
Abstract
Ilex cornuta, commonly known as Chinese holly, is an evergreen shrub from the family Aquifoliaceae, and it is widely distributed in Korea and China. In folk medicine, the leaves of I. cornuta are used for treatment of several disorders, including weakness of the waist and knees, arthrodynia, headache, acute conjunctivitis, toothache, urticaria, rheumatic arthralgia, and cardiovascular diseases. In this study, we investigated the anti-inflammatory effects of an I. cornuta leaf ethanol extract (ILE) and its underlying mechanisms of action. The anti-inflammatory activities of ILE were evaluated in murine RAW 264.7 macrophages, using lipopolysaccharide (LPS) stimulation. ILE treatment-related changes in the production of nitric oxide (NO), prostaglandin E2 (PGE2), and proinflammatory cytokines were also measured. Finally, the expression of signaling molecules involved in inflammatory reactions was also assessed. Pretreatment of macrophages with ILE attenuated the expression of inducible NO synthase and cyclooxygenase-2, resulting in a decrease in NO and PGE2 production. The secretion of proinflammatory cytokines such as interleukin (IL)-6 and IL-1β was also reduced. Furthermore, ILE reduced extracellular signal-regulated kinases (ERK) phosphorylation, without affecting the inhibitor of kappa Bα and other mitogen-activated protein kinases. Liquid chromatography-tandem mass spectrometry analysis (LC-MS/MS) demonstrated that 1 g of ILE contains 27 mg of kaempferol, 0.3 mg of vanillic acid, and 21 mg of combined amount of isoquercetin and hyperin, among which isoquercetin and kaempferol significantly suppressed IL-6, IL-1β, and PGE2 production. Our results demonstrated that ILE possesses anti-inflammatory effects mediated through inhibition of ERK phosphorylation.
Introduction
I
Inflammation plays a critical role in host defense against external irritants, pathogens, and cell injury. It eliminates the cause of cell injury, removes necrotic cells and tissues, and initiates tissue repair. 9 Although the main purpose of inflammation is to protect the host from harmful stimuli, chronic inflammation without appropriate resolution causes inflammatory diseases, including inflammatory bowel disease and rheumatism. 10,11 Moreover, it has been reported that inflammation is involved in all stages of cardiovascular diseases, which may develop into life-threatening conditions. 12,13
Macrophages are one of the most efficient phagocytes in the immune system. They engulf and digest pathogens or cellular debris and present antigens to naive lymphocytes for their activation and maturation. 14 For pathogen recognition, macrophages express pattern recognition receptors (PRRs), which recognize pathogen-associated molecular patterns present on diverse pathogens. Among the various types of PRRs, the toll-like receptor 4 recognizes lipopolysaccharide (LPS), which forms part of the outer membrane of gram-negative bacteria. 15 Upon LPS stimulation, macrophages are activated, which produce inflammatory mediators such as nitric oxide (NO) and prostaglandin E2 (PGE2), via the expression of inducible NO synthase (iNOS) and cyclooxygenase-2 (COX-2). 16,17 In addition, activated macrophages secrete proinflammatory cytokines, such as interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α, which stimulate leukocytes, endothelial cells, and macrophages to continue the inflammatory response. 18,19
Although the leaves of I. cornuta have been widely used in China and Korea as a folk medicine for a long time, their anti-inflammatory activity has not yet been experimentally evaluated. Therefore, we assessed the anti-inflammatory activity of the I. cornuta leaf ethanol extract (ILE) on LPS-stimulated murine macrophages and explored its underlying mechanisms of action.
Materials and Methods
Cell culture
RAW 264.7 cells, a murine macrophage cell line, were cultured in Dulbecco's modified Eagle's medium (Welgene, Gyeongsan, Korea) supplemented with fetal bovine serum (Welgene), 100 U/mL penicillin, and 100 μg/mL streptomycin (Gibco, Grand Island, NY, USA). This was performed at 37°C, in humidified air with 5% CO2.
Ethanol extract of the I. cornuta leaf and compounds
A 95% ethanol extract of the I. cornuta leaf (Code No.: FBM192-002) was obtained from the International Biological Material Research Center (Daejeon, Korea). The extract was dissolved in dimethyl sulfoxide (DMSO; Duchefa Biochemie, Haarlem, The Netherlands) to prepare a 100 mg/mL stock solution. Quercetin and protocatechuic acid were purchased from Tokyo Chemical Industry (Tokyo, Japan). Isoquercetin, hyperin, isorhamnetin, vanillic acid, formononetin, and kaempferol were purchased from Sigma-Aldrich (St Louis, MO, USA). The final concentration of DMSO in the culture medium did not exceed 0.5% in volume-to-volume ratio.
Compound quantification
The contents of hyperin, isoquercetin, kaempferol, and vanillic acid in ILE were quantified using an API 4000 LC-MS/MS system (AB SCIEX, Framingham, MA, USA) equipped with an electrospray ionization interface. The compounds were separated on a reversed-phase column (Poroshell 120 EC-C18, 50 × 2.1 mm internal diameter, 2.7 μm particle size; Agilent, Wilmington, DE, USA) in the mobile phase (0.1% formic acid aqueous solution and acetonitrile, 7:3, v/v). The column was heated to 30°C, and the mobile phase was eluted at 0.2 mL/min using an HP 1260 series pump (Agilent). The turbo ion spray interface was operated at 4500 V and 500°C. Hyperin, isoquercetin, vanillic acid, and diclofenac (internal standard, IS), except kaempferol ([M+H]+), were mainly identified as deprotonated ions [M-H]−. The ion transition was at m/z 462.9 → 299.7 for hyperin and isoquercetin, 167.1 → 107.9 for vanillic acid, 296.1 → 251.7 for the IS, and 289.0 → 210.9 for kaempferol. Quantification was performed by selective reaction monitoring of the precursor ions and the related product ions using the ratio of the area under the peak for each solution. The data were processed using Analyst software (version 1.5.2; Applied Biosystems, Foster City, CA, USA). 20
The compounds were individually dissolved in ethanol and serially diluted to obtain final concentrations of 10 ng/mL through 10 μg/mL. Using linear regression, calibration graphs were derived from the ratio between the area under the peak of each compound and the IS (100 ng/mL). ILE was also serially diluted with ethanol and injected onto the liquid chromatography-tandem mass spectrometry (LC-MS/MS) system to determine the compounds.
Cell viability assay
RAW 264.7 cells were plated at a density of 5 × 104 cells/100 μL per well in a 96-well plate. Following pretreatment with 0–400 μg/mL concentrations of ILE for 4 h, cells were rinsed with ice-cold phosphate-buffered saline (PBS) and stimulated with 1 μg/mL of LPS (Sigma-Aldrich). After 20 h, 10 μL of 5 mg/mL 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich) was added. Cells were incubated for 5 h, and 100 μL of 0.04 N HCl in isopropanol was added to dissolve the formazan crystals. To quantify the viable cells, the optical density was measured at 540 nm, using an EMax microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Measurement of NO production
RAW 264.7 cells were treated with ILE or compound for 4 h and rinsed with ice-cold PBS. Subsequently, cells were stimulated with 1 μg/mL of LPS. After 20 h, the culture medium was collected, and 50 μL of the supernatant was mixed with 50 μL of Griess reagent (Sigma-Aldrich). The optical density was measured at 540 nm, and NO production was quantified using sodium nitrite (NaNO2; Junsei Chemical, Co., Chuo-ku, Tokyo, Japan) to generate a nitrite standard reference curve.
Enzyme-linked immunosorbent assay
RAW 264.7 cells were treated with ILE or compound and LPS, and the cell culture medium was harvested to measure the secretion of IL-6, IL-1β, TNF-α, and PGE2. Secreted cytokine levels were measured using the sandwich enzyme-linked immunosorbent assay (ELISA) method according to the procedure of Im et al. 21 Purified anti-mouse IL-6, TNF-α antibodies, biotinylated anti-mouse IL-1β, and TNF-α antibodies were purchased from eBioscience (San Diego, CA, USA). Purified anti-mouse IL-1β antibody and biotinylated anti-mouse IL-6 antibody were purchased from BD Biosciences (San Diego, CA, USA). Each secreted cytokine level was quantified using a standard reference curve generated with recombinant murine IL-6, IL-1β, and TNF-α (Peprotech, Rocky Hill, NJ, USA).
The production of PGE2 in LPS-stimulated RAW 264.7 cells was measured using a PGE2 ELISA kit (Cayman Chemical, Ann Arbor, MI, USA), following the manufacturer's instructions.
Quantitative real-time PCR
RAW 264.7 cells were treated with ILE or compound and LPS and lysed in RNAiso (TAKARA BIO, INC., Otsu, Shiga, Japan). Total RNA was extracted and reverse transcribed into complementary DNA using M-MLV Reverse transcriptase (Promega, Madison, WI, USA). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using TOPreal™ qPCR 2X PreMIX (Enzynomics, Daejeon, Korea), following the manufacturer's instructions, and a CFX Connect™ Real-Time PCR Detection System (Bio-Rad Laboratories, San Francisco, CA, USA). The sequences of primers used in this study are listed in Table 1.
COX-2, cyclooxygenase-2; iNOS, inducible nitric oxide synthase.
Western blotting
After pretreatment with ILE, RAW 264.7 cells were stimulated with LPS for 15 min. The cells were then lysed in a RIPA Lysis and Extraction Buffer (Thermo Fisher Scientific, Rockford, IL, USA), and further experiments were performed as previously described. 22,23 Each membrane was incubated with an appropriate anti-mouse primary antibody (Cell Signaling, Danvers, MA, USA). Horseradish peroxidase-conjugated goat anti-rabbit IgG antibody (Bio-Rad) was used as a secondary antibody. A ChemiDoc XRS densitometer and Quantity One software (Bio-Rad) were used to detect and quantify the protein bands.
Statistical analyses
All data were presented as mean ± standard deviation. Experiments were repeated three to five times, and representative data were selected. The difference between the experiments was assessed using a two-tailed Student's t-test, and P < .05 was considered statistically significant. All statistical analyses were performed using Prism 5.0 (GraphPad Software, San Diego, CA, USA).
Results
Cytotoxic effects of ILE
To evaluate the cytotoxic effects of ILE on RAW 264.7 cells, an MTT assay was performed. As shown in Figure 1, ILE did not show any cytotoxicity at a concentration of ≤200 μg/mL; however, at 400 μg/mL, cell viability was decreased significantly compared with the control. Therefore, the ILE concentration used in the following experiments was ≤200 μg/mL to avoid a cytotoxic effect.
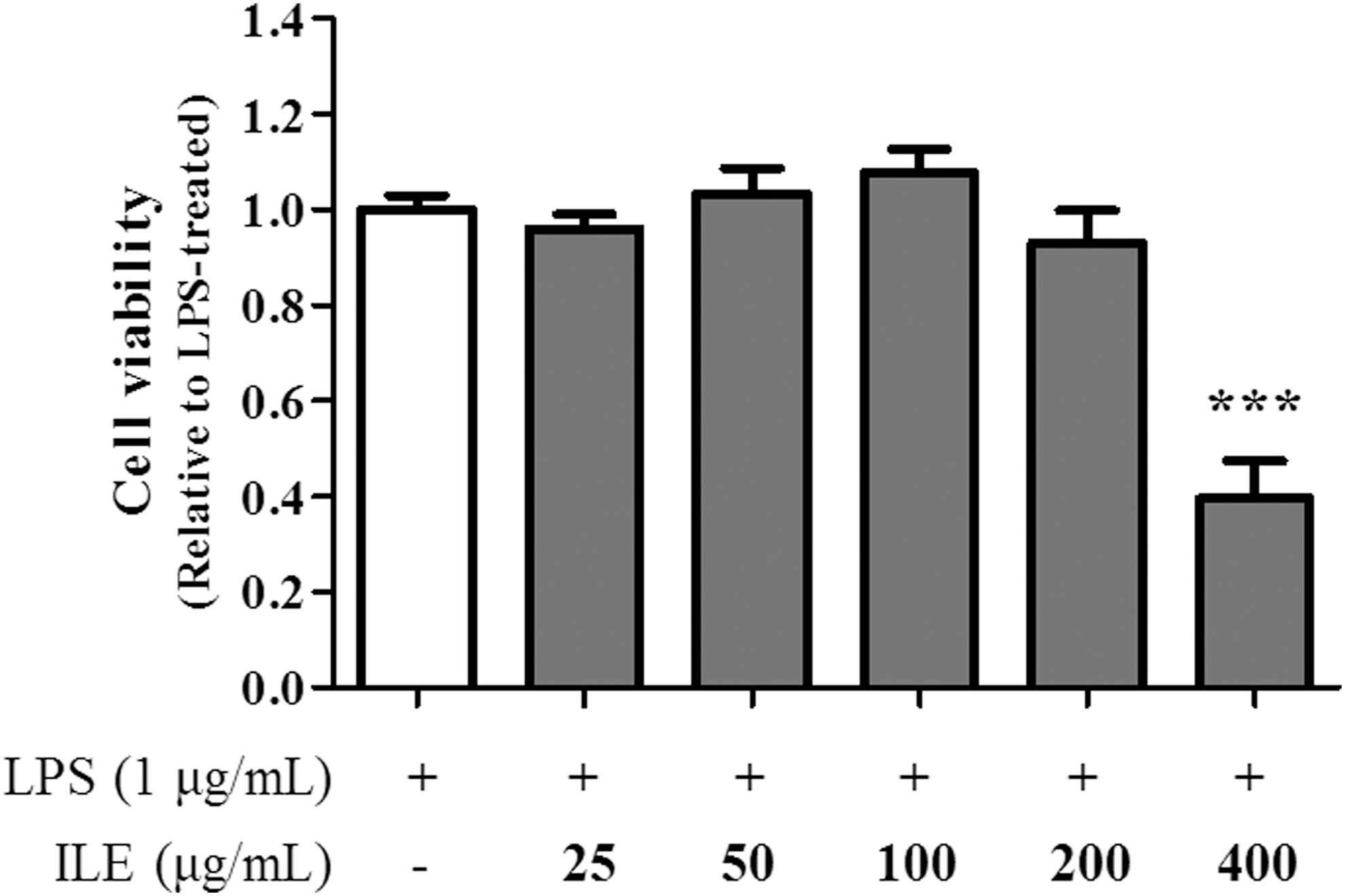
The effect of ILE on cell viability. RAW 264.7 cells were treated with ILE (0, 25, 50, 100, 200, and 400 μg/mL) for 4 hours, and activated using LPS (1 μg/mL) for 20 hours. Then cell viability was measured using an MTT assay. Cell viability was represented as fold change compared to the LPS-treated control. The data are representative of five independent experiments with similar results, and they are expressed as mean ± S.D. ***p < .001.
Inhibition of inflammatory mediators
To determine the anti-inflammatory effects of ILE, the expression of iNOS messenger RNA (mRNA) and the quantity of NO production were measured using qRT-PCR and the Griess reaction, respectively. As shown in Figure 2A and B, ILE suppressed the expression of iNOS mRNA, and this was accompanied by a decrease in NO production in a dose-dependent manner.

ILE suppressed NO and PGE2 production by inhibiting the expression of iNOS and COX-2 mRNA. RAW 264.7 cells were treated with ILE (0, 25, 50, 100, and 200 μg/mL) for 4 hours, and activated using LPS (1 μg/mL) for 20 hours. Total RNAs were extracted from the cells, and qRT-PCR was conducted to quantify
ILE treatment reduced the expression of COX-2 mRNA in LPS-stimulated RAW 264.7 cells in a dose-dependent manner (Fig. 2C). In accordance with the decrease in COX-2 expression, PGE2 production was also downregulated by ILE (Fig. 2D).
Inhibition of proinflammatory cytokine production
LPS induces the production of proinflammatory cytokines, such as IL-6, IL-1β, and TNF-α, in macrophages. To determine the anti-inflammatory effect resulting from ILE-mediated suppression of proinflammatory cytokine production, secreted cytokine levels were measured using ELISA. As shown in Figure 3, LPS treatment greatly increased the secretion of IL-6, IL-1β, and TNF-α in control macrophages. However, above a concentration of 50 μg/mL, ILE treatment selectively inhibited the production of IL-6 and IL-1β in a dose-dependent manner (Fig. 3A, B), without affecting TNF-α production (Fig. 3C).
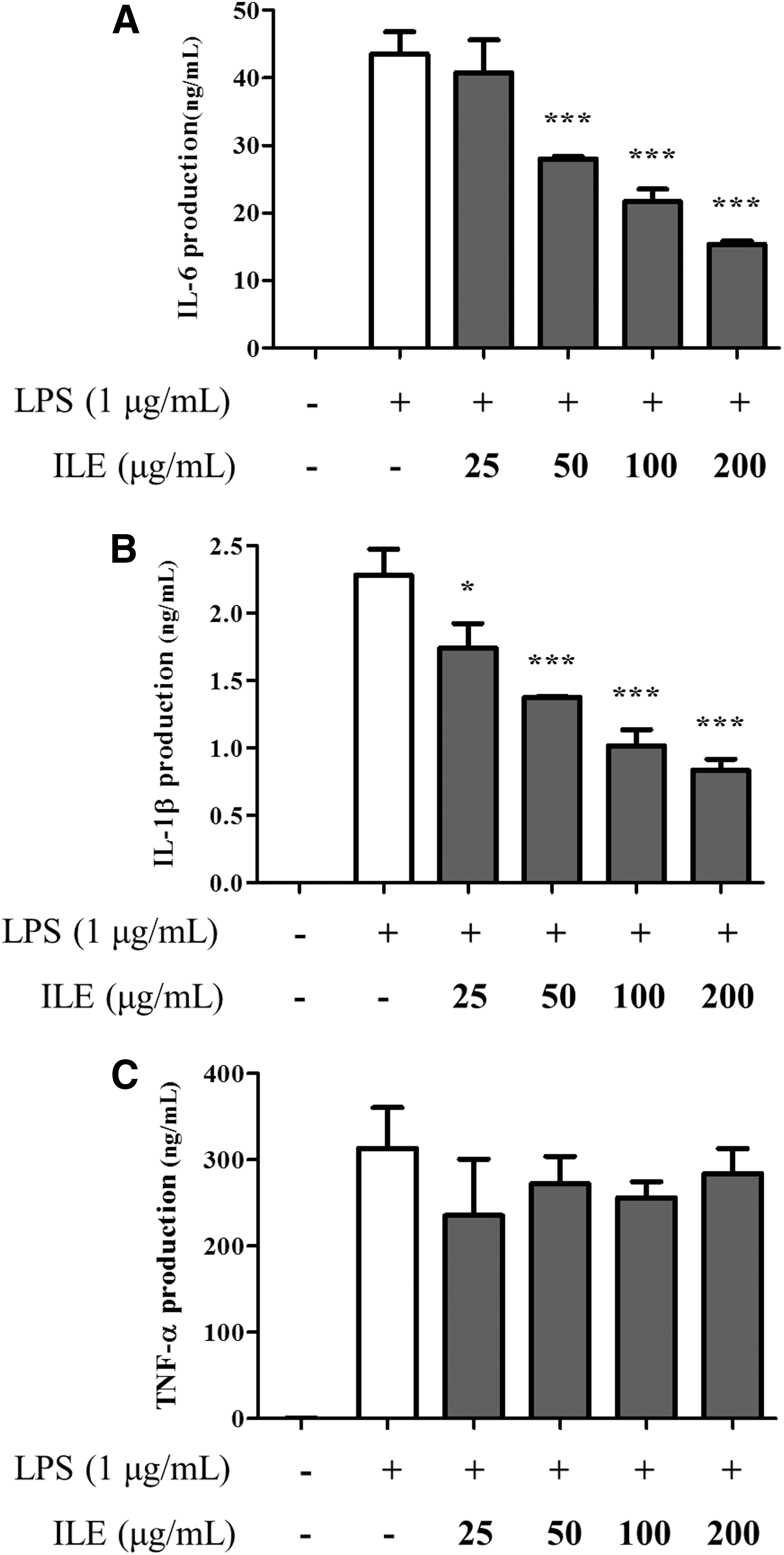
The inhibitory effect of ILE on IL-6 and IL-1β production. Cells were treated with ILE (0, 25, 50, 100, and 200 μg/mL) for 4 hours, followed by LPS (1 μg/mL) activation for 20 hours. The production of pro-inflammatory cytokines such as
Inhibitory effect of ILE on extracellular signal-regulated kinases pathway
The mitogen-activated protein kinase (MAPK) pathway consists of the extracellular signal-regulated kinases (ERK), c-Jun N-terminal kinases (JNK), and p38 pathways, and hence, the LPS-induced phosphorylation of ERK, JNK, and p38 was tested in RAW 264.7 cells. Our data demonstrate that ILE treatment decreased the ratio of phosphorylated ERK to ERK at 200 μg/mL by 27% (Fig. 4A, B). However, phosphorylation of JNK and p38 were not affected by ILE treatment (Fig. 4A, B). Upon LPS stimulation, inhibitor of kappa Bα (IκBα) is phosphorylated and subsequently degraded, allowing translocation of nuclear factor-kappa B (NF-κB) into the nucleus. 24,25 As shown in Figure 4C and D, ILE treatment did not affect the phosphorylation of IκBα, suggesting that NF-κB signaling is not a target for ILE. These results suggest that the anti-inflammatory effects of ILE are mediated by selective suppression of the ERK pathway.

ILE selectively inhibited the phosphorylation of ERK. Cells were treated with ILE (0, 25, 50, 100, and 200 μg/mL) for 4 hours, followed by LPS (1 μg/mL) for 15 minutes. Total cell lysates were used to perform immunoblotting analysis, and the expression levels of p-ERK, ERK, p-p38, p38, p-JNK, JNK
Anti-inflammatory effects of ILE compounds
Previous studies have reported that the leaf of I. cornuta contains formononetin, hyperin, isorhamnetin, isoquercetin, kaempferol, protocatechuic acid, quercetin, and vanillic acid. 26,27 To identify the active compounds responsible for the anti-inflammatory properties of ILE, the eight compounds were individually tested for their cytotoxic and anti-inflammatory effects. As shown in Figure 5A, hyperin, isoquercetin, kaempferol, and vanillic acid did not exert significant cytotoxicity, as cell viability remained ≥80% at concentrations ranging from 25 to 100 μM; the other four compounds showed cytotoxic effects. Among the four noncytotoxic compounds, isoquercetin, kaempferol, and vanillic acid reduced NO production in a dose-dependent manner (Fig. 5B).
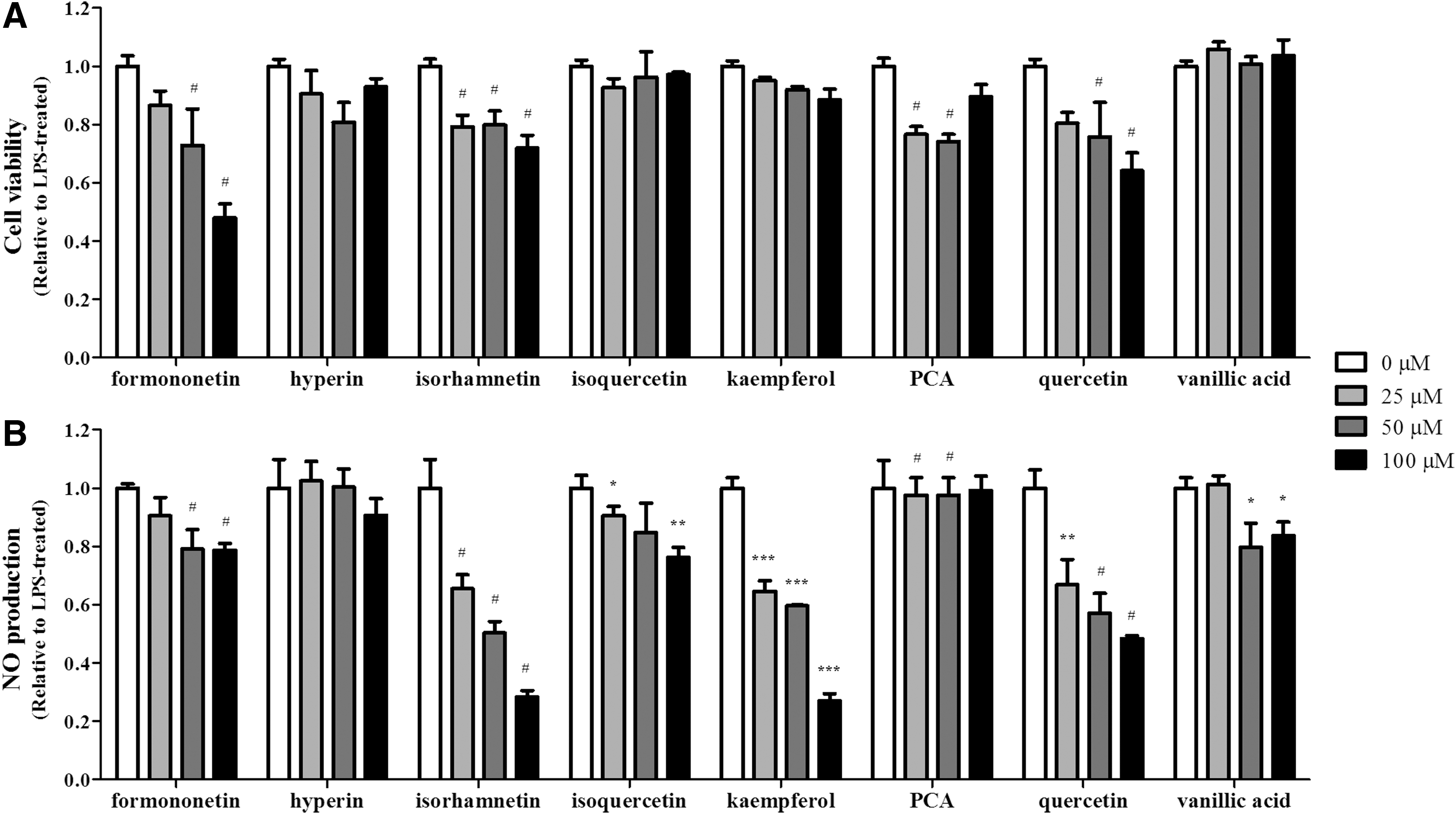
The effect of single compounds present in ILE on cell viability and NO production. RAW 264.7 cells were treated with each compound (0, 25, 50, and 100 μM) for 4 hours. Cells were then activated with LPS (1 μg/mL) for 20 hours.
We then assessed if the four noncytotoxic compounds would regulate the production of proinflammatory cytokines and PGE2, using ELISAs. The data demonstrate that isoquercetin and kaempferol significantly decreased IL-6, IL-1β, and PGE2 production (Fig. 6A, B, D). In contrast, hyperin only reduced IL-1β production, while vanillic acid did not possess any inhibitory effects on cytokine and PGE2 production (Fig. 6A, B, D). None of the compounds was found to suppress the production of TNF-α (Fig. 6C).
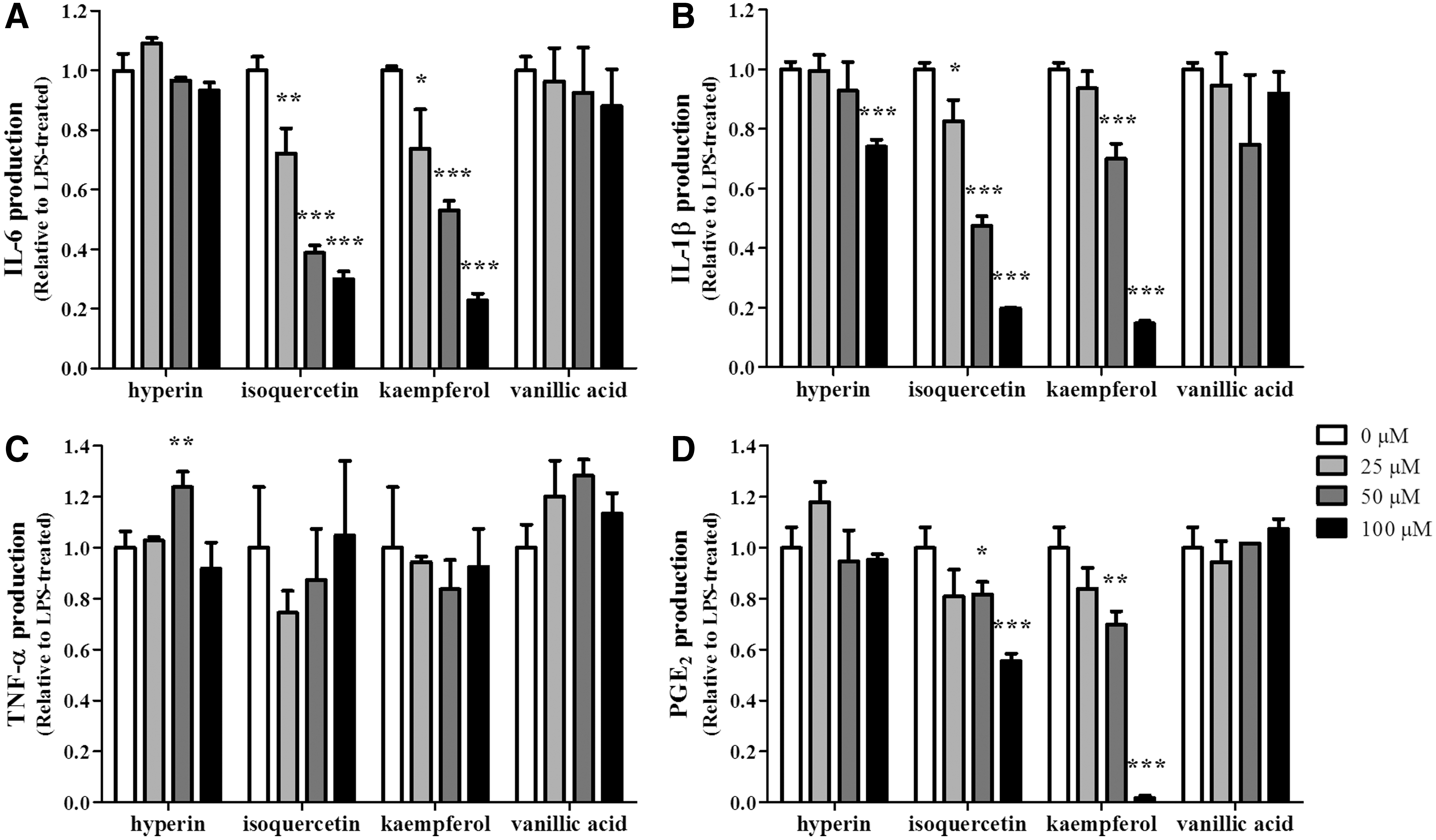
Isoquercetin and kaempferol play a pivotal role in the anti-inflammatory effect of ILE. Cells were treated with each compound (0, 25, 50, 100 μM) for 4 hours, followed by LPS (1 μg/mL) activation for 20 hours. The production of pro-inflammatory cytokines, such as
Quantification of the compounds present in ILE
To determine the quantity of hyperin, isoquercetin, kaempferol, and vanillic acid present in ILE, LC-MS/MS analysis was performed. One gram of ILE contains 27 mg of kaempferol, 0.3 mg of vanillic acid, and 21 mg of isoquercetin and hyperin (data not shown). Since isoquercetin and hyperin are epimers, they could not be separated by LC-MS/MS, so the value of 21 mg combines the amounts of isoquercetin and hyperin.
Discussion
Despite its use in treating arthrodynia, rheumatic arthralgia, and rheumatic arthritis in folk medicine, there is a paucity of literature experimentally validating the pharmacological properties of I. cornuta. In this study, we show that the ILE exhibits anti-inflammatory effects in murine macrophages.
It is well known that macrophages play a key role from the initiation to the end stage in diseases caused by chronic and relapsing inflammation. 28 Upon LPS stimulation, macrophages activate the NF-κB and MAPK pathways to provoke inflammatory reactions via inflammatory mediators such as NO, PGE2, and proinflammatory cytokines. 29 The MAPK pathways are major players in the inflammatory responses of macrophages. 30 Activated MAPKs are phosphorylated and translocated into the nucleus to induce the transcription of target genes such as proinflammatory cytokines, iNOS, and COX-2. 31,32
Our data demonstrated that ILE selectively reduced the activation of ERK, without affecting the JNK and p38 pathways. The inflammation-modulating properties of ERK have been well investigated, and studies have reported that ERK regulation of TNF-α production is highly dependent on the origin of the macrophages. Means et al. have shown that the ERK inhibitor PD98059 inhibited expression of the LPS-induced TNF-α gene in a murine cell line derived from alveolar macrophages, but not in a nonpulmonary macrophage cell line. 33 As RAW 264.7 cells originate from a murine ascites cell line, the inability of ILE to inhibit TNF-α production (despite the decrease in ERK phosphorylation) might relate to this cell type-dependent differential regulation of TNF-α. Furthermore, through an additional experiment using specific inhibitors of MAPK-ERK kinase (MEK), PD98059 and U0126, we observed that simultaneous inhibition of MEK1/2 by U0126 blocked TNF-α production, while selective inhibition of MEK1 by PD98059 did not in RAW 264.7 cells (data not shown). In accordance with our results, the failure of PD98059 to inhibit TNF-α release in RAW 264.7 cells has been previously reported. 33,34 Thus, we assume that ILE selectively inhibited MEK1 without affecting MEK2 as PD98059 did, and as a result, ILE did not reduce LPS-induced TNF-α production. Another possible explanation for the unchanged level of TNF-α may relate to a report demonstrating that suppression of TNF-α production in RAW 264.7 cells required the simultaneous blockade of two MAPK pathways: ERK and p38. 34 Perhaps, since ILE only inhibited the ERK pathway and not the p38 pathway, suppression of TNF-α production could not be observed in our study.
I. cornuta leaves have been reported to contain eight compounds that are known for their anti-inflammatory properties. However, we found that only isoquercetin and kaempferol suppressed IL-6, IL-1β, and PGE2 production at a concentration that did not induce cytotoxicity. When quantified by LC-MS/MS, our data demonstrated that 200 μg/mL of ILE contained 18.87 μM kaempferol and <9.04 μM isoquercetin. Given that neither kaempferol nor isoquercetin individually exerted substantial anti-inflammatory effects, even at 25 μM, our data suggest that the anti-inflammatory effects of ILE might result from the synergistic effects of the multiple compounds constituting ILE. 35 –37 In fact, we observed that 20 μM of kaempferol combined with 10 μM isoquercetin exerted significant anti-inflammatory effects by reducing NO production, while 20 μM kaempferol or 10 μM isoquercetin alone showed weak or little effects on NO production, corroborating the synergistic effects of kaempferol and isoquercetin (data not shown).
Although phosphorylation of ERK by isoquercetin and kaempferol has not been tested in this report, previous studies have shown the reduced activation of signaling molecules by isoquercetin and kaempferol. Kim et al., have reported that kaempferol selectively suppressed ERK phosphorylation in LPS-induced RAW 264.7 cells. 38 Similarly, the inhibition of ERK as well as p38 and JNK was observed in rat hippocampus neurons, indicating an inhibitory role of isoquercetin in MAPK activation. 39 Future study would be required to elucidate the selective inhibition of ERK by isoquercetin in RAW 264.7 cells.
In summary, this study demonstrates the ability of ILE to inhibit inflammatory mediator production in activated macrophages. ILE reduced the production of NO, PGE2, and proinflammatory cytokines such as IL-6 and IL-1β, by downregulating the phosphorylation of ERK. These results suggest that I. cornuta could be a promising therapeutic agent for inflammatory diseases.
Footnotes
Acknowledgments
This work was supported by a grant from the Korean Society of Ginseng funded by Korea Ginseng Cooperation (2014) and by the World Class 300 Project R&D grant funded by the Korea Small and Medium Business Administration (SMBA).
Author Disclosure Statement
No competing financial interests exist.