
Abstract
Sarcopenia, loss of muscle mass and function, is mainly observed in elderly people. In this study, we investigated whether fermented rice germ extract (FRGE) has some effects on the mouse gastrocnemius muscle by using behavioral and morphological analyses, Western blotting, and a murine model of immobilization-induced muscle atrophy. Daily oral FRGE administration increased muscle weight and strength. In addition, myofiber size in gastrocnemius muscle of FRGE-treated mice was increased as revealed by morphological quantification. Activation of AMP-activated protein kinase (AMPK) signaling, which inhibits protein synthesis and stimulates protein degradation in gastrocnemius muscle, was significantly attenuated in the FRGE-treated mice compared with control mice. Expression level of forkhead box 3a (FOXO3a) protein was also significantly decreased in the FRGE-treated group. Moreover, the decrease in mean myofiber cross-sectional area in immobilized hindlimb in vehicle-treated mice was inhibited by FRGE treatment in histological analysis. In conclusion, FRGE increased the strength and weight of gastrocnemius muscle and myofiber size, and reduced immobilization-induced muscle atrophy in mice. These findings indicated that FRGE might be beneficial in preventing motor dysfunction in a range of conditions, including sarcopenia.
Introduction
S
Sarcopenia is proposed to be causally related to changes in hormones and cytokine levels with age, 4 and an imbalance between rates of protein synthesis and degradation causes muscle loss. 2,5 Although a number of studies indicated that interventions incorporating nutritional supports and resistance exercise might be useful for prevention and/or treatment of sarcopenia, 6 –9 effective approaches have not yet been established. It is well known that AMP-activated protein kinase (AMPK) signaling and mammalian target of rapamycin (mTOR) signaling are key regulators of skeletal muscle protein turnover. 10 AMPK is well known as a key sensor of energy levels that activates metabolism when low cellular energy levels are detected. mTOR is a key regulator of protein synthesis and is essential for the maintenance of skeletal muscle function and mass. 11,12 AMPK and mTOR are both regulators of translation initiation process; however, they act in opposite ways. 13 AMPK also suppresses protein synthesis in rat skeletal muscle through the downregulation of mTOR signaling. 14 In this study, phosphorylation of AMPK was determined as protein degradation signaling.
Fermented rice germ extract (FRGE) is one of products obtained by the fermentation of rice germ. Rice germ is rich in a wide variety of nutrients, such as vitamins, minerals, dietary fibers, and γ-amino butyric acid (GABA). 15 Consequently, a number of physiological active substances that are isolated from rice germ are widely used in medications, health and functional foods, cosmetics, and food additives that include GABA 16 –18 and γ-oryzanol. 19 In addition, some of the bioactive constituents of rice germ following fermentation were shown to have beneficial health effects because fermentation enhances the nutritional value of food. 20 For example, α-ethylglucoside, which is found in FRGE, was shown to prevent ultraviolet B-mediated disruption of epidermal permeability barrier. 21 However, a role for FRGE or its constituents in maintaining gastrocnemius muscle has not been identified. Therefore, in this study, we investigated the impact of FRGE on gastrocnemius muscle using behavioral and histological analyses and Western blotting in mice. We also determined whether FRGE prevented gastrocnemius muscle atrophy in a murine unilateral immobilization model of skeletal muscle atrophy.
Materials and Methods
Plant materials and chemicals
Defatted rice germ (1.4 kg; Oryza Oil & Fat Chemical Co., Ltd., Aichi, Japan) was mixed with distilled water (1.4 L) and sterilized by autoclave (121°C, 15 min). Seed malt (560 mg) from Aspergillus oryzae (Bio'C Co., Ltd., Aichi, Japan) and 42 g of roasted barley flour (Owari Seifun, Inc., Aichi, Japan) were added to the rice germ, which was then fermented at 35°C for 3 days followed by extraction with methanol (3 L) at 70°C for 2 h. The extraction was repeated two more times. The extracted solution was then concentrated under reduced pressure to obtain FRGE (156.1 g, yield: 11.2%). Compounds in FRGE were isolated using the following procedure: FRGE (83 g) suspended in water (500 mL) was successively extracted with n-hexane (500 mL × 3), EtOAc (500 mL × 3), and CHCl3:MeOH (6:1; 500 mL × 3). The solvents were evaporated to obtain n-hexane fraction (17.7 g), EtOAc fraction (5.7 g) and CHCl3:MeOH fraction (3.7 g). n-hexane fraction (17.7 g) was separated with silica gel (130 g) column chromatography (n-hexane:EtOAc [9:1 → 4:1 → 7:3 → 3:2 → 1:1]→MeOH) to obtain fr. 1 (620 mg), fr. 2 (2.8 g), fr. 3 (894 mg), fr. 4 (441 mg), fr. 5 (228 mg), fr.6 (156 mg), and fr.7 (608 mg). Fr. 3 was purified by high performance liquid chromatography (HPLC) (Si, 20Φ × 250 mm; Fuji Silysia Chemical Ltd., Kasugai, Japan) with a mixture of n-hexane: EtOAc (1:1) to obtain stigmasterol (149.7 mg, yield: 0.18% from FRGE). 22 Fr. 4 was purified by HPLC (column: Si, 20Φ × 250 mm; Fuji silysia Chemical Ltd.) with a mixture of n-hexane: EtOAc (1:1) to obtain ergosterol peroxide (11.2 mg, yield: 0.013% from FRGE). 23 CHCl3:MeOH fraction (3.7 g) was separated with silica gel (160 g) column chromatography [EtOAc→CHCl3→CHCl3:MeOH (4:1 → 3:2 → 2:3 → 1:4)→MeOH] to obtain fr. 1 (299 mg), fr. 2 (8.4 g), fr. 3 (449 mg), fr. 4 (277 mg), fr. 5 (90 mg), and fr.6 (126 mg). Fr. 3 was purified by HPLC (column: [TSK-GEL Silica-60, 10Φ × 250 mm; Tosoh, Tokyo, Japan] and [Develosil C30 UG-5, 10Φ × 250 mm; Nomura Chemical Co. Ltd., Aichi, Japan] were connected in tandem) with CHCl3:MeOH:water (99:11:1). Then, we obtained β-sitrosterol-3-O-glucoside (4.4 mg, yield: 0.005% from FRGE) 22,24 and 1-O-glucosyl-2-(2-hydroxicosanoylamino)-4,8-octadecadience-1,3-diol (glucosylceramide [18:2]) (4.1 mg, yield: 0.005% from FRGE). 25 Each compound was identified by 1 H- and 13 C-(nuclear magnetic resonance) NMR spectra with a value in reference.
Sample treatment
FRGE (Oryza Oil & Fat Chemical Co., Ltd.) was dissolved in 0.5% carboxymethylcellulose sodium salt (Wako Pure Chemical Industries, Ltd., Osaka, Japan) and administrated orally at doses of 100 and 500 mg/kg/day every afternoon for 10 or 19 days. In the control group, 0.5% carboxymethylcellulose vehicle at 10 mL/kg was administrated. FRGE was treated at the doses of 100 and 500 mg/kg/day, as the previous report, which demonstrated that leucine supplementation extended muscle protein synthesis in rat. 26
Animals
Six-week-old male ICR mice (Japan SLC, Inc., Shizuoka, Japan) were used. All mice were housed at 24°C ± 2°C under a 12-h light/12-h dark cycle (lights on from 8:00 to 20:00) and had ad libitum access to food and water. All experiments were approved and monitored by the Institutional Animal Care and Use Committee of Gifu Pharmaceutical University and were performed after approval by the animal experimentation committee of Gifu Pharmaceutical University (Ethic Nos. 2015-072, 2015-170, 2016-108). All efforts were made to minimize both animal suffering and the number of animals used. All behavioral tests were performed between 13:00 and 17:00. For all tests, mice were brought to the experimental room more than 1 h before tests. The protocol is shown in Figure 1.

Protocol for FRGE administration and behavioral tests. FRGE or vehicle was orally administrated for 19 days. The potential effects of FRGE on muscle function were evaluated by a variety of behavioral tests during the last 5 days of treatment. FRGE, fermented rice germ extract; GST, grip strength test; HBT, horizontal bar test; OFT, open field test; RR, rotarod; RRT, rotarod test.
Induction of muscle atrophy
Procedures for the murine model of muscle atrophy were previously described. 27,28
Mice were anesthetized with 2.0–3.0% isoflurane (Mylan, Canonsburg, PA, USA) in 70% NO2/30% O2 and maintained with 1.0–1.5% isoflurane in 70% NO2/30% O2 delivered by an animal general anesthesia machine (Soft Lander; Sin-ei Industry Co. Ltd., Saitama, Japan). One randomly chosen hindlimb was immobilized with plastic tube, clay, adhesive bandage, and plastic tape. Mice were monitored daily for signs of chewing and removed plaster. In mice with removed or loosed immobilization, the hindlimb was immobilized once again. Every afternoon, during the immobilization period, the mice were orally administrated 500 mg/kg/day FRGE or 0.5% carboxymethylcellulose vehicle.
Open field test
Mice were placed in an open field apparatus (length 30 cm × width 30 cm × height 30 cm) and allowed to move freely for 10 min. Total distance moved was recorded using the computer-operated Etho Vision XT system (Noldus, Wageningen, the Netherlands).
Grip strength test
Mice were acclimated to the apparatus five times before data collection and allowed to grasp the grid with forelimbs. The maximum pressure exerted was measured using a bar attached to digital force gauge FG-5005 (Sato Shoji, Inc., Kawasaki, Japan). All mice were tested five times with more than 30-sec intervals between trials, and the highest score was recorded.
Rotarod test
Mice were first acclimated to the Rotarod apparatus (Ugo Basile, Monvalle Varese, Italy) for 1 day before test initiation. During acclimation, the mice were placed on the rotarod for 5 min, at a constant speed of 4 rpm. During testing, mice were placed on the rotarod for five consecutive trials per day for 3 days. The rotarod was accelerated from 4 to 80 rpm for a maximum score of 220 sec, and latency to fall was recorded.
Horizontal bar test
The horizontal bar test was performed as described previously. 29 Metal bars (diameter, 2 and 5 mm) were placed 50 cm above the floor. Mice were allowed to grasp the bar, and latency to fall from the bar was measured. Mice that failed to grasp the bar properly or those that fell the first 5 sec were tested again, and scores over 5 sec were recorded. The criterion point was either a fall from the bar or the touch of forelimb with the edge of the bar. Maximum testing time was 60 sec. Scoring was as follows: 5, staying on the bar for 60 sec and the attainment at the edge of bar; 4, falling between 45 and 60 sec; 3, falling between 30 and 45 sec; 2, falling between 15 and 30 sec; and 1, falling between 5 and 15.
Histological analysis
A previous report has demonstrated that both soleus muscle and gastrocnemius muscle are likely to atrophy in elderly women. 30 Therefore, we measured gastrocnemius muscle fiber in this study. At the end of experiments for behavioral testing or immobilization, mice (8 weeks old) were perfused with saline to remove blood and gastrocnemius muscles were extracted after behavioral tests or immobilization period. Briefly, mice were anesthetized with sodium pentobarbital (150 mg/kg, intraperitoneal; Nakalai Tesque, Kyoto, Japan) and perfused with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for histological analysis. Muscles were enucleated and immersed in 4% paraformaldehyde for at least 24 h at 4°C. Next, samples were immersed sequentially in 70% ethanol (12 h), 90% ethanol (1 h), anhydrous ethanol (1 h, five times), xylene (1 h, three times), and melted paraffin (1, 1.5, and 2 h, sequentially). Paraffin-embedded samples were cut into 5-μm sections with a microtome (Leica, Tokyo, Japan). After drying overnight at 37°C, sections were stained with hematoxylin and eosin (Sigma-Aldrich, St. Louis, MO, USA). Images from slides were acquired by BZ-9000 HS all-in-one fluorescence microscope (Keyence, Osaka, Japan).
Cross-sectional area of gastrocnemius muscle
Two cross-sections were made at intervals of 1 mm from the edge of the gastrocnemius muscle. The cross-sectional areas of two samples at each position (2, 3, 4, 5, 6, and 7 mm) from the edge of the gastrocnemius muscle were measured in ImageJ (
Number and cross-sectional area of myofibers
The average of total number of myofibers was determined in two cross-sections prepared from the largest cross-sectional area of the gastrocnemius muscle. The cross-sectional areas of myofibers at the same position were also measured. Briefly, images of the central position of these sections were acquired, and the cross-sectional areas of ∼150 myofibers in the gastrocnemius muscle of each mouse were measured using ImageJ. Mean myofiber cross-sectional area and the distribution of the myofiber area were determined according to previously published methods. 32–33
Immunostaining
Paraffin-embedded sections (thickness, 5 μm) from the largest cross-sectional area of the gastrocnemius muscle were used for immunohistochemistry. Sections were incubated at 90°C for 20 min in Histo VT One (Nacalai Tesque), treated with 0.3% H2O2 in MeOH, and blocked in 5% skim milk (Wako Pure Chemical Industries, Ltd.) for 30 min. For immunostaining, sections were incubated at 90°C overnight with primary rabbit antibody against advanced glycation end products (AGEs, 1:500; Abcam, Cambridge, MA, USA). Next, sections were incubated at room temperature for 30 min with anti-rabbit IgG (1:200; Millipore Corporation, Bedford, MA, USA) and visualized with 3,3′-diaminobenzidine tetrahydrochloride (DAB; Wako Pure Chemical Industries, Ltd.) in the presence of 0.3% H2O2. At the end of immunostaining, the sections were counterstained with hematoxylin for 30 sec.
Western blotting
Mice were euthanized with decapitation, gastrocnemius muscles were removed and immediately frozen in liquid nitrogen, and stored at −80°C. Samples were lysed in the radioimmunoprecipitation buffer (50 mM Tris-HCl, 150 mM NaCl, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, and 1% Igepal CA-630), supplemented with protease inhibitors (Sigma-Aldrich) and phosphatase inhibitor cocktails 2 and 3 (Sigma-Aldrich). Lysates were centrifuged at 12,000 g for 20 min, and the supernatants were stored at −80°C until analysis. Protein concentrations were determined with the BCA protein assay kit (Pierce Biotechnology, Rockford, IL, USA) with bovine serum albumin as the standard. Equal volumes of lysate and sample buffer containing 20% 2-mercaptoethanol (Wako Pure Chemical Industries, Ltd.) were mixed, and samples were separated by 5–20% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (Wako Pure Chemical Industries, Ltd.). Proteins were transferred to polyvinylidene difluoride membrane (Immobilon-P; Millipore Corporation). For immunoblotting, the following primary antibodies were used: rabbit anti-phosphorylated mTOR (Ser2448; 1:1000; Cell Signaling Technology, Danvers, MA, USA), rabbit anti-mTOR (1:1000; Cell Signaling Technology), rabbit anti-phosphorylated AMPK (1:1000; Cell Signaling Technology), rabbit anti-AMPK (1:1000; Cell Signaling Technology), rabbit anti-forkhead box 3a (anti-FOXO3a; 1:1000; Cell Signaling Technology), rabbit anti-eIF4E (1:1000; Cell Signaling Technology), rabbit anti-phospho eIF4E (S209) (1:1000; Cell Signaling Technology), and rabbit anti- glyceraldehyde 3-phosphate dehydrogenase (GAPDH) monoclonal antibody (1:1000; Cell Signaling Technology). Primary antibodies were diluted in Can Get Signal Solution 1 (Toyobo, Osaka, Japan), and incubated at 4°C overnight. The secondary antibodies used were goat anti-rabbit or anti-mouse horseradish peroxidase conjugated (Thermo Fisher Scientific, Waltham, MA, USA). Secondary antibodies were diluted in Can Get Signal Solution 2 (Toyobo), and incubated at room temperature for 1 h. Immunoreactive bands were visualized using ImmunoStar LD chemiluminescence reagent (Wako Pure Chemical Industries, Ltd.), and band intensities were measured with the LAS-4000 image analyzer (Fuji Film, Tokyo, Japan).
Statistical analysis
Data are presented as mean ± statistical error of means. Statistical comparisons were made using one-way analysis of variance followed by Tukey's test with SPSS statistics (IBM, Armonk, NY, USA) software. In Figure 5E, Student's t-test was used for statistic analysis. P < .05 was considered statistically significant.
Results
Characterization of FRGE with methanol
FRGE obtained by methanol extract in this study contained a variety of nutritional constituents, mainly carbohydrates and proteins (Table 1). In this study, lipid-soluble constituents in FRGE were isolated and structurally analyzed by NMR, as lipid-soluble molecules easily pass cell membranes and exhibit physiological activity. NMR analysis showed that FRGE contained stigmasterol, ergosterol peroxide, β-sitosterol glucoside, and glucosylceramide (d18:2) (Fig. 2A–D, Table 2). Among the constituents of FRGE (Table 3), stigmasterol was a major compound among the isolated compounds.
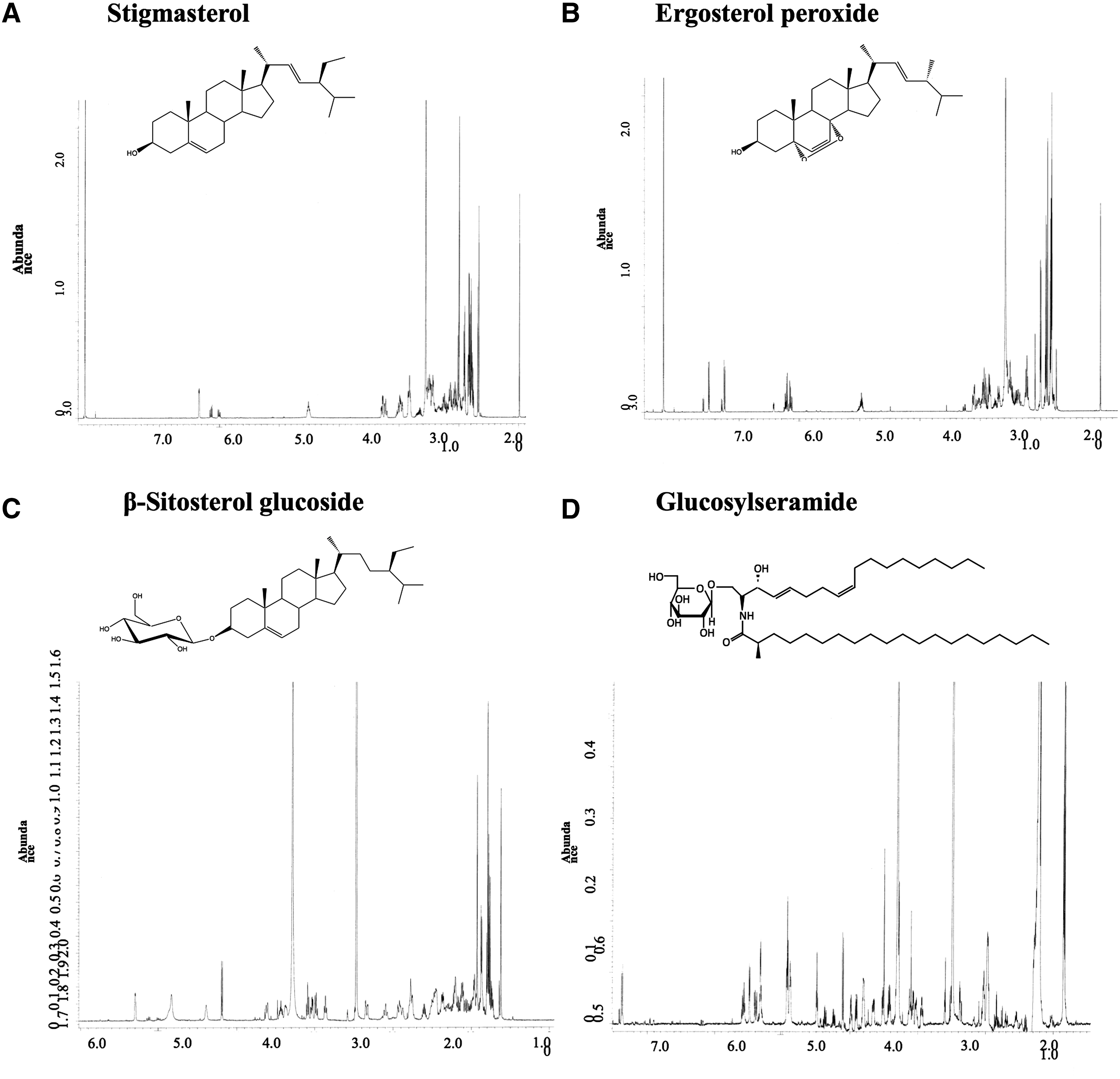
Identification of lipophilic constituents of FRGE.
FRGE contains a variety of nutritional components, mainly carbohydrate and protein.
FRGE, fermented rice germ extraction.
Fat-soluble constituents in fermented rice germ methanol extract were isolated and structures analyzed by NMR.
NMR, nuclear magnetic resonance.
Content and yields of each component from fermented rice germ methanol extract. In these components, stigmasterol was contained the most in fermented rice germ methanol extract.
FRGE increases muscle strength and muscle weight in mice
To investigate the effects of FRGE on motor function in mice, we performed several behavioral tests (Fig. 1). In the open field test, total distance moved was not different between the groups (Fig. 3A). In the grip strength test, FRGE treatment (100 and 500 mg/kg, per os [p.o.]) increased the grip strength score (Fig. 3B). In the rotarod test, no differences in the latency to fall were observed between the groups (Fig. 3C). In the horizontal bar test, latency to fall from the bar with a diameter of 2 mm did not change between the groups (data not shown). However, FRGE treatment (500 mg/kg, p.o.) tended to prolong the latency to fall from the bar with a bar diameter of 5 mm compared with untreated mice and increased the score of latency to fall (Fig. 3D). We also determined the effects of FRGE on body weight and muscle weight in mice. While total body weights did not change between the groups, gastrocnemius muscle weight in mice treated with 500 mg/kg FRGE significantly increased compared with that in untreated mice (Fig. 3E, F). Muscle weight per total body weight was also increased in mice treated with 500 mg/kg FRGE (Fig. 3G).
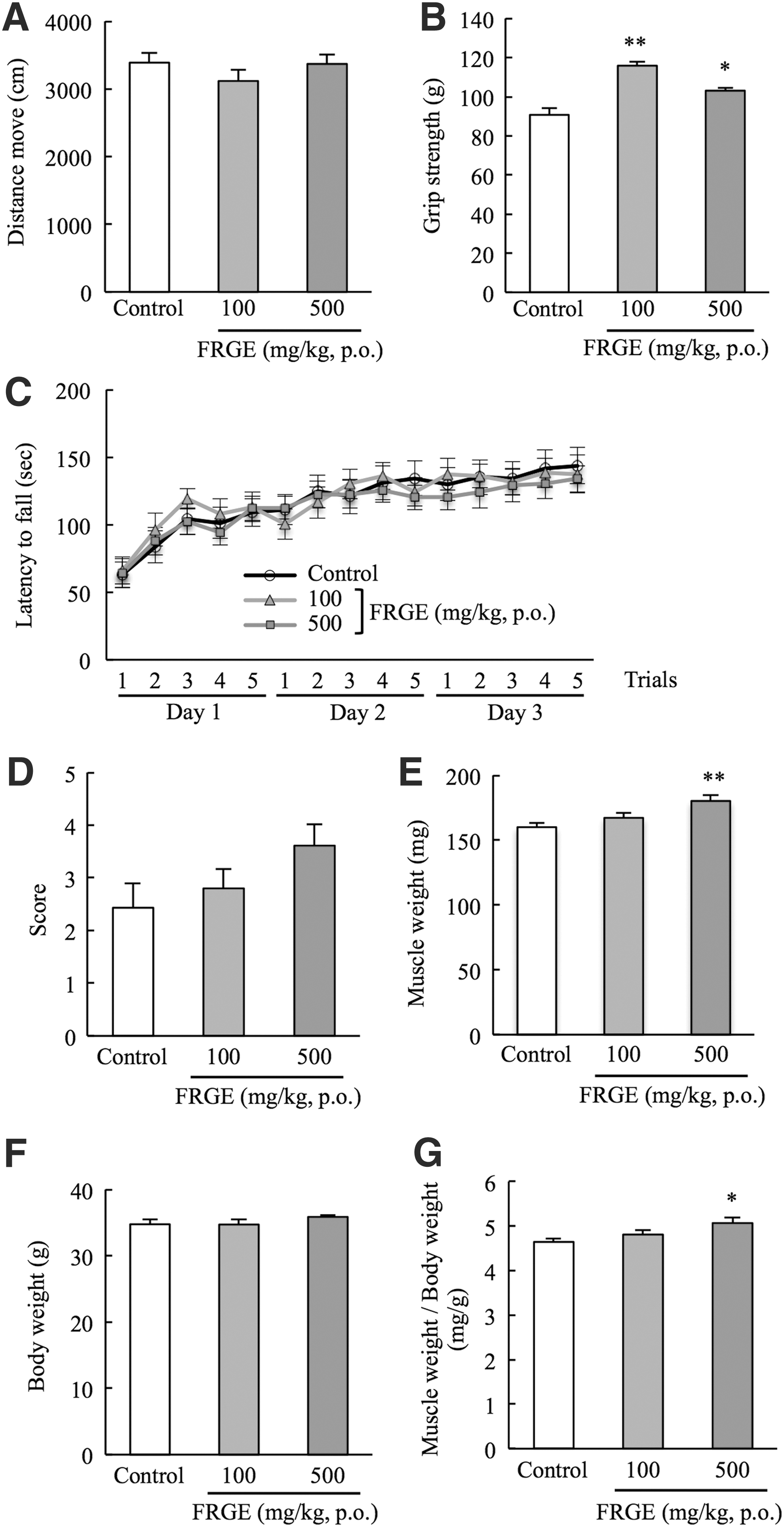
Effects of FRGE on muscle strength and muscle mass.
FRGE increases myofiber size in the mouse gastrocnemius muscle
We investigated the effect of FRGE on gastrocnemius muscle myofibers by histological analysis. FRGE treatment (100 and 500 mg/kg, p.o.) did not change cross-sectional area of each position from the edge of the muscle (Fig. 4A). We also determined the number of myofibers included in the cross-section 5 mm from the edge of the gastrocnemius muscle. Representative images of the gastrocnemius muscles are shown in Figure 4B. There was no difference in the number of myofibers between the groups (Fig. 4C). However, FRGE treatment (100 and 500 mg/kg, p.o.) significantly increased the mean myofiber cross-sectional area (Fig. 4D). The distribution of myofibers in FRGE-treated groups shifted to large-sized myofibers compared with the control group (Fig. 4E). In a previous study, the accumulation of AGEs in skeletal muscles of elderly was proposed as a potential mechanism of myofibrillar protein glycosylation underlying the age-related decline in muscle function. 34 Therefore, we investigated the effect of FRGE on expression of AGEs by immunostaining. However, there was no difference in the expression of AGEs between the groups (Fig. 4F).

Effects of FRGE on myofiber size.
FRGE inactivates the AMPK signal pathway in gastrocnemius muscle, but did not affect mTOR
To investigate the action mechanism of FRGE, we examined the phosphorylation of mTOR and AMPK by Western blotting, as mTOR and AMPK are key regulators of protein turnover in gastrocnemius muscle. Our results indicated that there was no difference in the phosphorylation of mTOR between each group (Fig. 5A, B). In contrast, the phosphorylation of AMPK was significantly decreased in the muscles of mice treated with 100 or 500 mg/kg FRGE (Fig. 5A, C). Moreover, FOXO3a, which is regulated by phosphorylated AMPK, was significantly decreased in the muscle of mice treated with 500 mg/kg FRGE (Fig. 5D, E). On the other hand, FRGE treatment did not affect the phosphorylation of eIF4E protein, which is regulated by mTOR and is involved in transfer initiation signaling (data not shown).
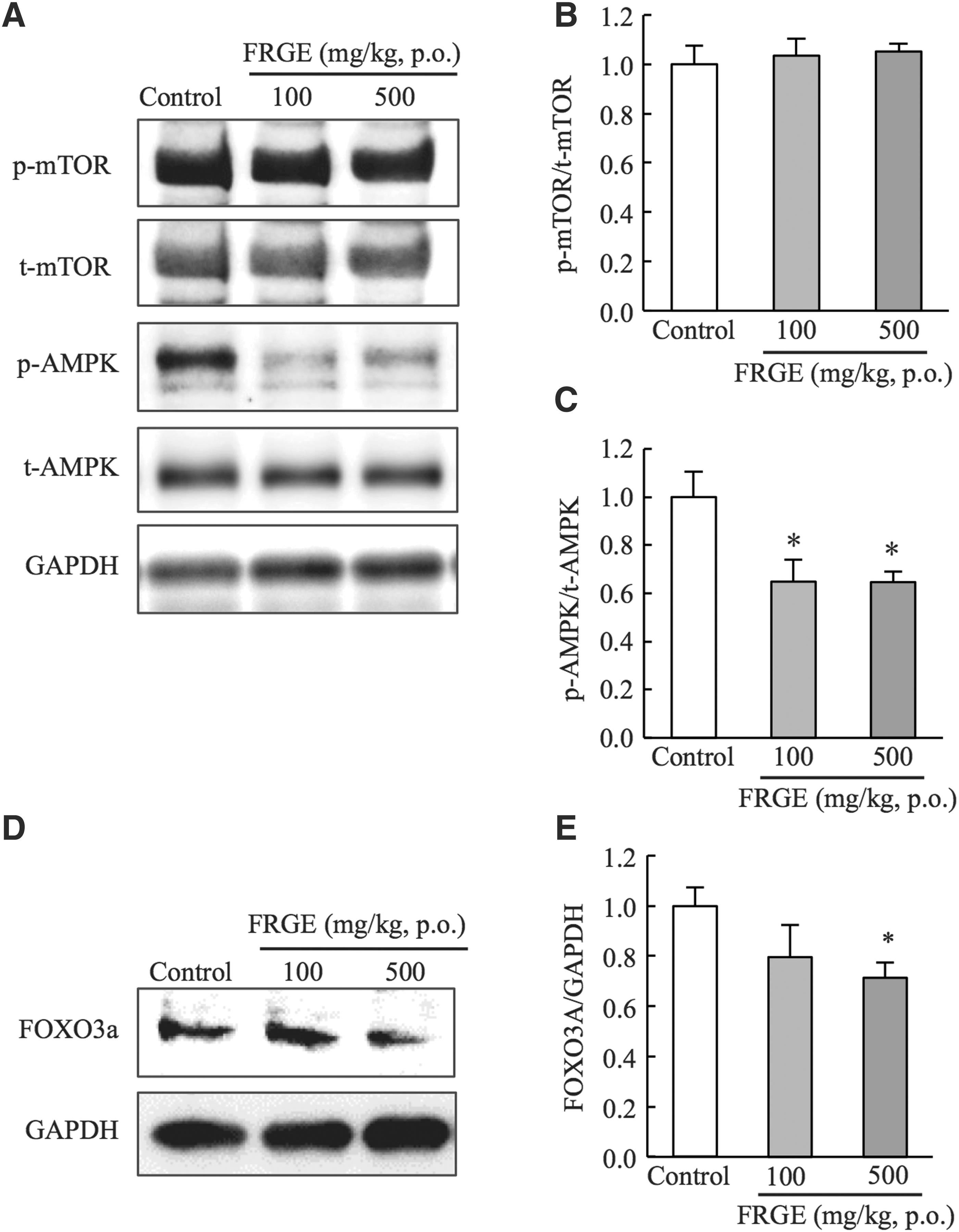
Effects of FRGE on AMPK activity and mTOR signaling pathway in skeletal muscle.
FRGE reduces immobilization-induced skeletal muscle atrophy
To investigate the effects of FRGE on gastrocnemius muscle atrophy, we administered FRGE at 500 mg/kg, p.o. to mice during unilateral hindlimb immobilization (Fig. 6A). We found that FRGE did not change muscle weight per total body weight (Fig. 6B). However, by histological analysis, FRGE treatment significantly inhibited the decrease in mean myofiber cross-sectional area in the immobilized hindlimb compared with the vehicle group (Fig. 6C, D).
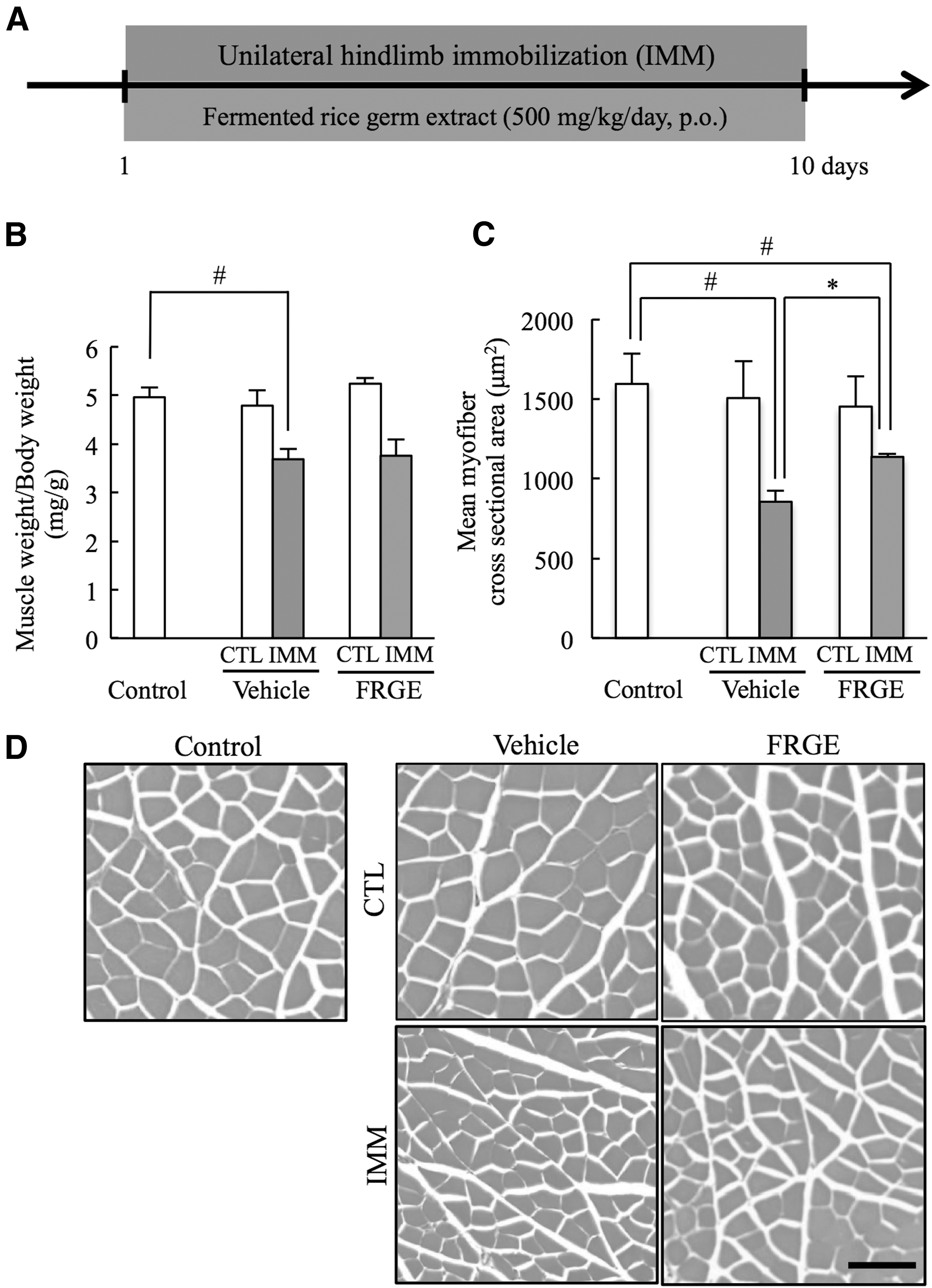
Effects of FRGE on immobilization-induced skeletal muscle atrophy.
Discussion
In this study, we investigated the effects of FRGE on muscle weight and function and also determined whether FRGE prevented gastrocnemius muscle atrophy in a murine unilateral immobilization model of skeletal muscle atrophy. We found that daily oral administration of FRGE increased muscle weights and muscle function, as well as myofiber size in mouse gastrocnemius muscle. Our analysis of lipid-soluble constituents of FRGE identified stigmasterol, ergosterol peroxide, β-sitrosterol-3-O-glucoside, and glucosylceramide (d18:2) (Fig. 2). Among these components, stigmasterol is also found in unfermented rice germ. Stigmasterol, ergosterol peroxide, and β-sitrosterol-3-O-glucoside are phytosterols, which are known to lower plasma cholesterol concentrations in both animals and humans 35 –37 and are also useful for treatment of benign prostatic hyperplasia. 38 Glucosylceramide (d18:2) was found to increase during the fermentation process 39 and improve transepidermal water loss in skin. 40 In this study, the identity of other FRGE constituents and the specific ingredients showing activity in skeletal muscle remain unclear. Therefore, further investigation is necessary to identify and characterize the bioactive FRGE constituents.
In the behavioral tests, we found that there were no differences in the total distance moved (Fig. 3A) in the open field test and latency to fall in the rotarod test (Fig. 3C) between the groups. Therefore, FRGE did not impact motor activity or the ability of motor coordination. Conversely, grip strength was significantly increased (Fig. 3B) and the score in the horizontal bar test tended to increase. These results indicated that FRGE improved muscle function. Furthermore, the mean weights of gastrocnemius muscles in mice treated with 500 mg/kg FRGE were significantly increased compared with those of untreated mice (Fig. 3E), whereas the total body weights did not change between the groups (Fig. 3F). Previous studies suggested that increased skeletal muscle mass was accompanied with loss of total body fat. 41 –43 Therefore, in this study, we could not rule out the possibility that FRGE increased muscle mass by decreasing fat mass.
Previous reports suggested that both the size and the number of myofibers were affected by aging. 2,44,45 Our results indicated that FRGE did not affect the cross-sectional area or the number of myofibers of gastrocnemius muscle (Fig. 4A, C), whereas FRGE significantly increased myofiber size (Fig. 4D, E) and inhibited the decrease in myofiber size (Fig. 6C, D). Therefore, we suggested that FRGE increased the muscle weight and the function as a result of increased myofiber size, but not due to an increase in myofiber number. As connective tissues increase with age, 46 –48 in contrast, skeletal muscle hypertrophy decreases with age 49 ; these findings altogether suggest that FRGE might act on myofibers and the connective tissues, but not the entire gastrocnemius muscle in mice.
Aging changes hormones and cytokine levels, 4 and alters the balance between rates of protein synthesis and degradation, 2,5 resulting in loss of muscle mass and subsequent physical dysfunction. A previous report suggested that phosphorylation levels of AMPK increased in the skeletal muscle of inactive frail older women. 50 AMPK directly regulates transcription factor FOXO3a. 51 FOXO3a transcription factor induces the atrophy-related ubiquitin ligase atrogin-1 and causes skeletal muscle atrophy. 52 Previous reports demonstrated that leucine supplements suppress muscle atrophy by reducing the AMPK activity. 26,53 These reports suggested that excessive AMPKα signaling is an important target for protein degradation in skeletal muscle atrophy. Therefore, we focused on AMPK signaling mediators as key regulators of protein turnover in gastrocnemius muscle in this study. AMPK acts in response to cellular energy levels and modulates turnover and mass in skeletal muscle. 10 AMPK is activated by a wide variety of cellular stresses that decrease adenosine triphosphate (ATP) levels, including exercise, nutrient deprivation, and hypoxia. 54 –56 AMPK activation increases the expression of FoxO transcription factors and stimulates the protein degradation by two pathways, ATP-dependent ubiquitin-proteasome system and autophagy-lysosomal pathway. 10 In this study, the phosphorylation of AMPK in the FRGE-treated group was significantly decreased compared with the control group (Fig. 5A, C). Moreover, expression level of FOXO3a was significantly decreased in the FRGE-treated group (Fig. 5D, E). These results suggested that FRGE treatment might inhibit protein degradation through AMPK and FOXO3a inactivation with a consequent increase in myofiber size (Fig. 7). For prevention and treatment of sarcopenia, regulation of AMPK activity, which mediates protein synthesis and degradation, might thus be critical.
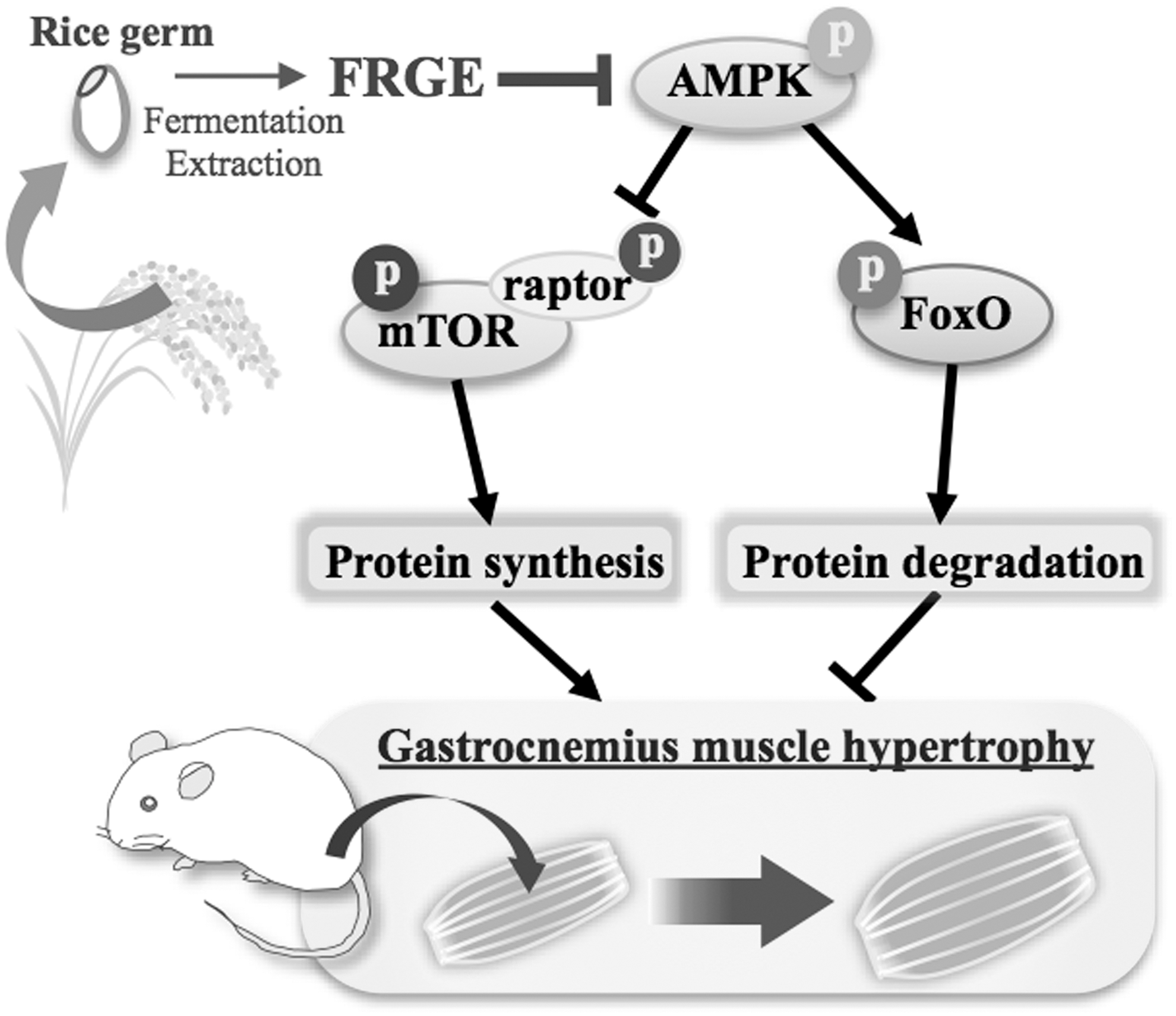
Mechanism for the action of FRGE. FRGE enhances murine gastrocnemius muscle strength by inactivation of AMPK.
On the other hand, we also investigated the phosphorylation levels of mTOR in gastrocnemius muscle after treatment with FRGE. The mTOR functions through two multiprotein complexes, mTOR complex 1 (mTORC1) and mTOR complex 2 (mTORC2). 57 mTORC1 signaling pathway has a wide range of functions, including regulation of protein synthesis, cell proliferation, apoptosis, and autophagy. 57 –59 To investigate whether FRGE stimulates the mTORC1 signaling pathway, we determined the phosphorylation of mTOR by Western blotting and found that p-mTOR levels were not different between the groups, suggesting that FRGE may not affect protein synthesis mediated by the mTORC1 signaling pathway (Fig. 5A, B). Also, the phosphorylation of eIF4E protein was not different between the groups, suggesting that FRGE treatment may not affect transfer initiation signaling. 60
In this study, we induced murine muscle atrophy by hindlimb immobilization. The amount of time required for immobilization-induced atrophy is much less than for age-related atrophy, although the mechanisms of muscle atrophy are the same. Hindlimb immobilization induces disuse muscle atrophy in a short period of time although muscle mass and function decrease gradually with age in sarcopenia. 1,2 Disuse muscle atrophy as well as sarcopenia have a primary effect on fast-twitch fiber, which accounts for a large portion of gastrocnemius muscle. 1,27 Therefore, we suggested that FRGE could enhance muscle function not only in disuse muscle atrophy but also in sarcopenia. In this study, we found that 500 mg/kg FRGE enhances the strength of gastrocnemius muscle in immobilization-induced skeletal muscle atrophy.
FRGE contained carbohydrate 52.89 g, protein 22.57 g, and fat 1.96 g/100 g, and normal meals (CLEA Rodent Diet CE-2; CLEA Japan, Inc.) contained carbohydrate 49.78 g, protein 24.88 g, and fat 5.03 g/100 g. Since doses of FRGE were clearly lower than the amount of normal meal, total amount of food intake might not be affected by FRGE treatment. On the other hand, previous reports demonstrated that branched-chain amino acid leucine supplementation (1 g/kg) affected protein synthesis of rat skeletal muscle. 26 Since FRGE contains protein, that protein might be a confounding effect for gastrocnemius muscle in this study; therefore, further analyses will be needed.
In conclusion, we demonstrated that FRGE increased muscle strength and weight, and the myofiber size, and reduced the immobilization-induced skeletal muscle atrophy in mice. Furthermore, FRGE decreased in the phosphorylation of AMPK. These findings indicated that FRGE might exert its beneficial effect by regulating protein turnover in gastrocnemius muscle and might be useful for the prevention and treatment of motor dysfunction observed in a number of conditions, including sarcopenia.
Footnotes
Acknowledgments
On the determination of chemical structures of isolated constituents, Professor Hisashi Matsuda and Dr. Seikou Nakamura in Department of Pharmacognosy, Kyoto Pharmaceutical University, supported our work. This research was supported by “Support business program for small and medium companies and small size enterprises developing new products and new technologies in 2013” from the Small and Medium Enterprise Agency, Japan. The funders had no role in study, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.