
Abstract
Acteoside, the predominant polyphenol of small-leaved kudingcha, the Chinese tea, has various biological activities. In this study, we examined the acyl migration of acteoside to isoacteoside with high-temperature treatment of acteoside. The inhibitory effects of acyl-migrated acteoside and acteoside on α-amylase were investigated, as were their binding interaction with α-amylase. The binding of acteoside and isoacteoside to α-amylase was investigated by using the fluorescence spectra assay, circular dichroism, and protein–ligand docking studies. Acteoside was more effective than preheated acteoside and isoacteoside in inhibiting α-amylase activity. Acteoside and isoacteoside binding to α-amylase may induce conformational changes to α-amylase, and the binding site of acteoside and isoacteoside being near the active site pocket of α-amylase may explain the decreased activity of α-amylase. The different affinities and binding sites of acteoside and isoacteoside for α-amylase resulted in different inhibition rates, which may be due to structural differences between acteoside and isoacteoside.
Introduction
I
Ligustrum purpurascens is a traditional medicinal plant common in southwestern China. 7 The leaves are used to make a beverage called small-leaved kudingcha (bitter tea), and were found to have health benefits. 8 In China, L. purpurascens kudingcha has long been used in folk medicine for lowering fat and blood sugar, excretion, and improving eyesight. L. purpurascens polyphenols, including the major components acteoside, osmanthuside, ligupurpuroside A, ligupurpuroside B, and ligupurpuroside C, have various physiological functions, such as antitumor, antibacterial, anti-inflammatory, and antiviral activities. 9 –14 Our previous study showed that acteoside is the predominant polyphenol of L. purpurascens and has extensive antioxidant activities. 15
Retarding carbohydrate digestion by inhibiting α-amylase may help to control postprandial blood glucose concentrations. Most studies on α-amylase inhibitors have focused on the utilization of plant-based foods. 16 –19 In a previous study, we identified the potential of phenylpropanoid glycosides from L. purpurascens for inhibiting α-glucosidase and α-amylase activity in vitro. 20 However, the detailed mechanism is not clear. In this study, we performed an in vitro study of α-amylase inhibition and evaluated the antihyperglycemic effects of acteoside. The binding of acteoside to α-amylase was studied by multispectroscopy and protein-ligand docking. With thermal treatments such as pasteurization, a considerable sum of polyphenols are epimerized or transformed. 21,22 We also examined whether preheating acteoside affects its inhibition of α-amylase.
Materials and Methods
Materials
Porcine α-amylase (Type I-A, 1000 U/mg) and 3,5-dinitrosalicylic acid were from Sigma Chemical (St. Louis, MO); acteoside and isoacteoside (purity >98%) were prepared in our laboratory. Preheated acteoside was prepared by autoclaving at 100 or 120°C for 20 min each.
High-performance liquid chromatography analysis of heated acteoside
High-performance liquid chromatography (HPLC) analysis of acteoside was as previously described, 23 with use of a Shimadzu LC-20AD HPLC system equipped with an SIL-20A autoinjector and SPD-M20A diode array detector (DAD) set to 227 nm (Shimadzu, Japan) and an Inertsil ODS-3 column (250 × 4.6 mm ID, 5 mm) fitted with a C18 guard column. Gradient elution of (A) water (containing 0.1% trifluoroacetic acid in water, v/v) and (B) methanol was as follows: 0.00–5.00 min, 35% B; 5.00–26.50 min, 35–42% B; 26.50–36.50 min, 42% B; 36.50–47.00 min, 42–44% B; 47.00–60.00 min, 44–51% B. The solvent flow rate was 1 mL/min. The column temperature was 30°C.
Liquid chromatography–mass spectrometry analysis
Liquid chromatography–mass spectrometry (LC-MS) data were obtained by use of Agilent Technology 6400 Series Triple Quadrupole LC/MS (1260 Infinity Binary LC; Agilent Technologies). The 1260 Infinity Binary LC instrument had a G1312B binary solvent-delivery system, G1379B thermostatted column compartment, G1316B vacuum degasser, G1367E autosampler, and G1315C DAD. Chromatography separation utilized an Inertsil ODS-3 column (250 × 4.6 mm ID, 5 mm) coupled with a C18 guard column at 30°C. The elution method was as described for RP-HPLC. An MS detector with electrospray ionization interface in positive ion mode (ESI+) was used for qualitative analysis, with acquisition in product ion scan mode. All parent and product ions were recorded. The optimized electrospray conditions were capillary voltage 4.00 kV; fragmentor: 250 V; source temperature: 100°C; desolvation temperature: 350°C; collision energy 40 V for all peaks; gas flow: 13 L/min; and nebulizer: 55 psi.
Enzyme activity and inhibitory pattern assay
The in vitro α-amylase activity assay involved the use of starch as a substrate, as described with slight modification. 24 In brief, 50 μL of 20 mM phosphate buffer (pH 7.0) containing α-amylase at 2 mg/mL and 25 μL of inhibitor (acteoside, preheated acteoside at 100°C, preheated acteoside at 120°C and isoacteoside) at various concentrations were incubated at 37°C for 10 min. Then, 10 μL of starch substrate solution (10 mg/mL in 20 mM phosphate buffer, pH 7.0) was added to an incubation medium to initiate the reaction. After incubation at 37°C for 30 min, 50 μL of 96 mM 3,5-dinitrosalicylic acid was added to stop the reaction. Then, the reaction mixture was incubated in boiling water for 5 min and cooled to room temperature. Absorbance at 540 nm was measured by a spectrophotometer. The percent inhibition of α-amylase was calculated as Inhibition rate% = (Abs control − Abs test)/Abs control × 100. All samples were analyzed in triplicate. The 50% inhibition (IC50) value was calculated.
Inhibition by acteoside and isoacteoside on α-amylase using the Michaelis–Menten constant (Km ) and maximum reaction velocity (Vmax ) was determined by Lineweaver–Burk plot analysis according to Michaelis–Menten kinetics. Acteoside and isoacteoside were measured at 0, 0.5, and 1 mg/mL with increasing concentrations of starch, 1, 3, 5, 7, and 9 mg/mL, as a substrate.
Fluorescence spectroscopy
Acteoside and isoacteoside solutions were prepared with 20 mM phosphate buffer (pH 7.0) at 0, 1.0, 2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, and 10.0 × 10−5 M, and then incubated with 1.0 × 10−5 M α-amylase at fixed temperatures (27 and 37°C, respectively) for 1 h. Fluorescence spectra were recorded under excitation at wavelength 295 nm in a 1-cm quartz cell by use of a Hitachi-850 spectrofluorometer (Hitachi Co., Japan). The emission and excitation bandwidths were 5 nm. The emission spectra were recorded from 300 to 450 nm.
Circular dichroism
The concentration of α-amylase was 1.0 × 10−6 M, and the α-amylase/inhibitor ratios in the complexes were 1:0, 1:0.5, 1:1, and 1:2. The samples were recorded with use of a JASCO-810 automatic spectrophotometer (JASCO Co., Japan) in cells of 1.0-mm path length at 25°C. The spectra were measured from 190 to 250 nm, with response time of 1 s and scan rate of 30 nm/min. Five scans were accumulated for each spectrum. The content of different secondary structures was analyzed by use of the online SELCON3 program. 25
Molecular docking studies
The 3D structure of porcine α-amylase was downloaded from the Protein Data Bank (accession no.: 1PIF). The 3D structure of acteoside and isoacteoside was generated by use of Chem3D Ultra 6.0, with MOE software. Acteoside and isoacteoside were considered fully flexible, with α-amylase as the receptor model and considered rigid in the virtual screening with docking simulations. The docked complex was optimized by its fit within the receptor pocket, for continuous and discrete Van der Waals interaction, electrostatics, hydrophobicity, hydrogen bonding, and entropy. Visualization and measurement of distances between the receptor and ligand involved use of PyMOL software.
Statistical analysis
Data are expressed as mean ± standard deviation (SD) and SDs calculated using Microsoft Excel (2007).
Results
When acteoside was preheated at 100 and 120°C, HPLC revealed that the acteoside peak gradually decreased and a new peak increased with heating (Fig. 1A). This new HPLC peak was at the same position as isoacteoside. LC/MS was used to identify the sample of acteoside preheated at 120°C, for a peak (peak 1) at 19.829 min and another (peak 2) at 28.196 min (Fig. 1B). The peak at 19.829 min showed an [M − H]− ion at 647.2 m/z in the ESI–MS spectrum (Fig. 1C). The peak at 28.196 min also showed an [M − H]− ion at 647.2 m/z (Fig. 1D). Isoacteoside is an isomer of acteoside, with molecular weight 647.2 kDa. Thus, peaks 1 and 2 were identified as acteoside and isoacteoside, respectively. The content of acyl-migrated acteoside is in Table 1. We found that the higher the temperature, the greater the acyl migration of acteoside to isoacteoside.

HPLC chromatograms of acteoside, preheated acteoside, and isoacteoside
Inhibition by acteoside, preheated acteoside, and isoacteoside
The inhibitory effects of acteoside, preheated acteoside, and isoacteoside against α-amylase are shown in Figure 2A. Inhibition of α-amylase activity increased with increasing concentrations of acteoside, preheated acteoside, and isoacteoside, with acteoside having a greater inhibitory effect than isoacteoside. According to IC50 values, the inhibitory effects on α-amylase were in the following order (Table 2): acteoside > preheated acteoside at 100°C > preheated acteoside at 120°C > isoacteoside. Isoacteoside conferred a relatively higher IC50 value, two times than did acteoside. Acteoside acyl that migrated to isoacteoside had a greater effect with the higher temperature. Therefore, preheated acteoside had a lower inhibitory effect than acteoside, and the higher the temperature, the lesser the inhibitory effect of acteoside on α-amylase.

Inhibition of α-amylase by acteoside, isoacteoside, and acteoside at 100 and 120°C
Data are mean ± SD (n = 3).
SD, standard deviation.
To further explore the inhibitory effect of acteoside and isoacteoside on α-amylase, we studied the kinetics of α-amylase with different concentrations of substrate and inhibitor. Lineweaver–Burk plots for acteoside and isoacteoside showed the same intersection on the Y-axis, so both were competitive inhibitors (Fig. 2B, C).
Fluorescence spectroscopy of α-amylase
The interactions of α-amylase with acteoside and isoacteoside were investigated by fluorescence quenching of α-amylase with the addition of acteoside and isoacteoside. On excitation at 295 nm, α-amylase showed a strong fluorescence emission with a peak at 334 nm because of its 19 tryptophan (Try) residues (Fig. 3A and C, data at 27 and 47°C not shown). The fluorescence intensity of α-amylase decreased gradually with the addition of acteoside and isoacteoside because acteoside and isoacteoside interacting with α-amylase leads to the unfolding of α-amylase Try residues, which causes quenching of the inherent fluorescence.
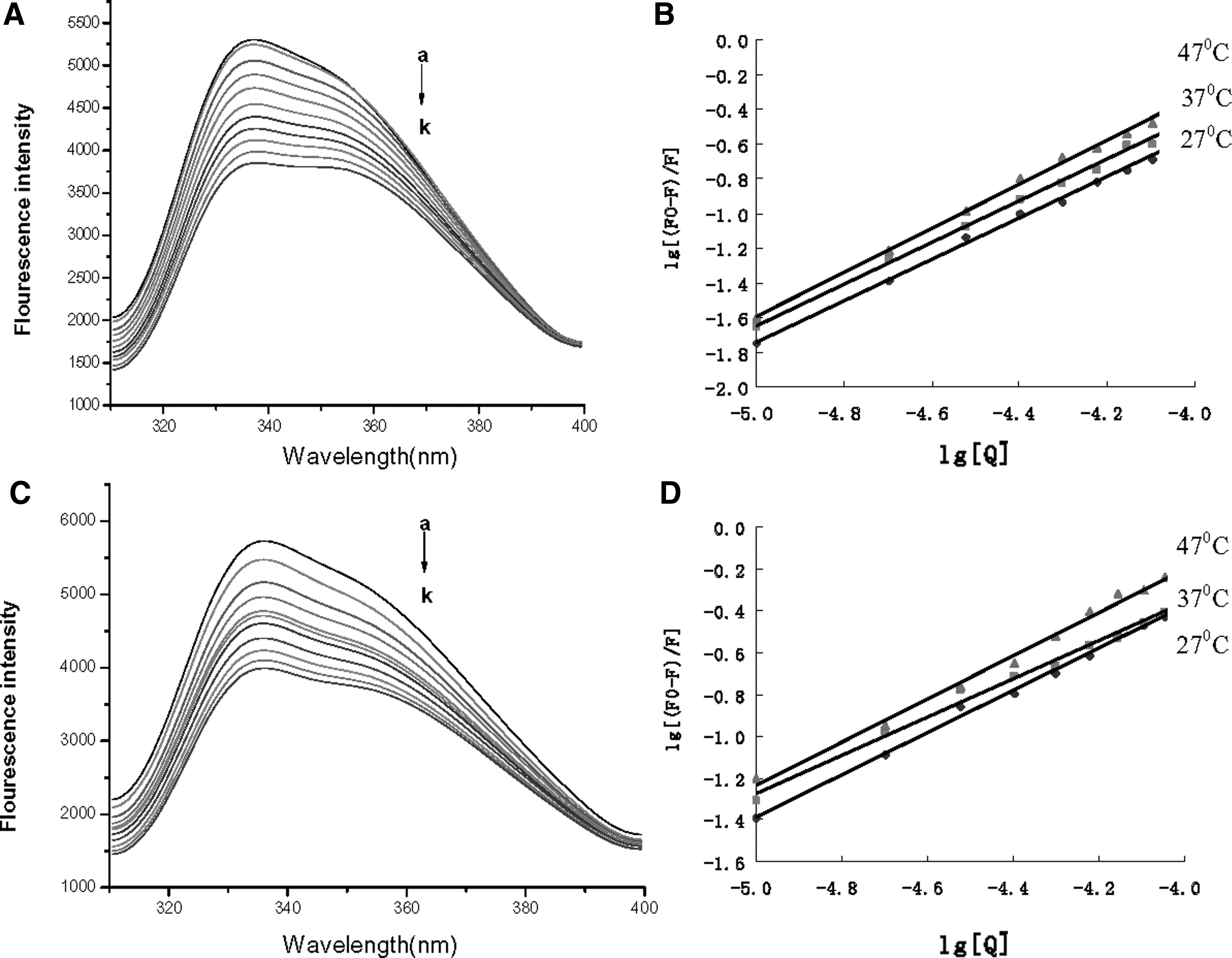
The quenching effect of acteoside
The fluorescence quenching mechanism can be described by the modified Stern–Volmer equation
26
:
where F 0 and F are the relative fluorescence intensities in the absence and presence of quencher, respectively; [Q] is the concentration of quencher; τ0 is the mean lifetime of the fluorophore; K SV is the Stern–Volmer quenching constant; K a is the binding constant; and n is the number of binding sites per protein. The equation was assessed by the double-logarithmic plot (Fig. 3B, D). The values for binding constants (K a) and number of binding sites per protein (n) from the double-logarithmic plot are shown in Table 3. The values of K a between acteoside and α-amylase were >1.0 × 104 L/mol, almost 1 order of magnitude for K a between isoacteoside and α-amylase. The results indicated a lower affinity of isoacteoside than acteoside for α-amylase.
The binding constants and number of binding sites per protein increased with increasing temperature, so temperature had a pronounced effect on the interactions of α-amylase with acteoside and isoacteoside. The increasing temperature may result in instability of the α-amylase–acteoside and α-amylase–isoacteoside system, but it also increased the diffusion coefficient. With increasing temperature, the increased diffusion coefficient of the α-amylase–acteoside and α-amylase–isoacteoside system played an important role in the interactions.
Change in entropy (ΔS) and enthalpy (ΔH) can be calculated from the following van't Hoff equation:
where T is experimental temperature, K is the binding constant at the corresponding T, and R is the gas constant.
The free energy change (ΔG) can be estimated as follows:
The values for the thermodynamic parameters are shown in Table 3. The negative sign for ΔG indicates the spontaneity of the binding of α-amylase with acteoside and isoacteoside. The ΔS and ΔH values are positive, so a hydrophobic interaction plays a major role in the α-amylase interaction with acteoside and isoacteoside.
Circular dichroism spectra
We used circular dichroism (CD) spectroscopy to study α-amylase conformational changes. The CD spectra for α-amylase showed two negative bands in the ultraviolet region at 209 and 226 nm (Fig. 4), characteristic of the α-helical structures in protein. 27 The absolute θ values were lower for the α-amylase–acteoside and α-amylase–isoacteoside complexes than native α-amylase at 209–218 nm, which suggests the absence of some α-helical structures in α-amylase. The secondary structure was determined by using SELCON3 in DICHROWEB. On binding with acteoside and isoacteoside, α-amylase showed decreased α-helical content, from 33.8% to 23.8% and 22.7%, respectively, with a slight increase in β-Sheet content from 16.7% to 20.5% and 21.1%, and the random coil changed from 49.4% to 55.7% and 56.2% (Table 4). Thus, the interaction of α-amylase with acteoside and isoacteoside caused conformational changes in α-amylase.
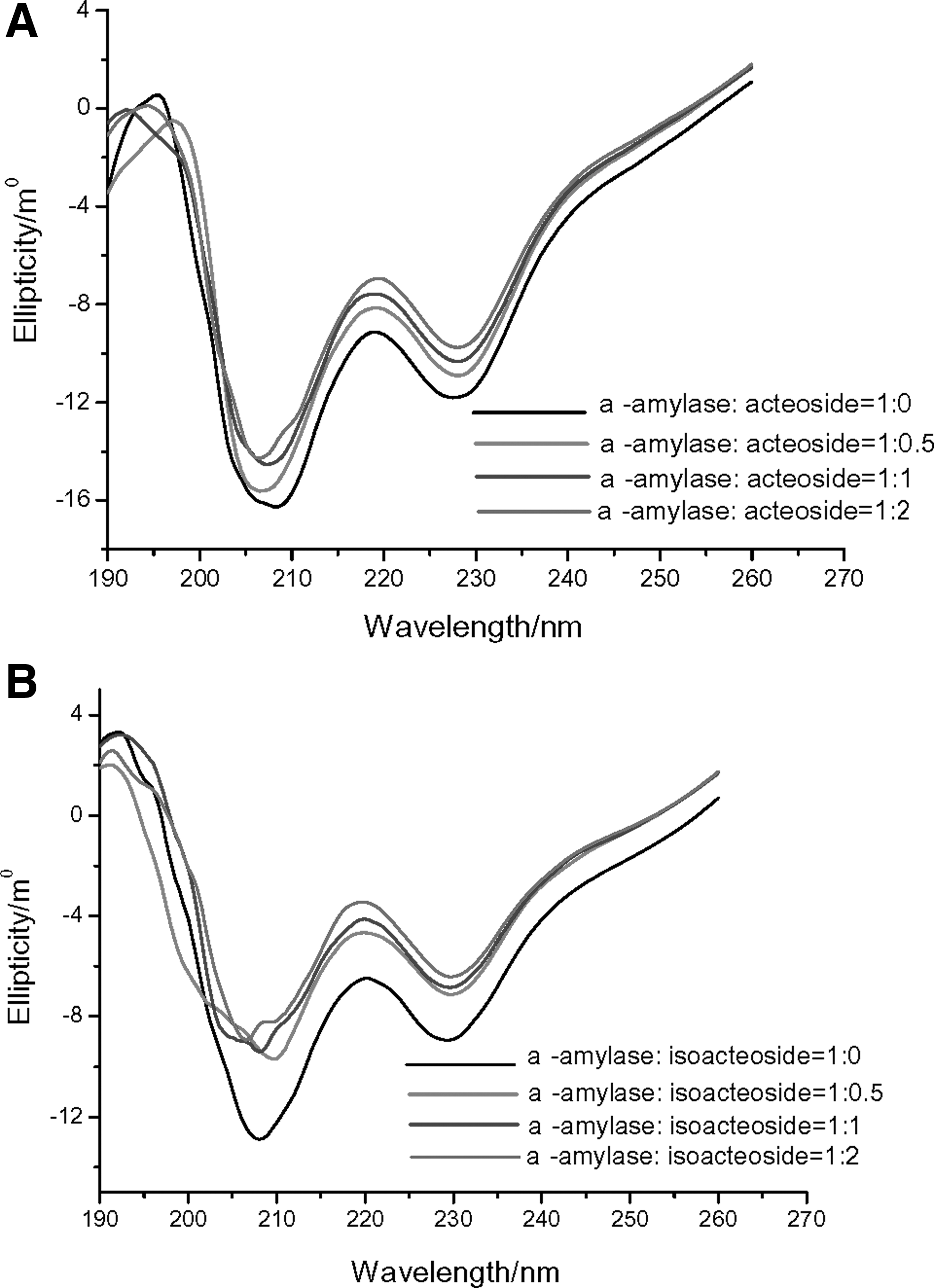
Circular dichroism assay of acteoside
Docking studies
Docking studies (Fig. 5) showed that the binding site for acteoside and isoacteoside on α-amylase differed. Acteoside was surrounded by Trp59, Gln63, Trp58, Asp300, Tyr62, Glu233, Asp197, Ala198, Leu162, Tyr151, Ile148, Gln 161, Val 163, and Leu165 in α-amylase, with possible hydrogen bonding between Asp197, Ieu148, and Glu233 (Fig. 5A). Isoacteoside was surrounded by Trp357, Pro54, Asn53, Val51, Asp300, Asp197, Arg 195, Tyr62, His305, Typ58, Gln63, Trp59, and Asp356, with possible hydrogen bonding between Asn53, Asp197, and Trp59 (Fig. 5B). The catalytic sites of porcine pancreatic α-amylase are Glu233, Asp300, and Asp197. 28 The hydrogen bonding distance between Asp197 for catalytic sites in α-amylase with acteoside and isoacteoside was 5.19 and 8.25 Å2, respectively. The binding sites for both acteoside and isoacteoside were close to the active site pocket of α-amylase, but closer for acteoside than isoacteoside.

Best-docked conformations of α-amylase–acteoside and α-amylase–isoacteoside interactions. The amino acid residues thought to interact with acteoside and iosacteoside are shown as a 2D representation by use of LigX in MOE. Acteoside
Discussion
Acteoside and isoacteoside are phenolic compounds, containing galloyl and hydroxyl groups. Phenolic compounds have a strong ability to interact with enzymes, which may be achieved by hydrophobic interactions in nature and stabilized by hydrogen bonds. 29,30 Our fluorescence spectroscopy results indicated that the binding of acteoside and isoacteoside to α-amylase was driven mainly by hydrophobic interactions of interior hydrophobic groups of α-amylase with acteoside and isoacteoside. Then, the interactions stabilized by the formation of hydrogen bonds between the polar groups at the surface of the α-amylase and OH groups of acteoside and isoacteoside. The docking studies further revealed this possibility.
There are at least two possibilities for the activity change of α-amylase. First, the change in protein structure may affect the catalytic activity of this enzyme. In this study, fluorescence and CD analyses demonstrated that the binding of acteoside and isoacteoside to α-amylase induced conformational changes of α-amylase, which resulted in changed enzyme activity. Second, the inhibitors' close proximity to the active site pocket of the enzyme may indirectly inhibit the enzyme activity. The competitive inhibition of α-amylase by acteoside and isoacteoside indicated that acteoside and isoacteoside might bind the catalytic sites of α-amylase. Some phenolic compounds, such as epigallocatechin gallate, luteolin, myricetin, apigenin, and daidzein, inhibit α-amylase. 31 –35 Computational ligand docking revealed that the inhibitory activity of phenolic compounds on α-amylase depends on hydrogen bonds between the catalytic residues of the binding site and the hydroxyl groups of phenolic compounds. 33,34 Our results seemed to agree with this finding. Therefore, acteoside and isoacteoside binding near the active site pocket of α-amylase may induce conformational changes in α-amylase, which changed its activity.
We showed that acteoside from small-leaved kudingcha strongly inhibited α-amylase activity in vitro, with preheated acteoside showing reduced inhibition because the preheated acteoside we used contained some isoacteoside. Acteoside was more effective than isoacteoside for inhibiting α-amylase activity. This phenomenon may be due to the following. First, according to the result of binding constants and the number of binding sites per protein n (Table 1), the affinity interaction was greater between acteoside and α-amylase than isoacteoside and α-amylase. Furthermore, docking studies indicated that the binding site for acteoside is closer to the active site pocket of α-amylase than the binding site of isoacteoside. Isoacteoside has a similar structure as acteoside, except that caffeoyl connects glucoyl carbon 4 and acteoside connects glucoyl carbon 6 (Fig. 6). Previous studies found that the interaction between α-amylase and phenolic compounds depends on the structures of α-amylase and phenolic compound. 35,36 The different affinities and binding sites of acteoside and isoacteoside resulted in different inhibition rates, which may be due to the structural difference between acteoside and isoacteoside. The detailed mechanism is worthy of further study.

Chemical structure of acteoside
Although various phenolic compounds that inhibit enzymes have been reported, 37 we showed for the first time a difference between acteoside and acyl-migrated acteoside inhibition of α-amylase. Small-leaved kudingcha is known as a healthy tea; however, the acyl migration of acteoside should be considered when the tea is heated.
In conclusion, we compared the inhibition of α-amylase by acyl-migrated acteoside and acteoside. Inhibition of α-amylase delays the digestion of carbohydrates and can reduce the rate of glucose absorption, so it may have importance in diabetes and obesity. Acteoside treated at high temperatures resulted in acyl migration of acteoside to isoacteoside. Acteoside was more effective than isoacteoside for inhibiting α-amylase activity. Acteoside and isoacteoside binding to α-amylase may induce conformational changes of α-amylase, and the binding site of acteoside and isoacteoside being near the active site pocket of α-amylase may explain the changed activity in α-amylase. The different inhibition rates of acteoside and isoacteoside may be due to their structural differences.
Footnotes
Acknowledgments
This study was supported, in part, by research funding from the Natural Science Funding of Guangdong Province (no. 2015A030313558), the Technological Research Funding of Guangdong Province (no. 2015A010107017), and the Research Funding of Shenzhen (nos. JCYJ20140418091413497 and XZZ20150529165110750) to X.W.
Author Disclosure Statement
No competing financial interests exist.