
Abstract
Type 2 diabetes mellitus (T2DM) is a common and increasingly prevalent metabolic disorder, and effective preventive strategies against this disease are needed. The aim of the present study was to evaluate the potential antidiabetic properties of a dietary apple/kale extract (AKE), which was rich in phlorizin and flavonoids, in laboratory mice. Mice were fed a control diet, a Western-type high-sugar, high-fat diet (WTD), or a WTD plus AKE for 10 weeks. Body weight, food and energy intake, body composition, and blood glucose level were recorded in addition to the postprandial rise in blood glucose concentration after a single administration of glucose (oral glucose tolerance test, OGTT). Furthermore, changes in glucose-induced short-circuit current (ISC) in response to AKE and phlorizin administration were evaluated in situ in intestinal tissues with Ussing chambers. In addition, the in vitro inhibition of α-glucosidase by AKE was determined.
The present data suggest that supplementation of an AKE to a WTD significantly improved both blood glucose levels and OGTT in mice. Furthermore, in situ uptake of glucose was significantly inhibited by AKE. Finally, we showed that AKE significantly inhibits α-glucosidase activity in vitro.
We conclude that AKE exhibits antidiabetic properties by a dual mechanism, including the inhibition of α-glucosidase and sodium-dependent glucose transporter 1 (SGLT1). Thus, AKE has the potential to serve as a natural plant bioactive compound for dietary prevention strategies against T2DM.
Introduction
T
Glucose absorption is, in large part, accomplished by sodium-dependent glucose transporters (SGLTs) located in the mucosa of the small intestine (SGLT1). 5 SGLT1 expression is regulated by the diet, for example, sweet taste stimulation, 6 and is induced in patients with T2DM. 7
Phlorizin (phloretin-2′-o-glucoside), a dihydrochalcone present in apples and processed apple products, 8 has been shown to be a specific and competitive inhibitor of SGLT1 5 and SGLT2, which is found in the proximal renal tubule. 9 SGLT1 inhibition by apple-derived bioactives may lead to reduced postprandial blood glucose levels in mice and humans. 10 Furthermore, phlorizin significantly improved the oral glucose tolerance test (OGTT) in control mice 11 and feeding a phlorizin-supplemented diet to diabetic mice significantly reduced blood glucose levels. 12 A phlorizin-enriched dried and powdered pomace of unripe apples was found to be a health-promoting natural product for the reduction of postprandial glycemia of patients with T2DM. 13 In addition, besides phlorizin, apple extract contains a large variety of polyphenols (e.g., procyanidin B1, B2, B5, epicatechin, quercetin, and chlorogenic acid), which are also supposed to exhibit bioactivity. 14 However, phlorizin shows poor oral bioavailability since phlorizin can be hydrolyzed in the intestine to phloretin by β-glucosidase. 15 Furthermore, it can be converted by the gut microbiota into simpler phenolic compounds. 9
Kale consumption may be associated with reduced risks of heart and other chronic diseases 16 and exhibits positive effects on diabetes outcomes. 17 Dietary treatment of diabetic rats with an aqueous kale leaf extract resulted in a dose-dependent decrease in blood glucose. 18 This antihyperglycemic activity has been associated with polyphenols, including flavonoids and tannins, 19 which may affect intestinal α-glucosidase activity, thereby reducing glycemia after starch-rich meals. 2 Kale contains also other bioactives, including hydroxybenzoic acid derivatives, hydroxycinnamic acid derivatives, and anthocyanins. 20 However, kale remains an “understudied” vegetable, 19 and little is known regarding its antidiabetic properties.
Therefore, the aim of the current study was to investigate the antidiabetic properties of a dietary apple plus kale extract (AKE) in laboratory mice fed a Western-type diet (high-sugar, high-fat, WTD), which may favor the development of T2DM. Body weight, food and energy intake, body composition, and blood glucose level were recorded in addition to the postprandial rise in blood glucose concentration after a single administration of glucose (OGTT). Furthermore, changes in the intestinal glucose-induced short-circuit current (ISC) in response to AKE or phlorizin administration were determined in situ in Ussing chambers. Finally, the in vitro inhibition of α-glucosidase by AKE, phlorizin, and quercetin was systematically evaluated.
Materials and Methods
Experimental animals and diets
Eight-week-old C57BL/6NRj mice were purchased from Janvier Labs (Le Genest-Saint-Isle, France). Mice had free access to tap water and the experimental diets throughout the experiment. Animals were housed in groups of five animals, for which reason we decided to use female mice. Makrolon cages were provided with bedding, including wood wool, in a conditioned room (temperature, 22 ± 2°C; relative humidity, 50–60%; 12-h light/dark cycle).
After 2 weeks of adaptation, mice were divided into three groups of 10 mice each with equal mean body weights (19.3 ± 1.33 g). Control mice were fed a purified semisynthetic diet (C; Ssniff E15000-04, Soest, Germany) with a low-fat content over 10 weeks. Experimental mice were fed either a purified semisynthetic, energy-dense, high-fat, and high-sugar Western-type diet (WTD; Ssniff E15721-34) or a WTD plus apple/kale extract (AKE; BioActive Food, Bad Segeberg, Germany) in a dose of 50 g/kg diet (Table 1). Representative chromatograms of AKE with respect to phlorizin and flavonoid analysis are summarized in Figure 1. The analyzed contents in AKE consisted of 48.3 ± 2.73 g/kg phlorizin (equals an average dietary intake over the trial of 5.94 ± 0.267 mg/mouse/day), 5.67 ± 0.188 g/kg quercetin (equals an average dietary intake over the trial of 0.70 ± 0.031 mg/mouse/day), 4.85 ± 0.382 g/kg quercetin-3-O-glucoside (equals an average dietary intake over the trial of 0.60 ± 0.027 mg/mouse/day), 1.43 ± 0.284 g/kg kaempferol (equals an average dietary intake over the trial of 0.18 ± 0.008 mg/mouse/day), and 5.76 ± 0.863 g/kg kaempferol-3-
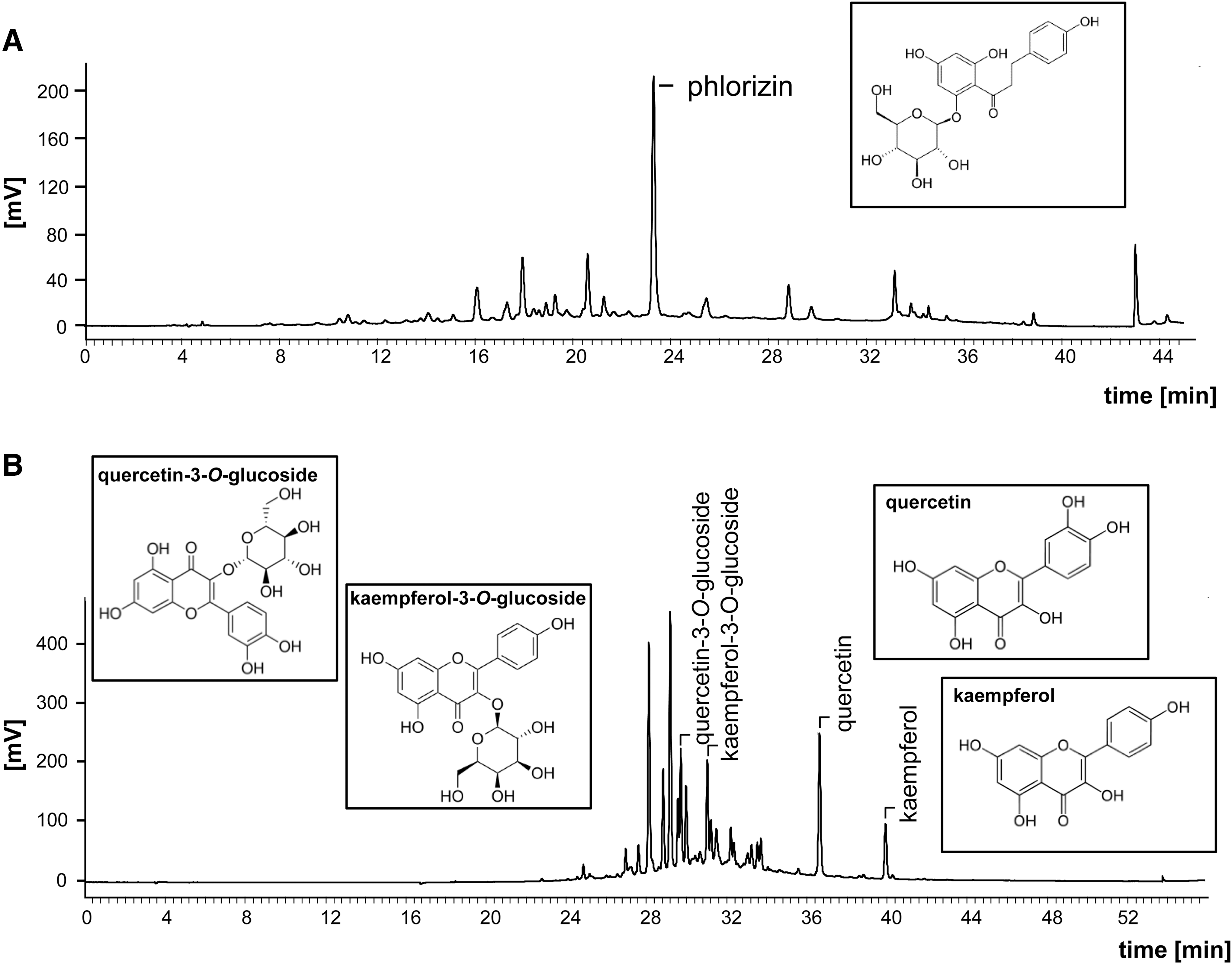
Representative HPLC chromatograms of phlorizin
AKE, apple/kale extract.
Food intake was recorded on a daily basis and body weight on a weekly basis. At the end of the trial, mice were fasted for 4 h before cervical dislocation, and blood was collected immediately after decapitation. Plasma was obtained by centrifugation and stored at −80°C until analysis.
High-performance liquid chromatography analysis of the AKE
The high-performance liquid chromatography-photodiode array (HPLC-PDA)-based analysis was performed using the method of Schieber et al. 21 Samples were extracted with either 70% (phlorizin) or 50% (flavonol) aqueous methanol. Analysis was conducted on an HPLC system (Jasco GmbH, Gross-Umstadt, Germany) equipped with a 250 mm × 4.6 mm Aqua 5 μ C-18 125 Å column (Phenomenex, Aschaffenburg, Germany). Two percent acetic acid in water (A) and acetonitrile (B) were utilized as the mobile phases for phlorizin detection. The following gradient conditions were used: 0 min—10% B, 28 min—38% B, 33 min—100% B, 38 min—100% B, 40 min—10% B, and 45 min—10% B. The flow rate was set at 0.8 mL/min and the detection wavelength was at λ 286 nm. The mobile phase for flavonol analyses consisted of (A) 0.01% aqueous trifluoroacetic acid (pH 2.9) and (B) 0.01% trifluoroacetic acid in acetonitrile. Gradient conditions were 0 min—0% B, 10 min—0% B, 40 min—50% B, 50 min—100% B, 51 min—0% B, and 56 min—0% B. The flow rate was set at 1 mL/min and the detection wavelength at λ 354 nm. Quantitation of phlorizin and flavonols was conducted by external calibration with ChromPass Chromatography Data System software (Version 1.8.6.1)
Blood glucose and OGGT
At week 9, five mice from each experimental group were fasted for 5–6 hours before the OGTT, with 2 g glucose/kg body weight orally administered by gavage. Blood samples were collected from the tail tip before and 15, 30, 60, 90, and 120 min after glucose administration. Blood glucose levels were determined by a glucometer (Abbott Freestyle Lite).
Further fasted blood samples were collected by puncture of the tail tip vein at weeks 2 and 7 and used for measuring glucose levels.
α-Glucosidase inhibition
α-Glucosidase inhibition measurements were conducted as described previously.
22
–24
In short, 50 μL of AKE, phlorizin, or quercetin was incubated for 5 min at 37°C with 50 μL of 0.5 U/mL α-glucosidase (from Saccharomyces cerevisiae; Sigma-Aldrich). Acarbose was used as a positive control. Subsequently, p-nitrophenyl-α-
Body composition
Body composition was determined using a time domain nuclear magnetic resonance technique after 8 weeks of the feeding trial using a MiniSpec (Bruker, BioSpin MRI GmbH, Ettlingen, Germany). Fat mass, lean mass, and free water were measured within 2 min in the living animal without anesthesia. The scan was performed using the following parameters: X-ray energy settings of 45 kVp and 177 μA, voxel size: 76 μm, integration time: 300 ms, 250 projections per 180°.
Ussing chamber experiments
C57BL/6J mice (age 4 months) were anesthetized with isoflurane before being decapitated. The distal ileum was removed, divided into four segments, cut longitudinally, and washed with ice-cold Krebs bicarbonate Ringer (KBR) solution (115 mmol/L NaCl, 2.4 mmol/L K2HPO4, 0.4 mmol/L KH2PO4, 1.2 mmol/L MgCl2, 1.2 mmol/L CaCl2, 25 mmol/L NaHCO3, pH 7.4). Serosa and the muscular layer were stripped off in KBR solution containing 5 μM indomethacin. The segments were mounted in the Ussing chamber with a 0.3 cm2 exposed surface area (EasyMount chamber system with P2300 chambers and P2304 sliders; Physiologic Instruments, San Diego, CA, USA). The Ussing chambers were filled with KBR solution containing 10 mmol/L mannitol apically and 10 mmol/L glucose basolaterally at 37°C and bubbled with 5% CO2/95% O2 (v/v). Tetrodotoxin (0.1 μM) was included in the serosal bath to minimize intrinsic residual neural activity. The transepithelial potential difference was continuously monitored under open-circuit conditions using a DVC 1000 amplifier (WPI) and recorded through Ag-AgCl electrodes and KBR agarose bridges. The ISC μA/cm was measured via an automatic VCC MC8 MultiChannel Voltage-Current Clamp (Physiologic Instruments) and stored using the Acquire & Analyze Data II acquisition software (Physiologic Instruments). The potential difference was allowed to stabilize for 20 to 30 min. Glucose (10 mM) was added apically, and 10 mM mannitol was given basolaterally. The glucose-stimulated ISC reached a stable plateau within 10 min and was then challenged by supplementing the mucosal bath with 100 μg/mL AKE or 25 μM phlorizin. The decline in the glucose-induced ISC was calculated.
Statistical analyses
Data are expressed as the mean ± SEM. Significant differences were calculated by testing for linear trends using one-way analysis of variance (ANOVA), followed by the Bonferroni test, or, in the case of inhomogeneity of variance, using Dunnett's multiple comparison post hoc test. The area under the curve (AUC) in the OGTT was determined with GraphPad Prism (Version 7.02). The data used for the IC50 determination were examined using a nonlinear regression sigmoid fit and was performed by GraphPad Prism. Results from Ussing chamber experiments were analyzed with a two-sided paired Students t-test; P-values less than 0.05 were considered significant. Unless stated otherwise, statistical analyses were performed with SPSS (Version 24.0).
Results
Body weight, food and energy intake, and body composition
Body weight significantly increased from the second to the 10th week in both WTD- and WTD plus AKE-fed groups compared to the control group (Fig. 2A). Food intake was significantly lower in the WTD group (P < .01) compared to the control group, whereas ANOVA revealed no significant differences between AKE and control mice in food intake over the entire feeding trial (Table 2). Control mice had a significantly lower energy intake than WTD- and AKE-fed mice (P < 0.01, Table 2).
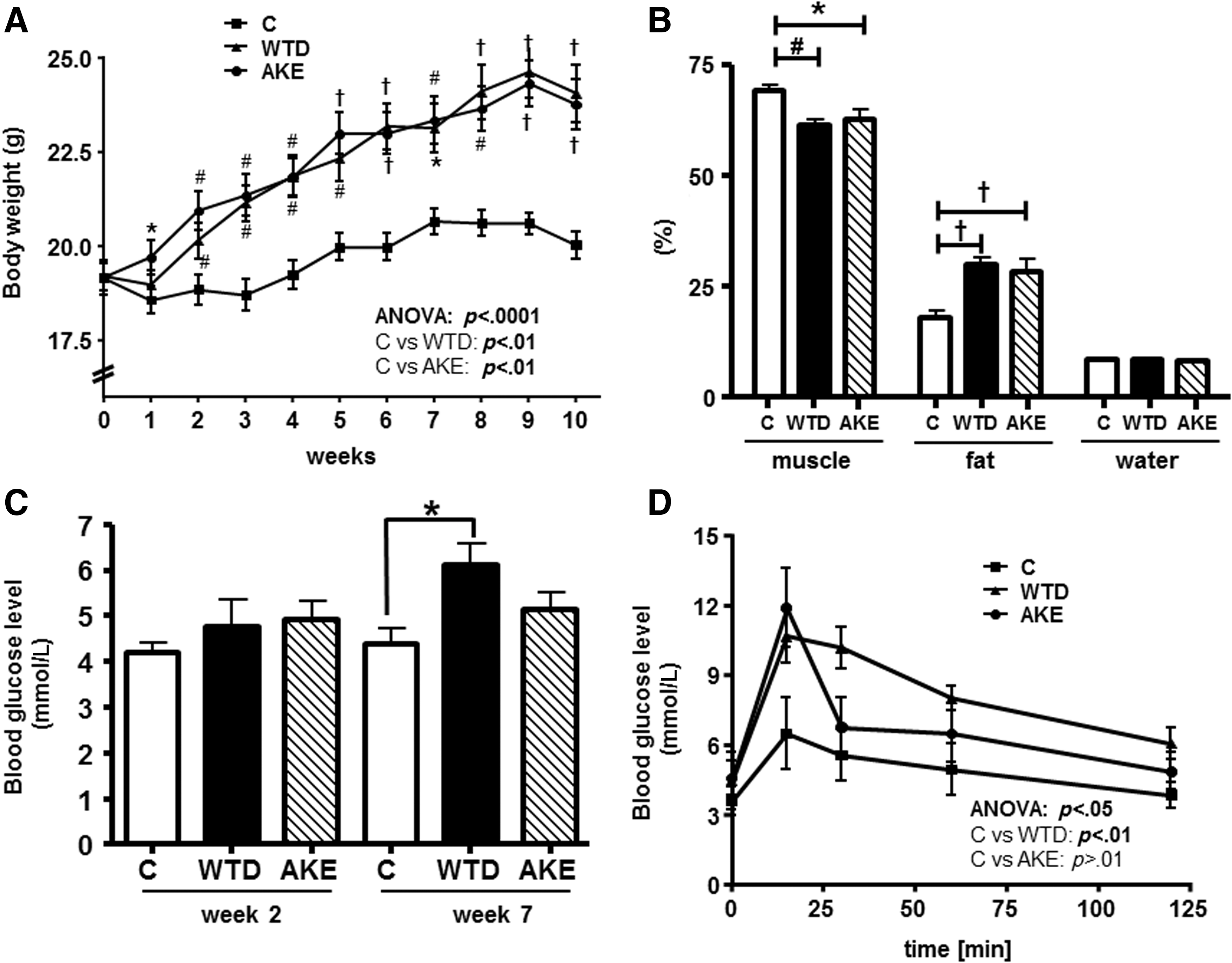
Body weight development
Data are presented as mean ± SEM in n = 10 mice/diet. Significant differences were calculated using ANOVA with repeated measures, followed by the Bonferroni test, or, in the case of inhomogeneity of variance, using Dunnett's multiple comparison post hoc test.
P < .05, # P < .01, † P < .001 compared to control within a week.
AKE, apple/kale extract; C, control diet; WTD, Western-type diet.
Percentages of muscle, fat, and water mass (absolute values related to body weight) in living animals obtained without using anesthesia are shown in Figure 2B. Our analysis revealed that control mice exhibited a significantly higher muscle and lower fat content compared to WTD- and AKE-fed mice with a similar water proportion. The AKE diet did not alter body composition compared to animals fed the WTD. Taken together, we showed that feeding a WTD plus AKE to mice did not significantly alter body weight, energy intake, and body composition compared to the mice fed the WTD only. Thus, the AKE was well tolerated by our mice.
Glucose levels and glucose response
Blood glucose levels of mice fed the different diets for 2 and 7 weeks are depicted in Figure 2C. In week 2, feeding a WTD or WTD plus AKE did not significantly alter the blood glucose levels compared to control animals. However, in week 7, feeding a WTD led to a significantly higher blood glucose level compared to controls, whereas feeding the WTD plus AKE did not. Therefore, mice on the AKE diet displayed lower blood glucose levels compared to mice on the WTD only.
The postprandial rise in blood glucose concentration (OGTT) was determined in mice after feeding the different dietary regimens for 9 weeks. Blood glucose levels at 0, 15, 30, 60, and 120 min after a single oral glucose administration of 2 g glucose/kg body weight are given in Figure 2D. Mice fed the AKE diet showed the highest blood glucose rise after 15 min, with a subsequent rapid decline to the control level. The AUC calculation revealed for the control group a total area of 588.2 ± 96.8 mmol/L × min, for the WTD group 963.0 ± 68.8 mmol/L × min, and for the AKE group 802.3 ± 113.4 mmol/L × min. ANOVA did not show any significant differences between control animals and mice fed the AKE-supplemented diet. In mice fed the WTD, the concentration of postprandial blood glucose was significantly higher (P < .01) compared to control mice. These results demonstrate that dietary supplementation of an AKE to the WTD significantly improved both blood glucose levels and OGTT in mice.
Glucose uptake in situ using Ussing chamber
Next, we evaluated the effect of AKE (100 μg/mL) and phlorizin (25 μM) on the uptake of 10 mM glucose in the apical intestine of wild-type C57BL/6J mice using Ussing chambers in situ (Fig. 3). Adding AKE significantly lowered the glucose-induced ISC from 135.8 ± 10.3 μA/cm2 to 73.7 ± 8.25 μA/cm2. Likewise, glucose uptake, measured by the ISC, was significantly decreased by phlorizin, leading to an ISC of 48.3 ± 6.82 μA/cm2 compared to an ISC of 160.3 ± 9.18 μA/cm2 for glucose only. Thus, our data suggest that uptake of glucose in mouse intestine was significantly inhibited by AKE and phlorizin in situ.

Changes in the short-circuit current (ISC) of glucose by AKE and phlorizin in small intestinal tissues mounted in Ussing chamber. Responses of the ISC with time in the intestine of nontreated wild-type C57BL/6J mice to apical glucose (10 mM) application and following AKE (100 μg/mL,
Discussion
Phlorizin is a highly specific and competitive inhibitor of SGLT1. 5 Apple extracts and apple-derived polyphenols have been reported to diminish SGLT1-mediated glucose uptake both in vitro and in vivo. 10 Furthermore, extracts from Bauhinia megalandra leaves, containing significant amounts of phlorizin, inhibited the intestinal glucose absorption in rats in a concentration-dependent manner and additively to phlorizin. 25 However, phlorizin applied via the diet exhibits poor bioavailability as it may be hydrolyzed in the intestine to phloretin by β-glucosidase. 15 Moreover, phlorizin can be converted by the gut microbiota into simpler phenolic compounds 9 ; therefore, phlorizin has been administered intraperitoneally in previous studies. 26
Kale contains polyphenolic compounds, including flavonoids and tannins. 19 Quercetin is one of the major flavonoids in kale. 27 The proportion of 50 g AKE/kg diet (Table 1) leads to a final concentration of ∼0.2% phlorizin (equals an average dietary phlorizin intake over the trial of 5.94 mg/mouse/day), 0.05% quercetin (equals an average dietary quercetin intake over the trial of 1.3 mg/mouse/day), and 0.03% kaempferol (equals an average dietary kaempferol intake over the trial of 0.89 mg/mouse/day) in the AKE-enriched diet. Previously, Rodriguez et al. 28 identified quercetin and kaempferol as compounds that reduce glucose absorption in isolated intestinal segments in situ. Most importantly, quercetin and kaempferol exerted an additive inhibitory effect on glucose absorption when combined with phlorizin. We therefore speculate that in contrast to feeding an apple extract only, the combination of apple plus kale may exhibit superior antidiabetic properties. In fact, the present results suggest that dietary supplementation of a WTD with AKE significantly improved both blood glucose levels and OGTT in mice. In addition, AKE as well as phlorizin alone significantly reduced glucose uptake in our Ussing chamber experiments in situ, which was shown by a reduced glucose-induced ISC in response to AKE and phlorizin administration. Unfortunately, in our mouse study, phlorizin concentration in the plasma could not be determined due to the limited sample volume.
In addition to inhibition of SGLT1, α-glucosidase, a key enzyme involved in the intestinal digestion of oligosaccharides/disaccharides into monosaccharides, 29 may be another target of AKE as far as its glucose-lowering activity is concerned. Currently, there are antidiabetic drugs available, namely acarbose and miglitol, which act via competitive inhibition through reversible binding to the catalytic site of α-glucosidase. 30,31 However, the use of these antidiabetic drugs may be associated with gastrointestinal side effects, such as abdominal pain, flatulence, and diarrhoea. 32 Hence, there is considerable interest in finding alternative α-glucosidase inhibitors from natural sources, including food and medicinal plants. To analyze the potential α-glucosidase inhibitory effect, α-glucosidase derived from yeast was incubated with AKE, phlorizin, and quercetin in vitro (Fig. 4). Acarbose served as a positive control. Inhibitory activities (%inhibition) were plotted against concentrations, from which corresponding IC50 values (the concentration of compound that reduces α-glucosidase activity by 50%) were calculated. We found that AKE dose dependently inhibited α-glucosidase activity with an IC50 of 9.35 mg/mL (Fig. 4A). Phlorizin exhibited an IC50 of 1.97 mg/mL and quercetin an IC50 of 0.196 mg/mL, suggesting a high potency of quercetin to inhibit α-glucosidase activity. We conclude from our in vitro experiments that dietary AKE may also reduce α-glucosidase activity in vivo, thereby delaying intestinal absorption of carbohydrates and subsequently retarding glucose entry into the systemic circulation. Indeed, in the present study, mice fed the AKE diet displayed lower blood glucose levels and improved OGTT compared to mice fed the WTD only. Likewise, it was found that mice fed an apple, cherry, or quercetin extract added at a concentration of 0.2% to a high-fat diet exhibited significantly lower blood glucose concentrations than mice fed the high-fat diet only. 33 Jeong et al. 34 discovered in T2DM mice that quercetin at a concentration of 0.04% to 0.08% in the diet was effective in ameliorating hyperglycemia and dyslipidemia in a concentration-dependent manner. In addition, feeding male C57BL/6J mice over 16 weeks an HFD, supplemented with a lower dosage (0.02%) of phlorizin compared to the dosage used in the present study, still lead to decreased plasma glucose, insulin, and glucagon levels and lower incidence of insulin resistance. 35 Furthermore, it was shown in a randomized, controlled, single-blind, crossover human intervention study that polyphenol- and fiber-rich dried fruits as well as green tea attenuated starch-derived postprandial blood glucose and insulin levels. 36 The authors concluded that the pronounced and significant glucose-lowering effect resulted at least, in part, from the inhibition of α-amylase and α-glucosidase activity by plant bioactive compounds.
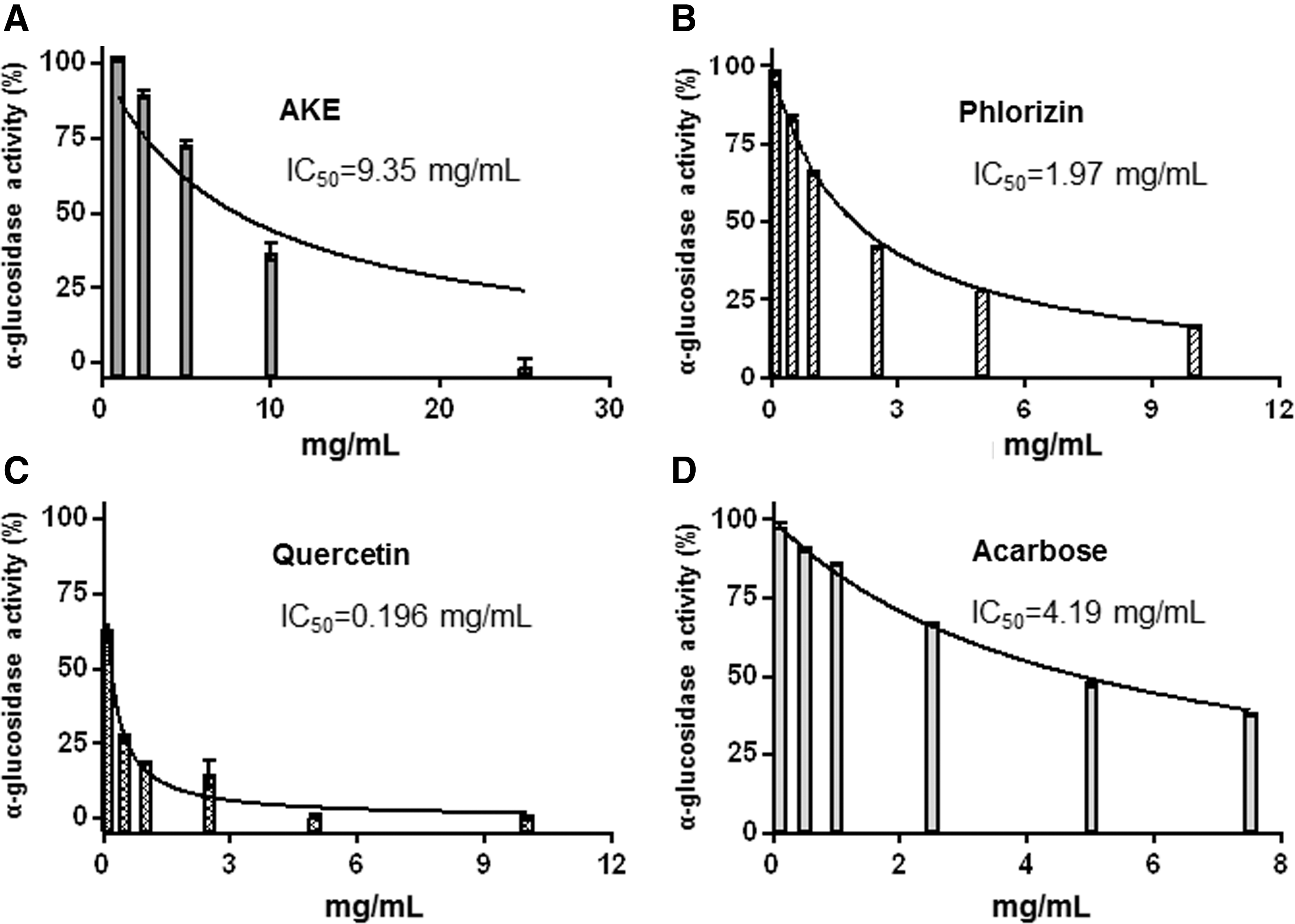
Dose-dependent inhibition of α-glucosidase activity by AKE,
In the present study, we introduced the antidiabetic properties of an AKE rich in phlorizin and flavonoids in vitro and in mice fed a WTD in vivo. We postulate that the combination of apple plus kale results in an improved antidiabetic potency through an additive inhibitory effect on SGLT1 and α-glucosidase activity. We conclude that AKE exhibits antidiabetic properties and has the potential to serve as a natural source of plant bioactive compound in a dietary prevention strategy for T2DM patients. Thus, the present findings on the antidiabetic properties of an AKE in vitro and in laboratory mice should be validated in human intervention studies in the future.
Footnotes
Acknowledgments
This study was supported financially by a grant from the Central Innovation Programme for SMEs (ZIM)—Funding for Innovation (KF2780802AJ3). Furthermore, we thank Dr. Ignacio Ipharraguerre for providing expertise with the Ussing chamber experiments and we further acknowledge the excellent technical assistance of Vivien Schmuck.
Ethical Considerations
The animal experiment was conducted in accordance with the German regulations on animal care and with permission from the responsible authority (V 242-16213/2016).
Author Disclosure Statement
Dr. Henning Vollert is the director of BioActive Food GmbH. All other authors have no competing financial interests to disclose.