
Abstract
Aralia elata (Miq) Seem (AES) is a medicinal plant used in traditional Chinese and Korean medicine for the treatment of several diseases, including diabetes. This study aimed to investigate the neuroprotective effect of AES extract against high glucose-induced retinal injury in diabetic mice. AES extract (20 and 100 mg/kg body weight) was orally administered to control mice or mice with streptozotocin-induced diabetes. Protein levels of O-linked β-N-acetylglucosamine (O-GlcNAc) transferase (OGT), carbohydrate-responsive element-binding protein (ChREBP), sterol regulatory element-binding protein (SREBP)-1, thioredoxin-interacting protein (TXNIP), fatty acid synthase (FAS), and acetyl CoA carboxylase (ACC) were analyzed by western blotting. Colocalization of terminal deoxynucleotide transferase-mediated dUTP nicked-end labeling (TUNEL)-positive ganglion cells and OGT, ChREBP, or TXNIP were monitored using double immunofluorescence analysis. Interaction between ChREBP and OGT was assessed using coimmunoprecipitation analysis. AES extract protected the retinas from neuronal injury and decreased levels of OGT, ChREBP, TXNIP, SREBP-1, FAS, and ACC in the diabetic retinas. AES extract reduced colocalization of TUNEL-positive ganglion cells and OGT, ChREBP, or TXNIP in the diabetic retinas. Coimmunoprecipitation analysis indicated that AES extract reduced interaction between ChREBP and OGT and attenuated ganglion cell death in diabetic retinas. Moreover, the ChREBP that colocalized with OGT or the TUNEL signal was significantly decreased in diabetic mice treated with AES extract. These findings show that AES extract can alleviate OGT-, ChREBP-, TXNIP-, or SREBP-1-related retinal injury in diabetic retinopathy.
Introduction
D
ChREBP plays a role in various processes, ranging from glucolipotoxicity to apoptosis or proliferation in specific cell types. 18 ChREBP activity is regulated through multiple posttranslational modifications. 19 Hyperglycemia-induced O-linked β-N-acetylglucosamine (O-GlcNAc) modification of proteins increases glucotoxicity. 20 Of note, O-GlcNAc modification increases ChREBP protein levels and transcriptional activity in liver. 21 O-GlcNAc transferase (OGT) facilitates the interaction and O-GlcNAc modification of key insulin signaling regulators. 22 Moreover, thioredoxin-interacting protein (TXNIP), which is transcriptionally regulated by ChREBP, 23 is a crucial mediator of glucotoxicity-induced β-cell apoptosis. 24 The mechanism through which glucose activates ChREBP is complex, 18 and its role in DR has not yet been elucidated.
SREBP, a master regulator of fatty acid synthase (FAS), controls lipogenesis 25 and is involved in type 2 diabetes. 26 SREBPs, especially SREBP-1c, have a critical role in the pathogenesis of the metabolic syndrome, and therefore identification of compounds that inhibit SREBPs may be useful in the treatment of metabolic syndrome or to ameliorate its inherent complications. 27 In this study, we assessed whether AES affects the adverse changes in retinal neuronal death of diabetic mice, in part, driven by posttranslational modification of ChREBP and SREBP, with the aim of facilitating the development of novel agents for protecting against retinal damage in DR.
Materials and Methods
Sample preparation for high performance liquid chromatography
AES freeze-dried powder prepared as described previously 6 was obtained from Medvill Co., Ltd. (Seoul, Korea). The reference standard (quercetin) and AES (1 g) was dissolved in CH3CN (50 mL) in the concentration of 1 mg/mL and stored at 4°C. The filtrates were concentrated under vacuum and filtered with a 0.45 μm filter before use.
High performance liquid chromatography analysis
Chromatographic separation was performed on Agilent 1200 high performance liquid chromatography (HPLC) system equipped with YMC-C18 column (4.6 × 250 mm, 5-mm) (Agilent Technologies, San Jose, CA, USA). The column temperature was maintained at 35°C, and the flow rate was 1 mL/min. The detector was UV 260 nm. One of the peaks detected by HPLC analysis was identified as the quercetin (Fig. 1A, B).

HPLC and MS chromatograms of CH3CN extract of AES
Mass spectrometry
Identification of peaks was performed using a Synapt G2 quadrupole time-of-flight mass spectrometer (Waters), which was equipped with an electrospray ionization (ESI). The instrument was operated using ESI source in positive and negative mode simultaneously. The source voltage was set to 3100 and −2500 V for ESI+ and ESI−, respectively. The cone voltage, desolvation gas (nitrogen) flow rate, and desolvation temperature were 35 V, 800 L/h, and 350°C, respectively. Mass data were obtained in positive ion mode by a full scan of m/z 50–1200. Data acquisition and processing were carried out using a MassLynx software workstation. One of the abundant peaks shown by HPLC analysis was identified as quercetin (Fig. 1C, D), which is likely to be one of the bioactive compounds, which may protect the retinas from neuronal injury.
Animals
Diabetes was induced in male C57BL/6 mice (KOATEC, Pyeongtaek, Korea), as previously described. 28 All mice were treated in accordance with the guidelines of the Institutional Animal Care and Use Committee of Gyeongsang National University. Control mice were injected with phosphate-buffered saline (PBS). All mice were sacrificed at 2 months after the final injection of 2-deoxy-2-(3-(methyl-3-nitrosoureido)-D-glucopyranose (STZ) or saline. Blood was obtained by tail puncture, and induction of diabetes was verified weekly after STZ injection by evaluating blood glucose concentrations using a Precision glucometer (Abbott, Alameda, CA, USA). Mice with a blood glucose concentration ≥250 mg/dL were considered diabetic.
Cell culture
The ARPE-19 human retinal pigment epithelial (RPE) cell line was obtained from American Type Culture Collection (Manassas, VA, USA) and grown on cell culture plates at 37°C in Dulbecco's modified Eagle medium with 10% fetal bovine serum (Invitrogen, Carlsbad, CA, USA), 100 μg/mL streptomycin, and 100 units/mL penicillin (Invitrogen). Cells were grown in low-glucose (5 mM) or high-glucose (25 mM) medium.
Administration of AES
AES freeze-dried powder was orally administered, as described previously. 16 Mice were treated once a day for 8 weeks after the final STZ or saline injections. Mice were divided into four groups; (1) saline-treated control group, (2) saline-treated diabetic group, (3) AES (20 mg/kg)-treated diabetic group, and (4) AES (100 mg/kg)-treated diabetic group. Blood glucose levels and body weights were measured weekly.
Antibodies
The following antibodies were used: OGT (sc-74546, Santa Cruz Biotechnology; ab96718, Abcam, Cambridge, United Kingdom), ChREBP (NB400-135; Novus Biologicals, Littleton, CO, USA), TXNIP (sc-166234; Santa Cruz Biotechnology, Santa Cruz, CA, USA), SREBP-1 (sc-8984; Santa Cruz Biotechnology), FAS (#3180; Cell Signaling Technology, Danvers, MA, USA), acetyl CoA carboxylase (ACC) (#3662; Cell Signaling Technology), γ-Syn (ab55424; Abcam), β-actin (A5441; Sigma, St. Louis, MO, USA), secondary horseradish-peroxidase-conjugated goat anti-mouse IgG (#31430; Pierce Biotechnology, Rockford, IL, USA), and goat anti-rabbit IgG (#31460; Pierce Biotechnology).
Western blotting analysis
Protein extraction and western blotting were performed as described previously. 28
Immunoprecipitation
Immunoprecipitation was performed as described previously. 29
Immunohistochemistry analysis
Immunohistochemistry was performed on frozen retinal sections (5-μm thick), as described previously. 30 In brief, after blocking with serum, the sections were incubated with a primary antibody against OGT, TXNIP, ChREBP, or SHREBP followed by an Alexa 488- or −594-conjugated secondary IgG (Molecular Probes, Carlsbad, CA, USA). The sections were mounted with mounting medium that contained DAPI (ProLong Gold Antifade Reagent; Invitrogen). All retinal images were captured at a distance of ∼0.8–1.0 mm from the optic nerve head using a JP IX2-DSU disk-scanning confocal microscope (Olympus, Hamburg, Germany).
Immunofluorescence analysis
Immunofluorescence analysis was performed as described previously. 29 In brief, for double staining of OGT and ChREBP or SREBP, we prepared 5-μm-thick retinal sections. The slices were incubated in blocking solution (2% normal donkey serum, 0.5% Triton X-100, 0.05% sodium azide in 0.05 M PBS, pH 7.4) for 1 h, followed by incubation at 4°C overnight with mixed primary antibodies (anti-OGT and ChREBP or SREBP antibodies). After several washes in 0.1 M PBS, fluorescent secondary antibodies (goat anti-Mouse IgG conjugated to Alexa Fluor 488 [A-11029; Thermo Scientific, Waltham, MA, USA] or donkey anti-Rabbit IgG conjugated to Alexa Fluor 594 [A-21207; Thermo Scientific]) were applied, and sections were mounted with mounting medium, including DAPI (Invitrogen). For double staining of ChREBP, SREBP, or OGT along with TUNEL, ChREBP, SREBP, or OGT was labeled with Alexa 488, and immunofluorescent staining for ChREBP, SREBP, or OGT was followed by TUNEL staining. Digital images were captured using a fluorescence microscope (BX51-DSU; Olympus, Tokyo, Japan), and merged to reveal colocalization of proteins in the retinas.
Statistical analyses
Quantitative analyses were performed using Image J image analysis software (Molecular Devices, Sunnyvale, CA, USA) and GraphPad Prism 5 (GraphPad Software, San Diego, CA, USA). Data are representative of three independent experiments and presented as the mean ± standard error of the mean. Data were analyzed using one-way ANOVA followed by Bonferroni's post hoc analysis or t-test to compare groups. Results were considered statistically significant when P was less than .05.
Results
AES reduces levels of OGT in the diabetic retinas
OGT facilitates O-GlcNAc modification of ChREBP, and O-GlcNAcylation increases ChREBP, 21 which plays a potential role in glucolipotoxicity and apoptosis. 18 Therefore, we assessed whether AES affects levels of OGT in the diabetic retinas. We found that levels of OGT were significantly elevated in the diabetic retinas compared with that in controls (P < .005; Fig. 2A). However, AES treatment significantly lowered OGT levels in the diabetic retinas, compared with those not treated with the low or high doses of AES (20 or 100 mg/kg, respectively; P < .005; Fig. 2A), consistent with the data in the immunofluorescence analysis (Fig. 2B). We also found that OGT is present mostly in the nuclear region in the ganglion cell layer (GCL) of the diabetic retinas (Fig. 2B).

AES decreases ChREBP and TXNIP levels and inhibits OGT-ChREBP interaction in the diabetic retinas
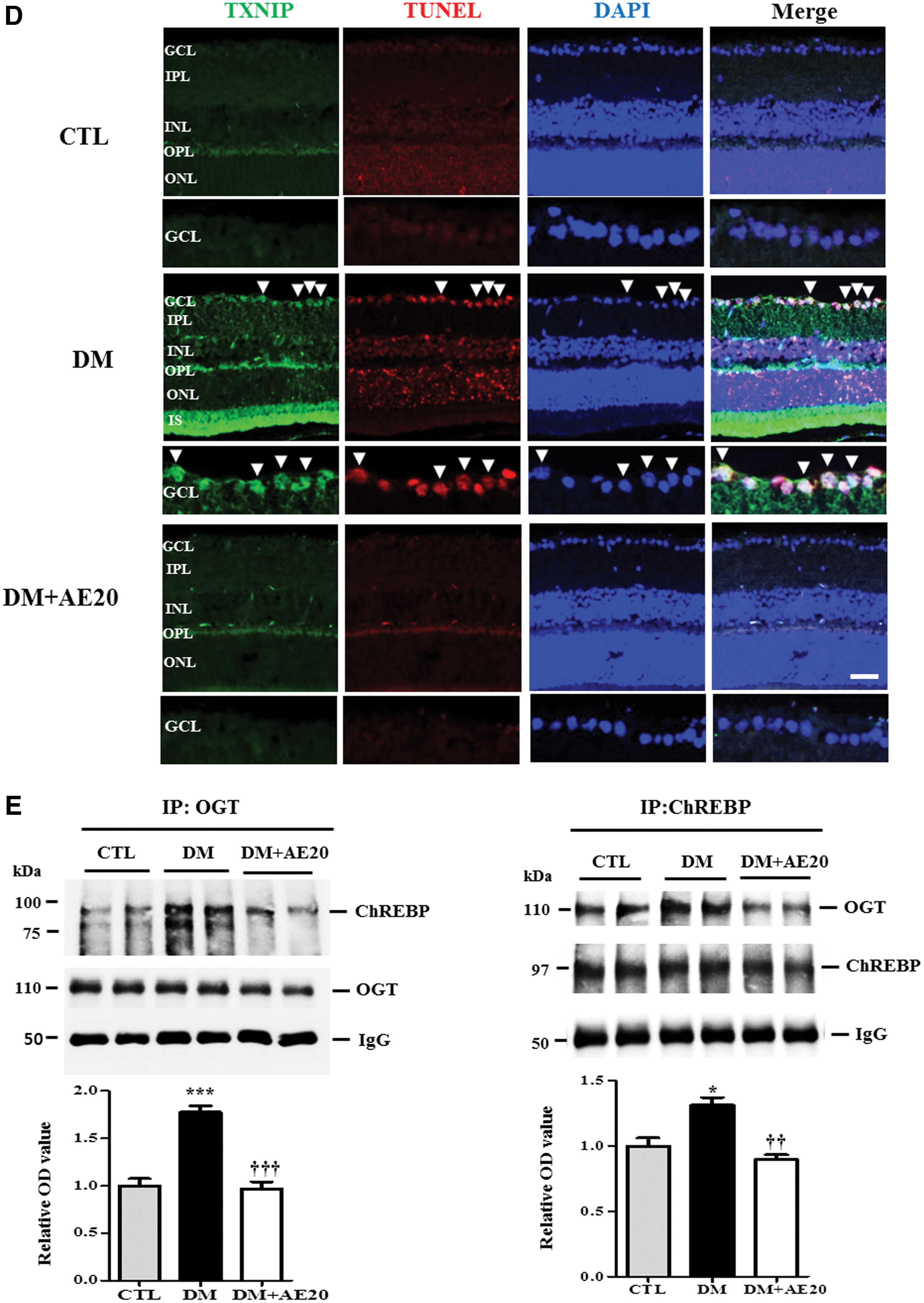
ChREBP promotes glucolipotoxicity as well as apoptosis and affects insulin sensitivity. 18 Therefore, we tested whether AES affects ChREBP activation in the diabetic retinas. Western blotting analysis showed that levels of ChREBP were significantly increased in the diabetic retinas compared with that in controls (P < .05; Fig. 3A). Importantly, AES was able to reverse this increase in ChREBP in the diabetic retinas (P < .05; Fig. 3A). In addition, to evaluate the effect of glucose on ChREBP, we monitored ChREBP levels in RPE cells exposed to low or high glucose. Our data show that ChREBP levels were significantly increased in RPE cells exposed to high glucose, compared with the levels in cells exposed to low glucose (P < .05; Fig. 3B). TXNIP is induced by diabetes and transcriptionally regulated by ChREBP, 23 but the potential link between proapoptotic TXNIP and retinal cell death remains elusive. To address the role of TXNIP in retinal cell death in our model, we assessed whether AES affects retinal TXNIP expression levels in the diabetic retinas. Indeed, we found that TXNIP levels were significantly increased in the diabetic retinas, compared with that in the controls; however, AES treatment greatly reversed the diabetes-mediated increase in TXNIP (P < .005 or P < .001, respectively; Fig. 3C). Notably, immunofluorescence analysis showed significant colocalization of TXNIP and the TUNEL signal in the GCL of the diabetic retinas (Fig. 3D), whereas AES treatment attenuated this colocalization (Fig. 3D). Furthermore, we assessed the interaction between OGT and ChREBP, in part, because O-GlcNAcylation activates ChREBP under high glucose conditions, and determined whether ChREBP is O-GlcNAc modified in diabetic retinas. Immunoprecipitation assays showed that the interaction between OGT and ChREBP was significantly increased in the diabetic retinas, compared with the interaction levels in controls (P < .001 or P < .05; Fig. 3E). Strikingly, AES treatment mostly reversed this change in the diabetic retinas (P < .001 or P < .005; Fig. 3E), suggesting that AES lowered the levels of O-GlcNAc-modified ChREBP in the diabetic retinas. Consistent with these data, we found that ChREBP and OGT were mostly colocalized in the GCL of the diabetic retinas (Fig. 3F). However, AES treatment decreased the extent of colocalization, compared with that in diabetic retinas of mice not treated with AES (Fig. 3F). In addition, immunofluorescence analysis showed that ChREBP, OGT, and the TUNEL signal were colocalized in the GCL of the diabetic retinas, but AES treatment greatly reduced the extent of colocalization (Fig. 3G), suggesting that AES protects against GCL death, at least partially through modulation of ChREBP and/or OGT.

Representative Western blot and quantification of ChREBP
AES suppresses SREBP-1 and decreases FAS and ACC in the diabetic retina
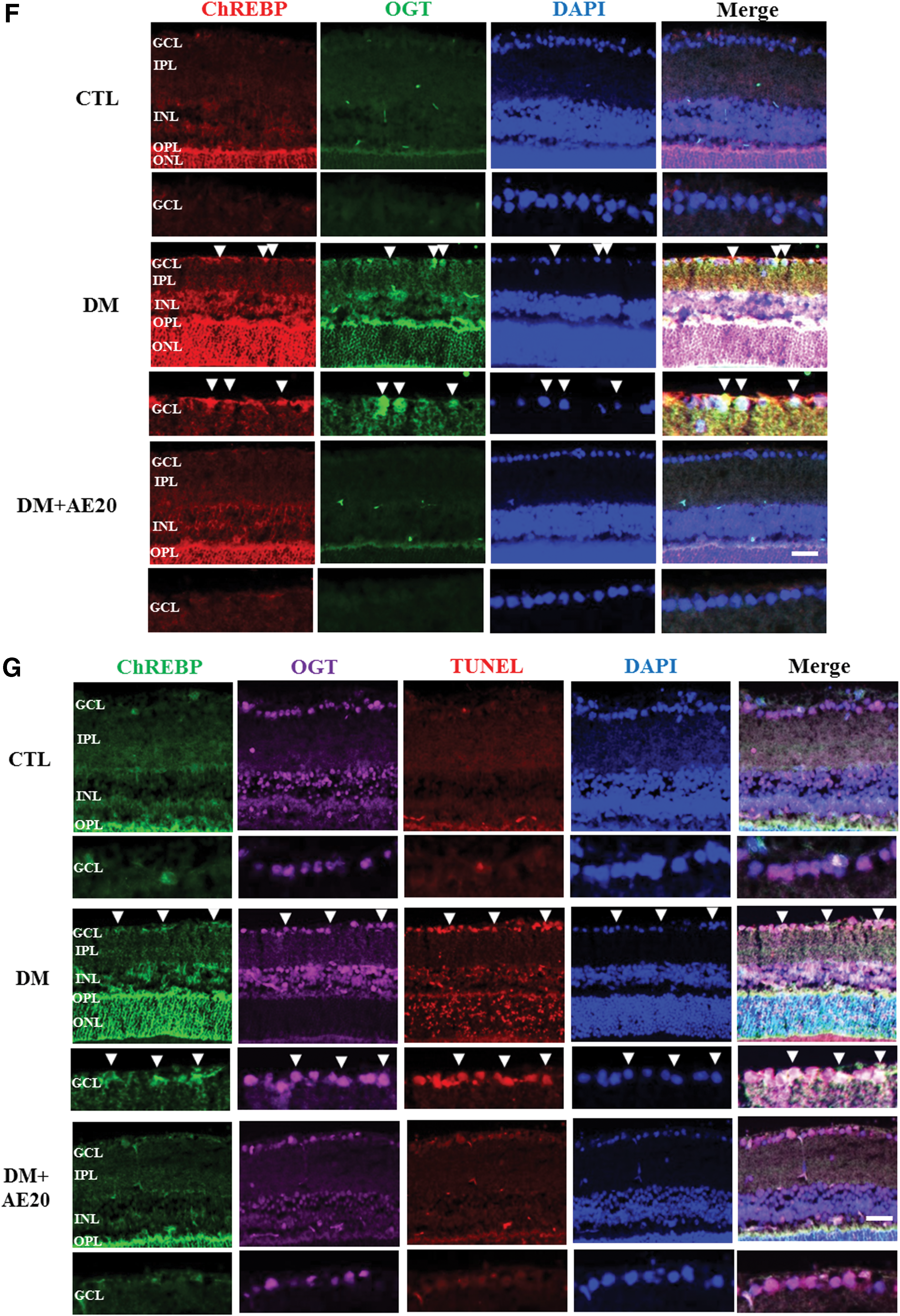
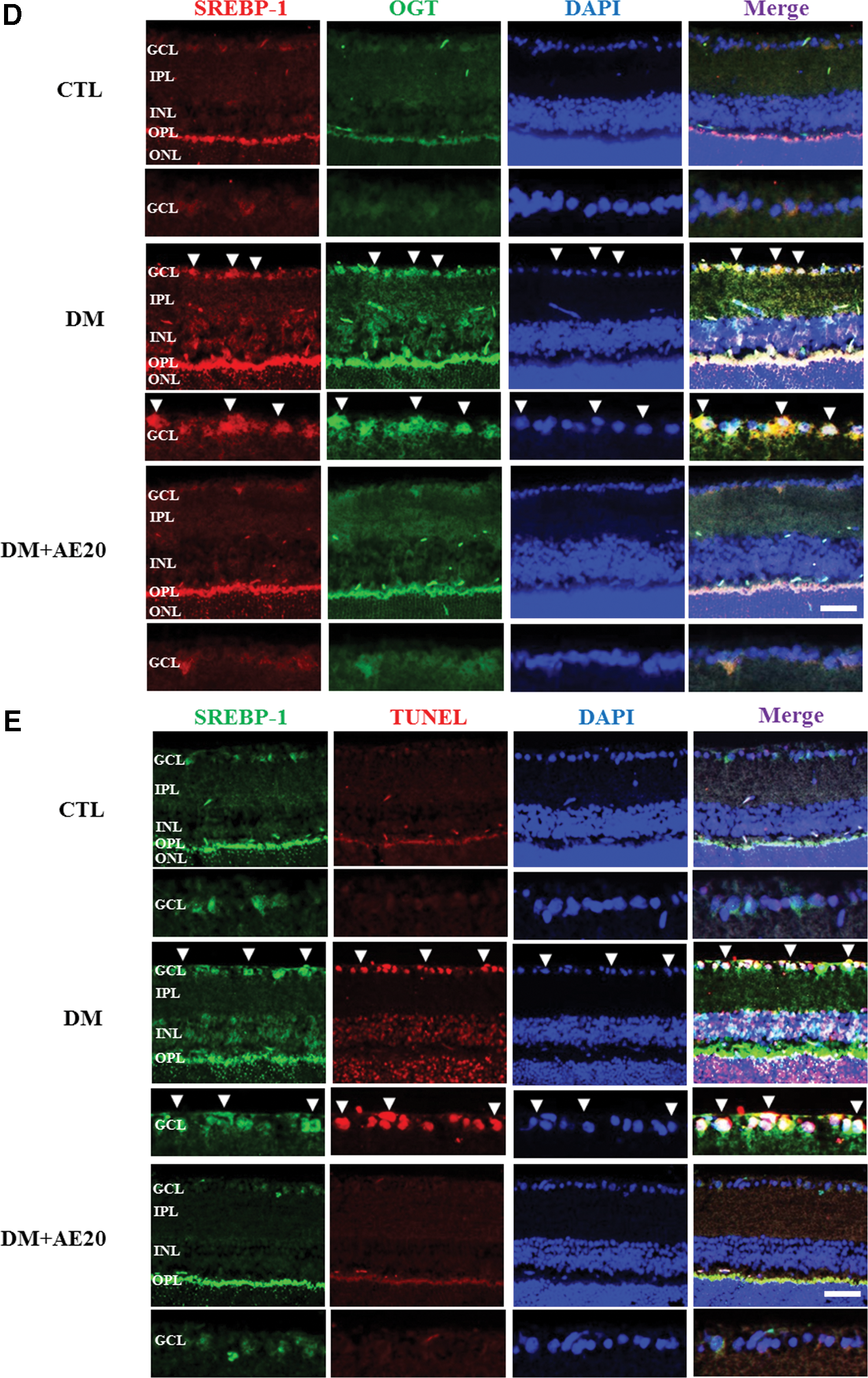
SREBP-1c expression is transcriptionally stimulated by insulin and is itself, in turn, a major mediator of insulin action. 31 Therefore, we assessed whether SREBP-1 is involved in retinal cell death in DR and whether AES affects SREBP-1 in the diabetic retinas. SREBP-1 is synthesized as a precursor form, and after proteolytic cleavage, its mature active form migrates into the nucleus where it can bind both sterol regulatory elements and E-boxes. 32 Western blotting analysis showed that levels of both the precursor and mature SREBP-1 protein were significantly increased in the diabetic retinas compared with that in controls (P < .05; Fig. 4A). However, AES treatment significantly reversed this increase in precursor and mature SREBP-1 in the diabetic retinas (P < .005 and P < .05, respectively; Fig. 4A), suggesting that AES significantly inhibits activation of SREBP-1. SREBP-1 also activates the transcription of lipogenic genes such as FAS 33 and ACC. 34 Next, we evaluated the effect of AES on FAS and ACC. Indeed, we found that levels of FAS and ACC were significantly increased in the diabetic retinas, compared with that in controls (P < .05 and P < .005, respectively; Fig. 4B, C), These data suggest increased activity of SREBP-1 in diabetic mice, which may, in turn, promote insulin action on FAS and ACC. However, AES treatment significantly reversed the elevation in FAS and ACC levels in the diabetic retinas (P < .005 and P < .05, respectively; Fig. 4B, C). Because activated AMP-activated protein kinase (AMPK) may reduce ACC and suppress lipogenic enzymes such as SREBP-1, 35 our results suggest that AES activates AMPK in the diabetic retinas, which would eventually lower SREBP-1 levels. Moreover, we found that SREBP-1 colocalizes with OGT and the TUNEL signal (Fig. 4D or E, respectively), suggesting that SREBP-1 may be O-GlcNAc modified or contribute to GCL cell death in the diabetic retinas. Intriguingly, SREBP-1 colocalization with OGT and the TUNEL signal was greatly reduced in the retinas of diabetic mice treated with AES (Fig. 4D or E, respectively).
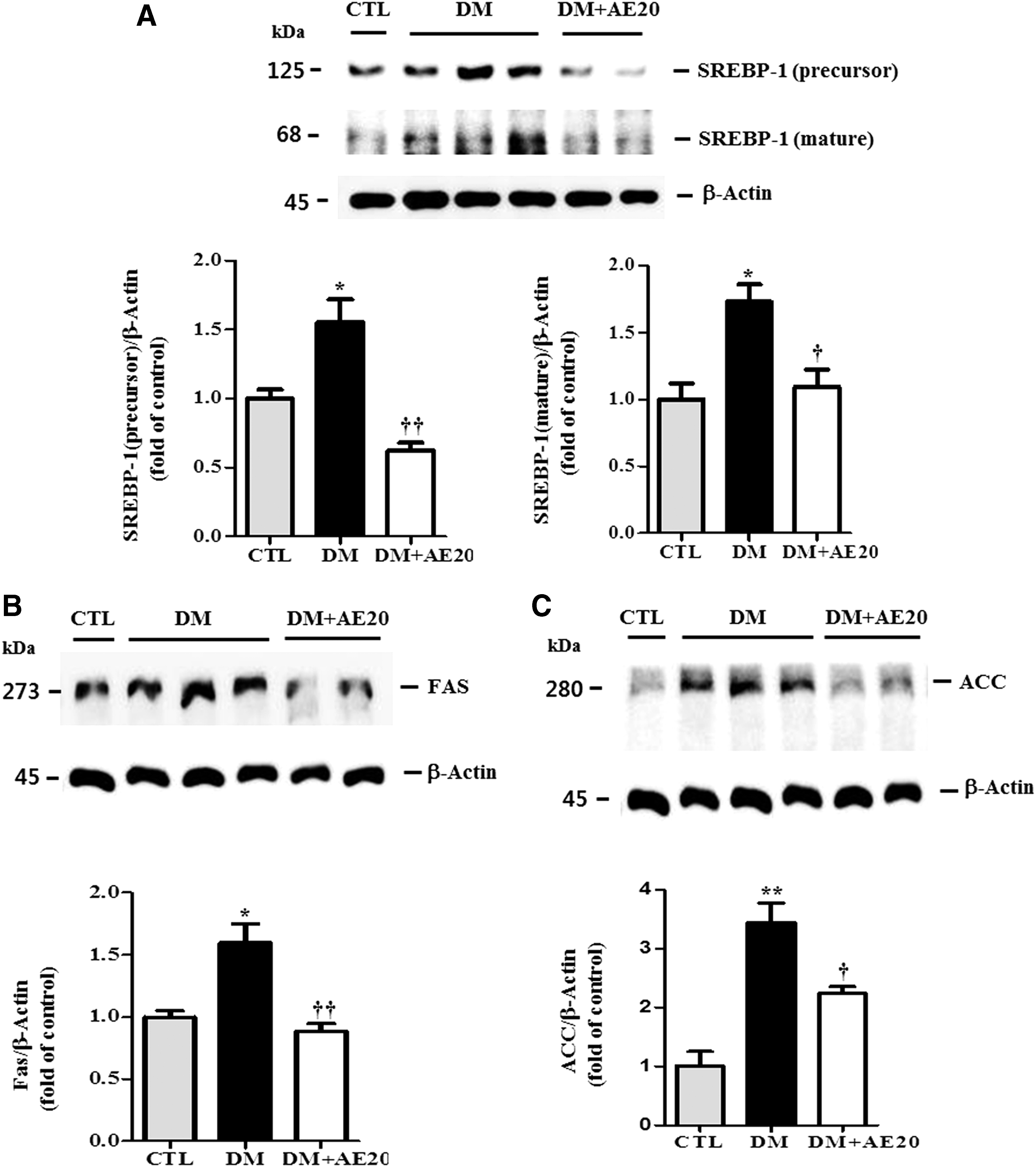
Representative Western blot and quantification of SREBP-1
AES inhibits retinal cell death through OGT
OGT is associated with insulin resistance 36 and may trigger the programmed cell death in diabetes mellitus and neurodegeneration. 37 Based on this role of OGT in diabetes, we hypothesized that OGT promotes retinal cell death in the inner retinal layers, which is an early characteristic of diabetes that eventually progresses to retinopathy. 38,39 To test whether AES affects OGT-related cell death, we first assessed OGT immunoreactivity in the GCL of the diabetic retinas. We found that OGT immunoreactivity in the GCL of the diabetic retinas was greatly increased compared with that in controls, while AES treatment restored this diabetes-associated increase (Fig. 5). Moreover, TUNEL-positive ganglion cells were also significantly increased in the diabetic retinas, compared with that in controls, and AES treatment decreased the numbers of dying ganglion cells in the diabetic retinas (Fig. 5). Double immunofluorescence analysis showed that OGT colocalized with most TUNEL-positive ganglion cells in the GCL of diabetic retinas, while AES treatment significantly reduced the OGT signal in TUNEL-positive regions (Fig. 5). Sequential double-immunofluorescent staining for γ-Syn, the ganglion cell marker, and OGT was performed. Indeed, most of the TUNEL-positive and OGT signal also colocalized with γ-Syn, confirming that the OGT signal was coincident with ganglion cells undergoing apoptosis.
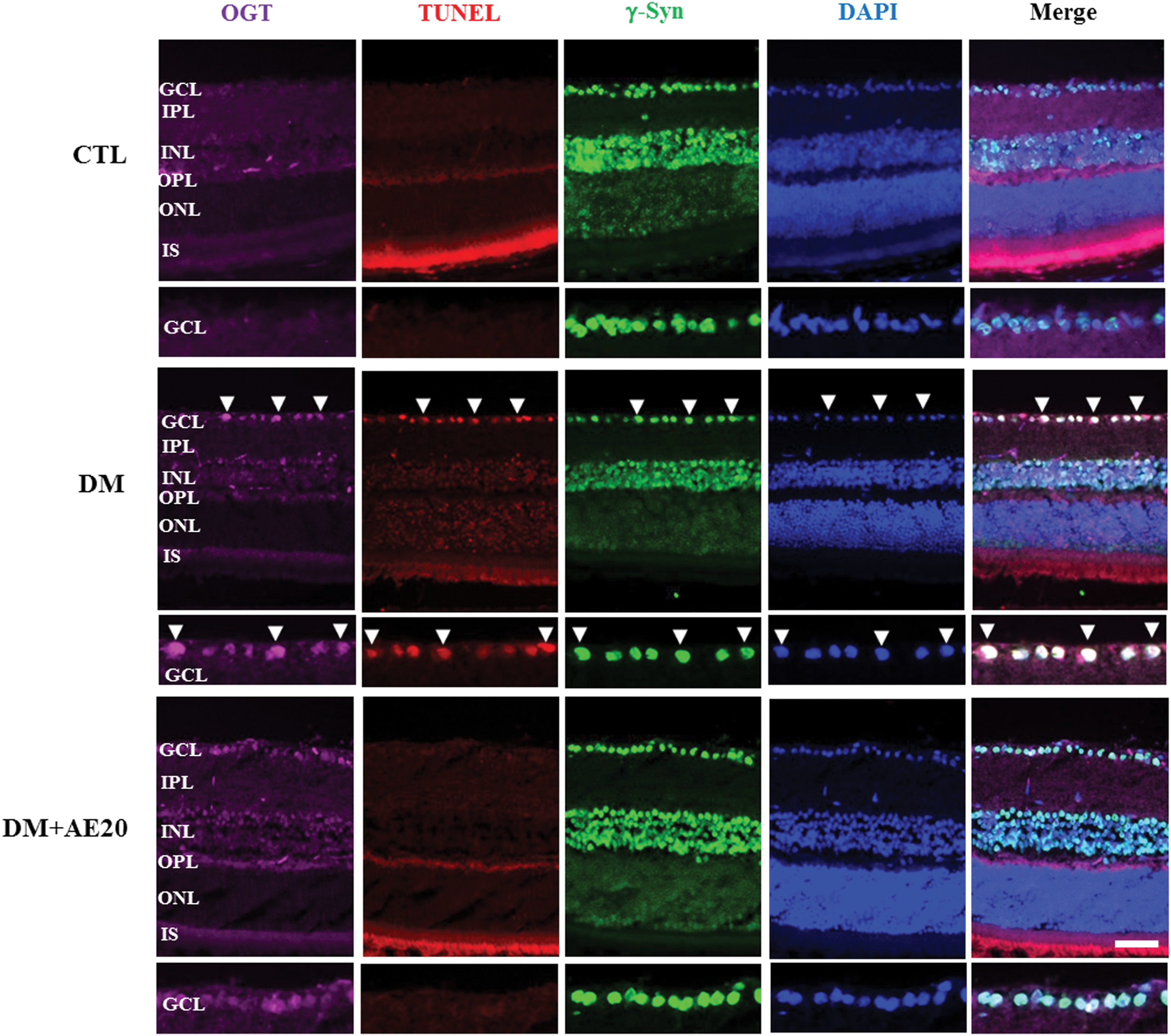
Effect of AES on OGT and ganglion cell death in the retinas of diabetic mice. Color images available online at
Discussion
O-GlcNAc dysregulation is linked to diabetes, 40,41 while OGT triggers apoptosis in diabetes. 37 Hyperglycemia promotes the rapid and reversible accumulation of O-GlcNAc protein, specifically in beta cells in vivo, and animals pretreated with STZ also accumulate O-GlcNAc protein in their beta cells when hyperglycemic. 42 In this study, we provide new evidence for increased OGT protein expression in the retinas of diabetic mice. Our data also show that AES treatment reversed in the diabetes-associated OGT increase and protected the retinas of diabetic mice from neuronal cell death in the GCL. OGT may play a role in triggering the programmed cell death observed in diseases such as diabetes mellitus and neurodegeneration. 37 Our data showed that OGT was related to the cell death in the GCL of the diabetic retinas, as indicated by double immunofluorescence analysis of OGT and the TUNEL assay. Our finding that AES administration reduced retinal cell death related to OGT in diabetic mice is of particular note, as this finding supports further development of AES as a protective agent in DR.
We also found that ChREBP protein and its colocalization with the TUNEL signal in the GCL of the diabetic retinas was increased. Importantly, AES reduced levels of ChREBP, which is known to play a crucial role in apoptosis 18 and is correlated with insulin sensitivity in early type 2 diabetes. 43 Because ChREBP is involved in metabolic adaptation to changing glucose levels, and alterations in glucose homeostasis are associated with metabolic diseases such as type 2 diabetes, 18 reduction of ChREBP by AES will likely uncover new therapeutic opportunities in DR. Moreover, treatment with AES downregulated TXNIP, consistent with a recent study where AMPK was found to decrease TXNIP protein levels. 44 AMPK activation has been suggested to be a good target in treatment of type 2 diabetes. 45 Therefore, the similarity in the effects of AES and AMPK are intriguing. Also, a previous study showed that increased beta cell TXNIP expression induced by high glucose plays a major role in the pathogenesis of pancreatic beta cell loss and diabetes. 24 As such, we found that TXNIP was increased and colocalized with TUNEL-positive ganglion cells in the GCL of the diabetic retinas, and these diabetes-associated changes could be attenuated by treatment of the diabetic mice with AES. Furthermore, ChREBP has been shown to interact with OGT and is O-GlcNAc modified in HEK293T cells and mouse hepatocytes. 21 Consistent with these data, we found that ChREBP interacts with and is colocalized with OGT as well as with TUNEL-positive GCL of the diabetic retinas. Our data also show that ChREBP contributes to neuronal cell death of the diabetic retinas. Strikingly, AES restored the extent of the ChREBP-OGT interaction, suggesting that AES reduces GCL cell death through its effects on ChREBP and OGT. In response to elevated glucose, ChREBP translocates to the nucleus. 46 We detected nuclear ChREBP in the diabetic retinas, suggesting that ChREBP is activated by high glucose, whereas AES reverses this activation. Previous studies also showed that ChREBP acts in synergy with SREBP-1 and is necessary for the induction of lipogenic genes such as ACC and FAS in response to glucose and insulin. 47 –49 Notably, SREBP has a critical role in the pathogenesis of the metabolic syndrome and controlling pathways for insulin resistance. 26,50 Intriguingly, AES reduced levels of activated SREBP-1 in the diabetic retinas, and concomitantly lowered levels of FAS, a known target of SREBP, consistent with a previous report. 25 Because SREBP-1 has been implicated in type 2 diabetes 26 and the pathogenesis of β cell dysfunction, 51 our results suggest that SREBP-1may be a novel therapeutic target in DR, and that AES can inhibit SREBP-1. Moreover, AES-mediated decrease in SREBP-1 levels may indicate activation of AMPK, because activated AMPK suppresses SREBP-1. 35 Therefore, AES-mediated reduction in SREBP-1 suggests that AES could be a useful therapeutic agent in DR. Therefore, the combined potent effects of AES on FAS and ACC, an action likely induced synergistically by ChREBP and SREBP-1, warrant further studies of this unique antidiabetic agent.
STZ, a GlcNAc analog, specifically blocks the enzyme that cleaves O-GlcNAc from intracellular proteins, and therefore, STZ toxicity is a result of the sustained accumulation of O-GlcNAc after a hyperglycemic episode. 42 These data suggest that the diabetogenic effect of STZ may enhance OGT action.
In conclusion, our findings demonstrate the neuroprotective action of AES extract in the retinas of this mouse model of diabetes, and provide support for the role of AES as a novel inhibitor of OGT, an important regulator of retinal cell death in the GCL of diabetic mice. Our data suggest that the neuroprotective function of AES in DR is mediated at least partially through ChREBP. Further studies are needed to elucidate the molecular pathway(s) invoked by AES and to address its utility as a potential therapeutic agent in DR.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation (NRF) of Korea funded by the Ministry of Science, ICT, and Future Planning (2014049413) and NRF-2015R1A5A2008833.
Author Disclosure Statement
No competing financial interests exist.