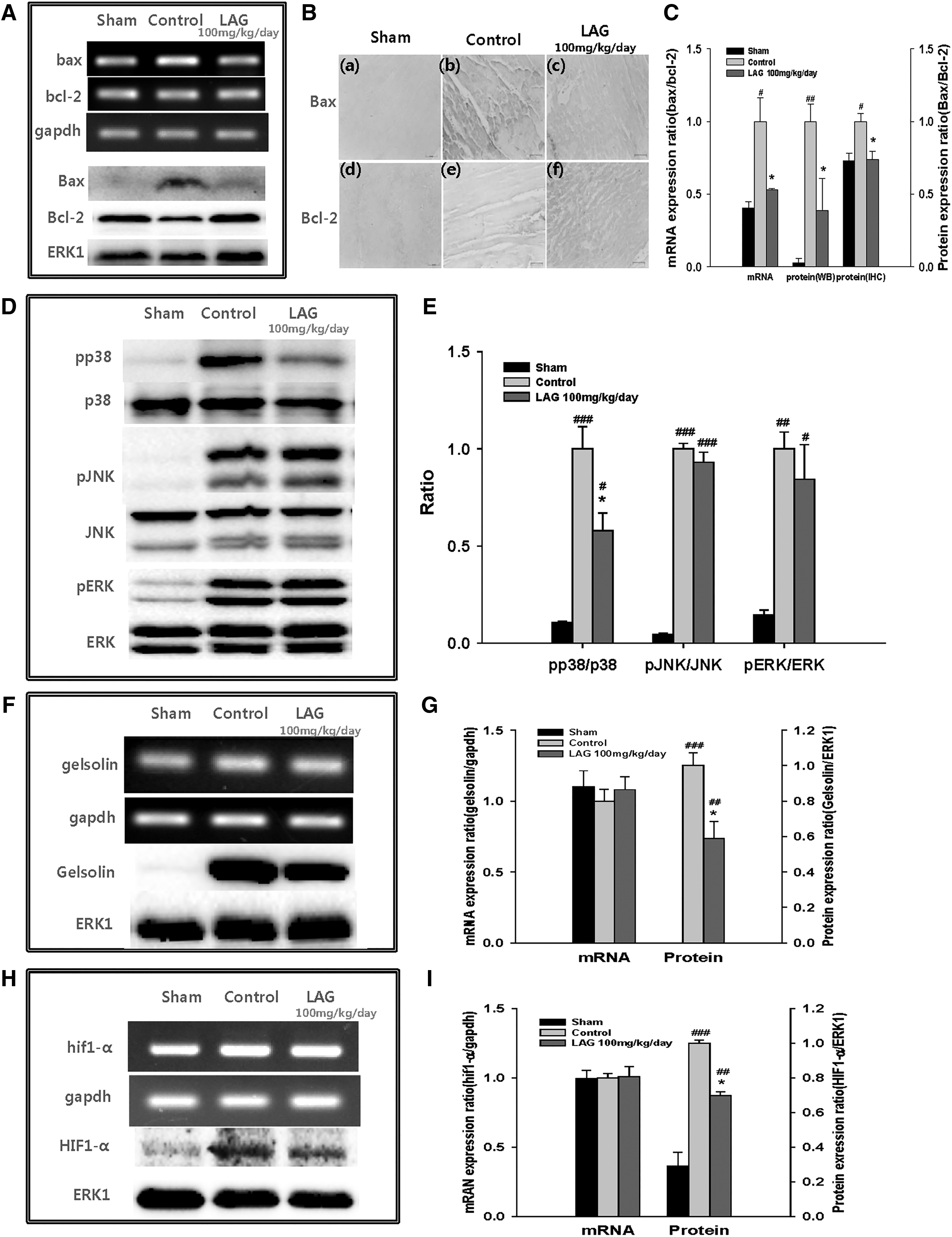
Effect of LAG supplementation on gene expression of bax, bcl-2, gelsolin, and hif1-α and on phosphorylated levels of p38, JNK, and ERK.
The results suggest that protective effects of LAG intake are mediated by steps involving Bcl-2 and Bax in the apoptotic cascade, expression of which was correlated with mRNA levels of the corresponding genes.
Effect of LAG supplementation on MAPK pathways
Phosphorylation levels of mitogen-activated protein (MAP) kinases that modulate levels of bcl-2 and/or bax gene expressions under oxidative stress 22 were evaluated through Western blotting (Fig. 4D). In the quantitative assessment, only pp38/p38 ratio was significantly attenuated in the LAG-treated group (100 mg/kg/day) compared with the control group (0.58 ± 0.09 vs. 1.00 ± 0.11) (Fig. 4E). The results suggest that protective effects of LAG intake are mediated by p38 phosphorylation.
Effect of LAG supplementation on gelsolin gene expression
mRNA (gelsolin) and protein (Gelsolin) expression levels of the gelsolin gene, which modulates pp38/p38 ratio, 23 as well as the levels of Caspase-3, Bax, and Bcl-2 24 were evaluated through RT-PCR and Western blotting 23 (Fig. 4F). In the quantitative assessment, there was no significant difference in the mRNA level between the control and LAG-treated groups (Fig. 4G). However, at the protein level, Gelsolin was significantly downregulated in the LAG-treated group (100 mg/kg/day) compared with the control group (0.59 ± 0.10 vs. 1.00 ± 0.07) (Fig. 4G). The results suggest that protective effects of LAG intake are mediated by the reduction of gelsolin gene expression at the protein level.
Effect of LAG supplementation on hif1-α gene expression
mRNA (hif1-α) and protein (HIF1-α) levels of the hif1-α gene were evaluated through RT-PCR and Western blotting because ablation of hif1-α gene expression attenuates HIF1-α level 24 (Fig. 4H). In the quantitative assessment, there was no significant difference in the mRNA level between the control and LAG-treated groups (Fig. 4I). However, at the protein level, HIF1-α was significantly downregulated in the LAG-treated group (100 mg/kg/day) compared with the control group (0.70 ± 0.02 vs. 1.00 ± 0.02) (Fig. 4I). The results may suggest that protective effects of LAG intake are associated with mitigated level of HIF1-α caused by suppressed gelsolin gene expression.
Discussion
Our previous study revealed that supplementation with apple pectin protects against ischemia–reperfusion injury. 8 As an extension of the previous study, we demonstrated in this study, using LAG as a model constituent, that the arabinogalactan domain was a component in pectin responsible for the observed effect. The mechanism proposed is illustrated in Figure 5. LAG intake decreases gelsolin gene expression at the protein (Gelsolin) level, a calcium-regulated actin that severs and caps proteins in the ischemic area. 24 The downregulated expression of Gelsolin in turn attenuates p38 phosphorylation and decreases Bax/Bcl-2 ratio mediated by bax/bcl-2 ratio at the mRNA level. In addition, the lower level of Gelsolin decreases HIF1-α level, 24 which can promote Bcl-2 expression, 25,26 This combined effect of decreasing Bax/Bcl-2 ratio inhibits activation of Procaspase-3 into Caspase-3, finally leading to attenuation of myocardial cell death by apoptosis, at least in part. Previously, Li et al. 24 demonstrated that the ablation of gelsolin gene expression reduced IS, apoptosis, Bax, and HIF1-α levels, but increased the Bcl-2 level. In addition, Hu et al. 23 demonstrated that gelsolin gene overexpression in cardiomyocytes increases pp38/p38 ratio. Taken together, these findings support our findings in that attenuation of gelsolin gene expression blocks the apoptotic cascade through inhibition of p38 phosphorylation.
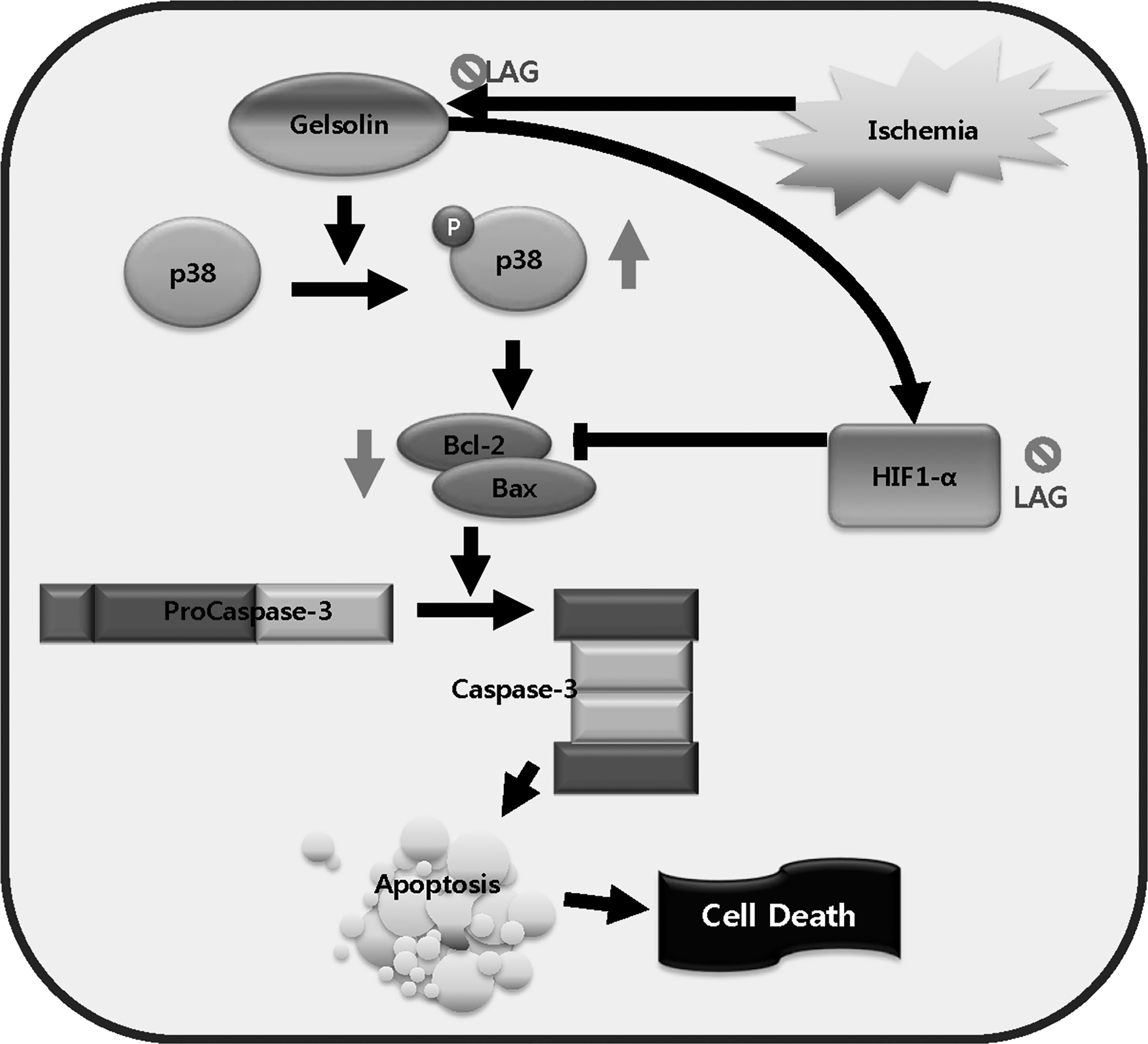
An illustration of the gelsolin gene pathway for blocking apoptotic cascades. LAG intake blocks the apoptotic cascade through downregulation of gelsolin gene expression at the protein (Gelsolin) level. The blockage in turn inhibits p38 phosphorylation, decreases Bax/Bcl-2 ratio, inhibits Procaspase-3 activation, and ultimately attenuates apoptosis. The blockage also reduces HIF1-α level and increases Bcl-2 level, which also contributes to apoptosis inhibition.
Traditionally, it has been thought that intake of dietary fiber, including pectin, reduces the risk of CHD, 27 –29 by blocking preocclusion steps through improving serum lipid levels and blood glucose concentrations. 28,–30 In this study, we demonstrated, for the first time to our knowledge, that LAG intake prevents MI by blocking postocclusion steps. Interestingly, however, LAG supplementation did not affect serum lipids and glucose levels in human clinical trials, 31,32 suggesting that LAG intake may not block preocclusion steps. Overall, the findings suggest that arabinogalactan is an active component of pectin in blocking postocclusion steps.
One plausible mechanism, by which LAG intake might protect against myocardial injury, is through the short-chain fatty acids (SCFA) generated by fermentation of LAG in the large intestine. Indeed, butyrate and propionate can be generated from arabinogalactan by microorganisms inhabiting in the intestine. 33,34 In our previous study, we showed that intraperitoneal administration of SCFA, butyrate and propionate, in particular, protected against myocardial injury in the same rat MI model used in this study. 12 Therefore, butyrate and propionate generated in the large intestine and absorbed into the body might be responsible for reducing myocardial injury.
So far, LAG has been developed as a digestive aid to increase beneficial anaerobes and SCFA production, 35 and immune enhancer to prevent infection. 35,36 In this study, we showed a possibility that LAG can also be developed as a cardioprotectant to prevent MI.
Footnotes
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (grant number: NRF-2016R1A6A3A01010013).
Author Disclosure Statement
No competing financial interests exist.