
Abstract
Dyslipidemia is caused by disturbances in lipid metabolism that lead to chronic elevations of serum lipids, especially low-density lipoprotein (LDL)-cholesterol and triglycerides, increasing the risk of metabolic syndrome, obesity, diabetes, atherogenic processes, and cardiovascular diseases. The oil from the fruits of Euterpe oleracea (OFEO) is rich in unsaturated fatty acids with potential for treating alterations in lipid metabolism. In this study, we aimed to investigate the effect of OFEO on hyperlipidemia induced by Cocos nucifera L. saturated fat (GSC) in Wistar rats. Chromatographic profile showed that unsaturated fatty acids account for 66.08% in OFEO, predominately oleic acid (54.30%), and saturated fatty acids (palmitic acid 31.6%) account for 33.92%. GSC-induced dyslipidemia resulted in an increase in total cholesterol, LDL-cholesterol, triglycerides, glucose, and liver and abdominal fat, as well as atherogenic processes in the thoracic aorta. OFEO treatment did not reduce hypertriglyceridemia, but did reduce total cholesterol and LDL-cholesterol, thus contributing to the antiatherogenic action of OFEO. OFEO treatment inhibited the formation of atheromatous plaques in the vascular endothelium of the treated rats, as well as those who were treated with simvastatin. The results obtained suggest that OFEO has an antiatherogenic effect in a rat model of dyslipidemia.
Introduction
D
These lipid abnormalities are responsible for 56% of heart disease and are associated with a third of mortality worldwide. Thus, many studies have addressed various treatments to prevent and control this metabolic condition. 5 The biodiversity of the Amazon provides many plants rich in bioactive compounds. The polyphenols and antioxidant unsaturated fatty acids (UFAs) present in Açai (Euterpe oleracea Mart.) have been widely studied in the control of metabolic parameters, 6 metabolic syndrome, 7 atherosclerosis, 8 oxidative stress, 9 hypercholesterolaemia, 10 and inflammatory processes, among others. 11
The E. oleracea Mart. is a member of the family Arecaceae and is commonly known as açaí or “iça-iça” by the Indians, which translates as the fruit that cries. E. oleracea is a typical palm of lowland rainforest from the Amazon region, but can also be grown in upland soils and predominates spontaneously in the states of the northern region of Brazil, extending up the Guianas, Colombia, Ecuador, and Venezuela. 12 E. oleracea Mart. has high agronomic, technological, nutritional, and therapeutic potential. 13
The açai berry fruit is composed of about 13% protein, 48% fat, and 1.5% total sugars, is rich in phenolic compounds (mainly anthocyanins), mono- and poly-UFAs, and dietary fiber, as well as being a good source of minerals such as potassium, magnesium, calcium, phosphorus, sodium, and vitamins E and B1. 10
The oil contained in the açai fruit accounts for ∼50% of its total dry matter, and its lipid profile is rich in mono- and poly-UFAs, which account for more than 75% of its lipid composition, with oleic and linoleic fatty acids being major components. 14
Several studies have described the potential of UFAs in the treatment and prevention of lipid metabolism disorders, especially hyperlipidemia and dyslipidemia. 15,16 In this study, we aimed to investigate the effect of E. oleracea Mart. oil treatment in rats with Cocos nucifera L. saturated fat-induced dyslipidemia.
Materials and Methods
Obtaining the Euterpe oleracea Mart. oil
Oil from the fruits of Euterpe oleracea (OFEO) (Product code 018, date of extraction: 14/08/2015, Lot number–AMO 018-004/2015) was obtained from Amazon Oil Industry, located in the city of Ananindeua, State of Pará, Brazil.
The extraction method has been standardized by the company and cannot be described here because of patent protection.
Obtaining of Cocos nucifera L. saturated fat (GSC)
The GSC was obtained from the Cocos Empire Company—Belo Horizonte Municipal Market, State of Minas Gerais, Brazil. The GSC extraction method was performed by mechanical pressing of endocarp, followed by the addition of water (1:1 [w/v]) and separation of the fat by heating at 80°C, as described in Handayani et al. 17
Transesterification of OFEO and GSC by enzymatic catalysis
OFEO and GSC (150 mg) were added to 5 mL vial bottles, followed by 475 uL of ethanol and 15 mg of acrylic resin of Candida antarctica lipase (CAL-B ≥ 5000 U/g). The reaction was maintained at 32°C under orbital shaking (130 rpm) (Tecnal model TE-421-Brazil) for 24 h. The enzyme reaction was then transferred to a separator funnel and the lower phase (glycerol) was separated, and the upper layer (monoester) was washed with distilled water (2 × 5 mL). The organic phase was dried over anhydrous sodium sulfate and filtered. Finally, the product was purified by column chromatography on a flash silica gel with a mixture of ethyl acetate and n-hexane (9:1) as eluent. 18
Analysis of OFEO and GSC by gas chromatography–mass spectrometry
Coupled gas chromatography–mass spectrometry (GC-MS) analyses were performed on a Shimadzu system/GC 2010 coupled to a self-gun Shimadzu/AOC-5000 and mass detector (Shimadzu MS2010 Plus) with electron impact (70 eV), equipped with a fused silica column of DB-5MS (Agilent Advanced J&W 30 m × 0.25 mm × 0.25 μm). The parameters were as follows: split ratio, 1:20; helium as carrier gas (65 kPa); injection volume, 1.0 μL; injector temperature, 250°C; detector temperature, 250°C; initial column temperature, 50°C for 1 min; and heating rate, 5°C/min to 250°C. The total analysis time was 35 min; the identification of fatty acid esters was performed using the NIST 5.0 equipment library. 19
Animals
This study was approved by the Ethics Committee on Animal Use of Amapá Federal University—UNIFAP under Protocol No. 013/2015 on September 17, 2015–CEUA/UNIFAP.
Thirty-five male Wistar rats weighing 210 ± 20 g were used. The rats were kept in standard cages for rodents in an air-conditioned environment with a controlled temperature of 25°C ± 2°C, humidity of 50 ± 10%, and a photoperiod of 12 h, with free access to standard rodent food and water ad libitum.
Treatments and induction of hyperlipidemia
The animals were divided randomly into five groups (n = 7 per group) and treated orally: (1) treated with 1226 mg/kg OFEO daily for 40 days (OFEO group); 11 (2) treated with 1226 mg/kg OFEO daily for 40 days, plus 2 mL/day GSC from the 20th to the 40th day (OFEO+GSC group); (3) treated with 0.5 mL of distilled water for 40 days and, 2 mL/day of GSC from the 20th to the 40th day (GSC group); (4) treated with 20 mg/kg/day simvastatin for 40 days and, 2 mL day−1 GSC from the 20th to the 40th day (SIN+GSC group); and (5) the DW group, treated with only 0.5 mL of distilled water daily for 40 days. 20
Body mass changes and caloric consumption of the animals
Animal body weight was measured every 4 days throughout the treatment period. Variation in weight increase was calculated by subtracting the final animal weight (F1) from the initial animal weight (F0). The power consumption value was calculated by dividing the food intake by the consumption of OFEO and GSC; these values were expressed in kilocalories (Kcal) according to the energetic properties of the lipids, carbohydrates, and proteins.
Biochemical analysis
After 41 days, the animals were fasted for 12 h and then anesthetized with sodium thiopental 45 mg/kg intraperitoneally. Blood (1.5 mL) was then collected through the retro-orbital plexus and centrifuged for 10 min at 5000 rpm. The collected blood was then subjected to biochemical analyses (glucose, triglycerides, total cholesterol and fractions, aspartate aminotransferase and alanine aminotransferase, urea, and creatinine). All tests were performed using reagents and kits from Doles Reagents and Equipment Lab Industry Co. (Goiânia, GO, Brazil), and samples were analyzed using a UV-VIS UVmini-1240 model spectrophotometer (Shimadzu Corporation, Kyoto, Japan). 21
Removal of organs and aortic analysis by scanning electron microscopy (SEM)
On the 41st day, the animals were euthanized and their organs (liver, kidney, pancreas, spleen, heart, lungs, and abdominal fat) removed and weighed on an electronic analytical balance (Model Bioprecisa FA-2104N). The isolation of the aorta was performed from the aortic arch to the iliac bifurcation. The thoracic region was divided into 0.5 cm sections and analyzed by scanning electron microscopy (SEM, Hitachi Model-TM3030PLUS) for detecting atherogenic processes.
Statistical analysis
Values are presented as mean ± standard error (SE) of the mean. Analysis of variance (one way) followed by Tukey tests for multiple comparisons were used to identify statistically significant (P < .05) differences between groups.
Results and Discussion
Excessive consumption of lipids can promote increased plasma free fatty acid (FFA) content and thus can contribute to the dyslipidemia process, a main risk factor associated with atherogenesis and cardiovascular disease. Saturated fatty acids (SFA) are among the main lipids that contribute to hyperlipidemia, these are partly responsible for the elevated serum triglycerides and LDL-cholesterol levels seen in dyslipidemia, and have close correlation with the development of atherosclerosis and chronic inflammation in blood vessels. 22,23
Our chromatographic data (Table 1 and Fig. 1) indicate that the GSC used in this study to induce dyslipidemia contains 92.64% SFA, especially lauric acid (57.8%), myristic acid (17.30%), and palmitic acid (9.40%). These data are consistent with those obtained in other studies, where GSC is reported as being greater than 90% saturated fats and to include large amounts of lauric and myristic acid. 24,25

Gas chromatograms (gas chromatography–mass spectrometry) of ethyl ester derived from GSC
MS database (NIST 5.0).
% of FAEE corresponding fatty acid.
Animal models of induced hyperlipidemia by GSC administration have been described. The high saturated fat content of GSC can greatly increase the rate of cholesterol production, as well as the proportion present as triglycerides. 20,26
OFEO chromatographic data show a 66.08% UFA content, of which oleic and linoleic acid accounted for 54.30% and 4.9%, respectively (Table 1 and Fig. 1). The remaining SFA (33.92% of total) was rich in palmitic acid (31.6%). These results are consistent with other analyses of OFEO. 11
Various data have suggested that UFAs have beneficial effects on lipid profile and dietary UFAs can contribute to a reduction of adipose tissue formation. 4 This is consistent with our body weight data. The OFEO group had a lower body weight than the GSC group (130.2 ± 11.65 g vs. 148.9 ± 8.2 g, P < .001) (Fig. 2). This reduction in body weight in the OFEO group may be due to the high UFA content of the OFEO, since UFA supplementation is associated with reduced hyperplasia and hypertrophy of adipocytes. UFAs reduce adipose tissues by modulating the metabolism through a stimulation of β-oxidation, which leads to an induction of PPAR-α, ultimately controlling the differentiation and proliferation of adipose cells. 27
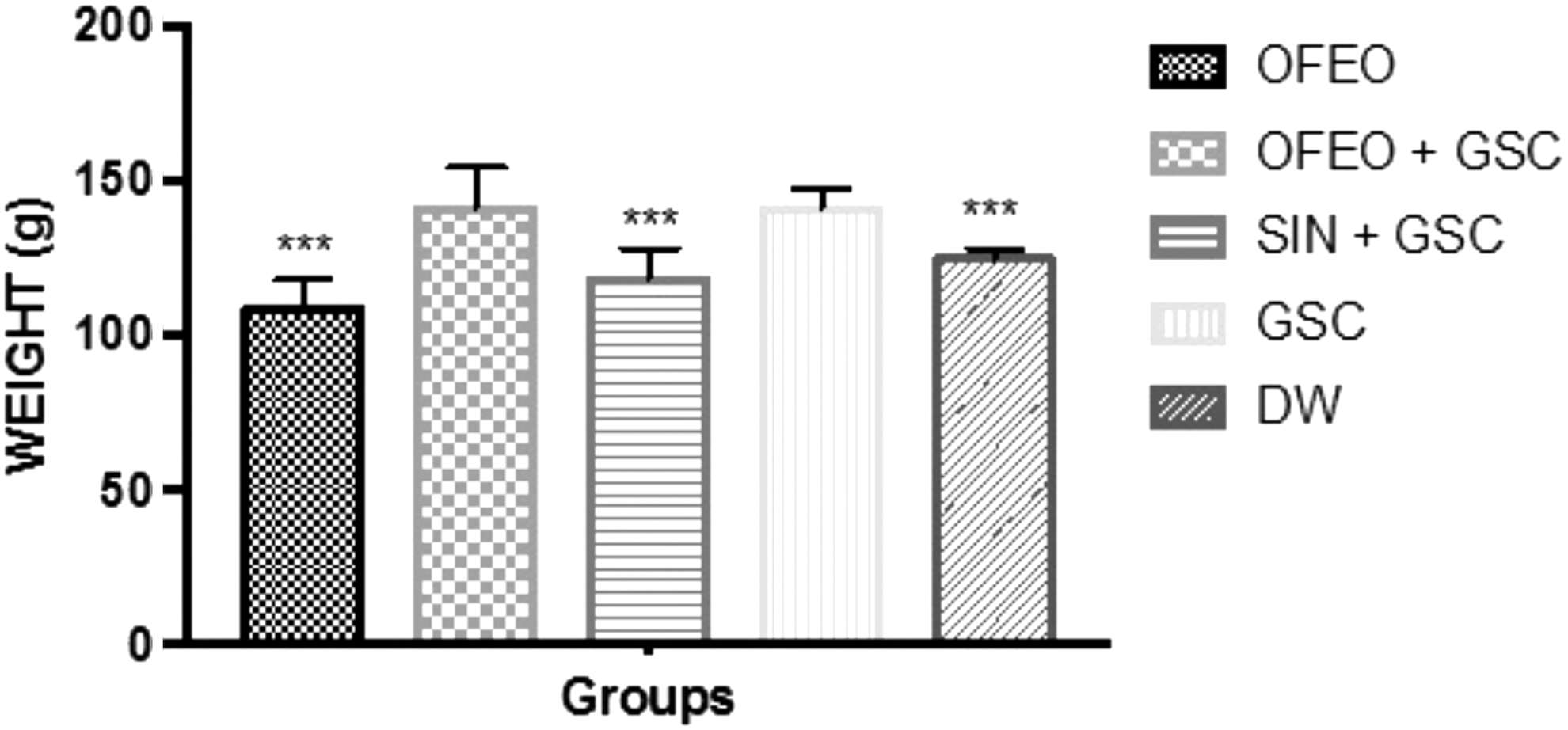
Body weight variation of the animals after the period of treatment and induction of hyperlipidemia by GSC. The bars represent mean ± SE of the mean (n = 7/group). ***(P < .001) statistically significant results compared with GSC. One-way ANOVA followed by Tukey test. ANOVA, analysis of variance; SE, standard error.
The DW and SIN+GSC groups were also significantly different from the GSC group (both P < .001). The increase in body mass observed in the OFEO+GSC and GSC groups is partly related to the increased daily calorie consumption of these rats; diets rich in SFAs provide considerable amounts of energy and contribute to the increase of adipose tissue. 22 Of the five groups studied here, the GSC (657.4 ± 24.8 Kcal/day) and GSC+OFEO (632.1 ± 18.7 Kcal/day) groups had the highest caloric consumptions (Fig. 3). The OFEO group consumed significantly less than the GSC group.
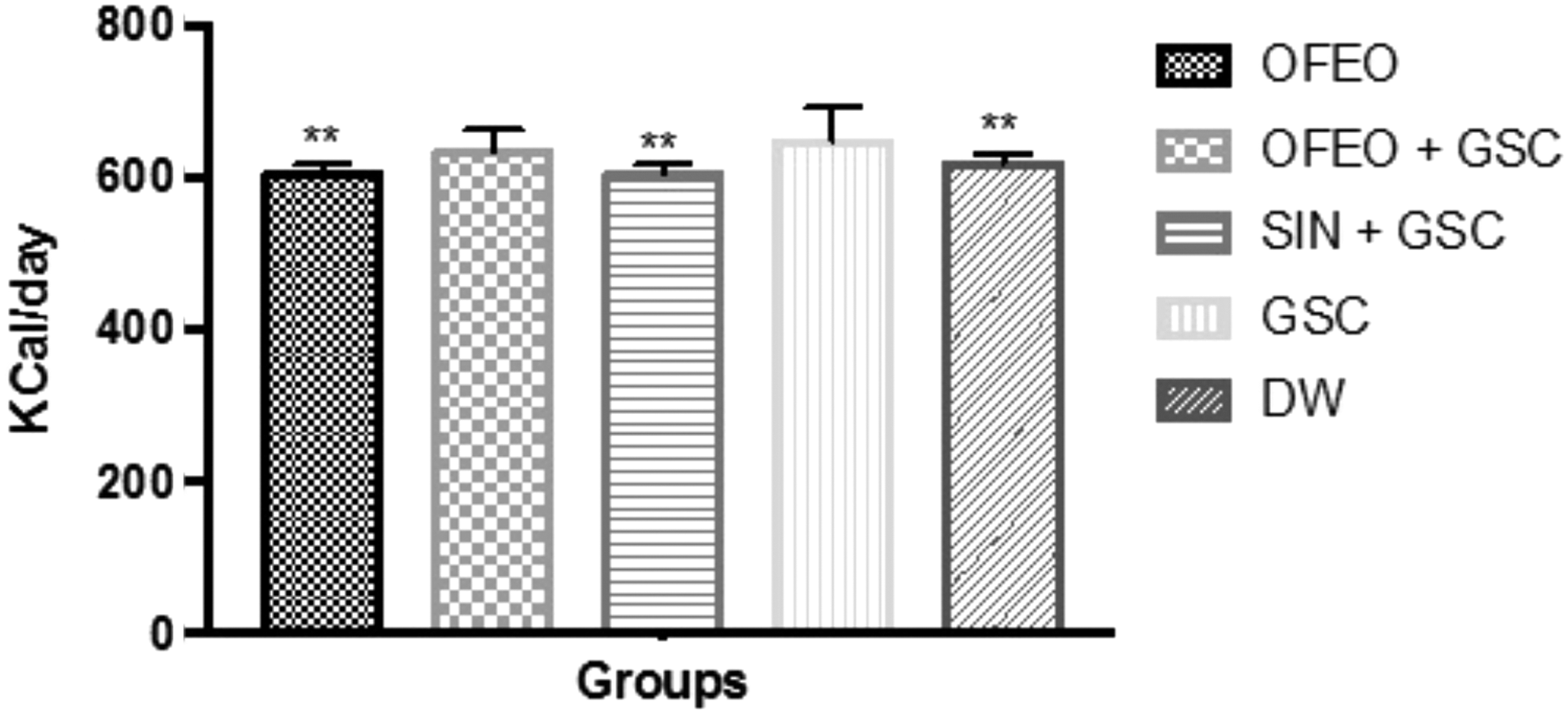
Average values of daily caloric intake of different groups during the period of treatment and induction of hyperlipidemia by GSC. The bars represent mean ± SE of the mean (n = 7/group). **(P < .01) statistically significant result compared to the GSC. One-way ANOVA followed by Tukey test.
UFAs can reduce food intake and thus prevent body weight increases. Increased UFA plasma concentration can also modify lipid metabolism, stimulating the expression of mitochondrial proteins involved in thermogenesis, thus greatly increasing daily energy expenditure and promoting the reduction of adipose tissue. 22,28
We found that the groups with the highest body mass gain and caloric intake (the GSC and OFEO+GSC groups) also showed a significant increase in liver and abdominal fat mass compared to the other groups (P < .001) (Table 2). This increase in the absolute mass of the organs may be due to excess SFA contained in the GSC. This is supported by studies showing that excess administration of SFA leads to lipid biosynthesis in the liver and increased hepatic flow of FFA, which contribute to the accumulation of lipids in the liver and lipogenesis. 26
The values represent the median ± SE of the mean (n = 7/group). #(P < .001) and **(P < .01) represent statistically significant results compared to GSC. One-way ANOVA followed by Tukey test.
ANOVA, analysis of variance; SE, standard error.
The GSC-induced hyperlipidemia group had a significantly higher glycemic value than the DW and OFEO groups (P < .01), since the OFEO+GSC group, although it had reduced blood glucose, was not statistically significant when compared to the GSC group, showing that treatment with OFEO had no effect on blood glucose, as well as the group treated with SIN+GSC (Table 3).
The values represent the mean ± SE of the mean (n = 7/group). * P < .05, ** P < .01, and # P < .001 represent statistically significant results, acompared with OFEO, bcompared with OFEO+GSC, ccompared with GSC, dcompared with SIN+GSC, and compared with #cDW. Test One-way (ANOVA) followed by Tukey test.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; LDL, low-density lipoprotein.
The increased blood glucose of the GSC group may be due to an increased SFA intake, which contributes to the accumulation of body fat; an excessive intake of energy from carbohydrates and lipids can contribute to resistance to insulin action, thereby leading to a chronic and gradual increase in glycemia. 29,30
The GSC and OFEO+GSC groups had significantly higher total cholesterol than the DW group (P < .001); however, the OFEO+GSC group had significantly lower total cholesterol than the GSC group. These data show that OFEO acts to prevent GSC-induced hyperlipidemia. The SIN+GSC had significantly lower total cholesterol than the GSC group. The total cholesterol of the OFEO group was similar to that of the DW group, suggesting that OFEO alone does not promote an increase in cholesterol (Table 3). These findings are in line with other studies involving E. oleracea species. 6,7,10
The OFEO group presented a significant increase (P < .05) in triglyceride levels in relation to the DW group. These data, indicating that OFEO+GSC can contribute to increasing triglyceride levels, are according to Celeski et al., 20 where GSC administration in rats produced a 171% increase in triglyceride levels. This increase may be related to the presence of 33.92% AGS in the oil composition; however, it did not induce lesion in the endothelium, a fact that may have been confirmed in the histopathological analyzes by SEM. The triglyceride levels of the SIN+GSC group were not significantly different from the DW group (Figs. 4 and 5, and Table 3).
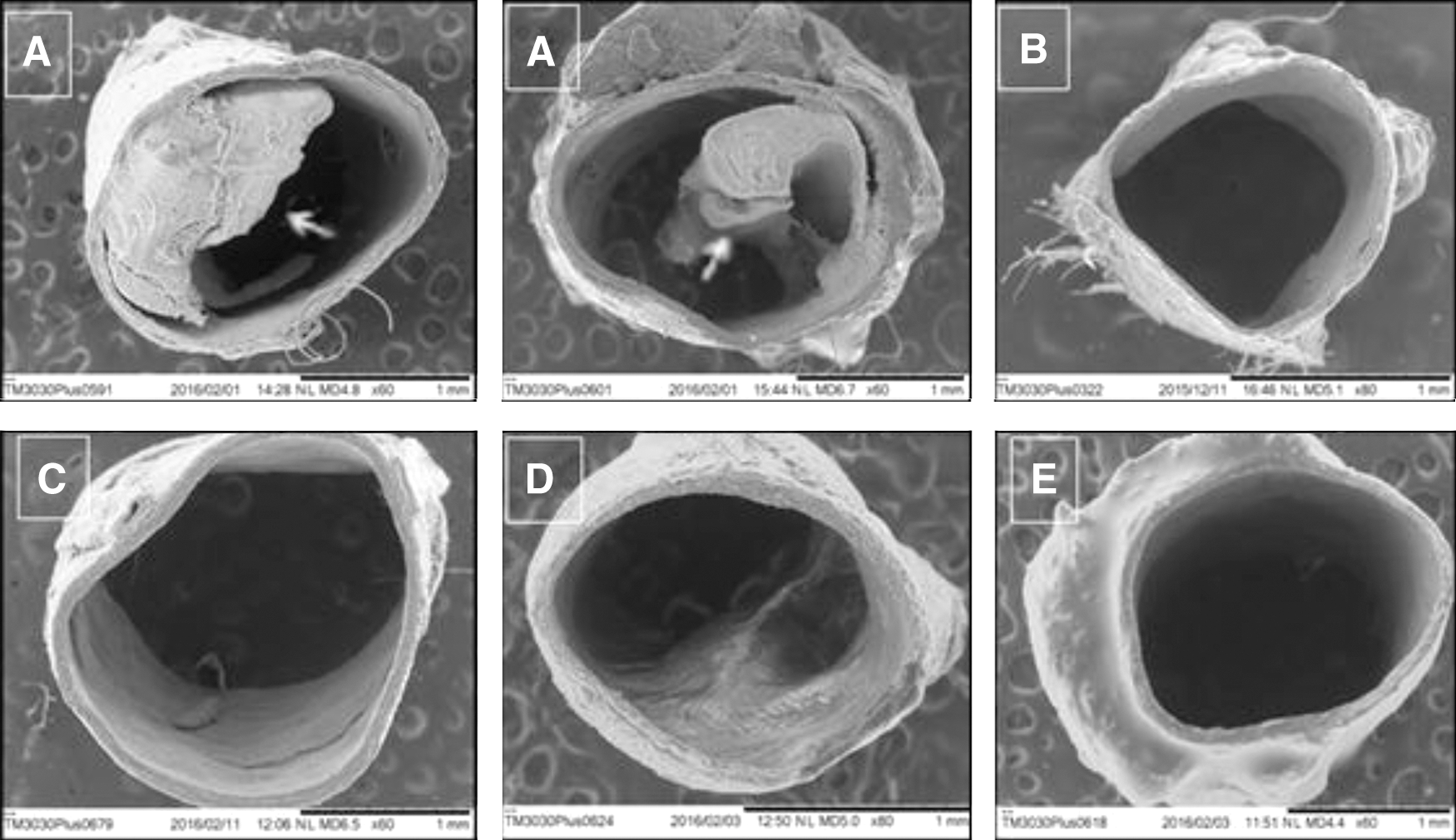
SEM photomicrographs of cross-section of the thoracic aorta. The arrows indicate the formation of atheromatous plaques in the arterial endothelium. Were:

SEM photomicrographs of longitudinal section of thoracic aorta. The arrows indicate the formation of atheromatous plaques in the arterial endothelium. Were:
The OFEO group had the highest HDL-cholesterol levels (24.7 ± 2.9 mg/dL), but these were not significantly different from the GSC group. The GSC group had the highest LDL-cholesterol levels (55.5 ± 2.2 mg/dL), and the OFEO and OFEO+GSC groups showed reduced and significant amounts compared to the GSC group, indicating that OFEO treatment was able to prevent LDL-cholesterol increases in our disease model. Urea, creatinine, and transaminases did not differ significantly between the groups, indicating that none of the treatments were harmful to the kidney or liver systems (Table 3). 21
On the positive results of lipid profile of the SIN+GSC group are highlights that are related to the mechanism of action of statins. Statins act by inhibiting the HMG-CoA reductase enzyme activity, thereby preventing the formation of mevalonate, contributing to a reduction in hepatic synthesis of cholesterol and, consequently increasing synthesis of LDL-cholesterol receptors in hepatocytes, thus increasing their capitation of the circulation to restore the intracellular cholesterol. 5,31
Our biochemical data show that OFEO treatment was unable to prevent GSC-induced hypertriglyceridemia in the rats; however, OFEO was able to reduce total and LDL-cholesterol levels, possibly due to the lipid profile of OFEO, which includes a rich composition in UFAs, including oleic and linoleic acid. 13,32
Free radicals, including reactive oxygen species (ROS) and reactive nitrogen species (RNS), are responsible for the oxidation of LDL. ROS include the superoxide anion, the hydroxyl radical, and hydrogen peroxide. RNS are mainly nitric oxide and peroxynitrite. 33,34 UFAs can act as an antioxidant, primarily preventing lipid peroxidation; since UFA molecules contain more double bonds, they are more susceptible to loss of hydrogen to ROS and RNS. 15 In vitro treatment with UFAs significantly reduces free radical levels and has a beneficial effect on lipid profile, mainly by reducing LDL-cholesterol levels. 4,35
The formation of atherosclerotic plaques begins with the aggression against vascular endothelium due to various risk factors. An increase in plasma LDL-lipoprotein concentrations, which after entering in the intima of arteries, undergo oxidation in the pro-oxidizing environment, causing the exposure of multiple neo-epitopes that make immunogenic and trigger inflammation and the recruitment of immune cells, particularly monocytes. These immune cells migrate into the subendothelial space where they differentiate into macrophages and capture the oxidized LDL. Called foam cells, they are filled with lipids, the main component of the fatty matrix in the formation of atheroma. 36 –38
In the formation of atherosclerosis in animal models, the development of atherosclerotic lesions is accelerated when the animals are given a high SFA content diet. The thoracic and abdominal aorta are the most commonly affected arteries during atherosclerosis. 39,40 This is in line with our data, which show the formation of atherosclerotic plaques in the vascular endothelium of the GSC group (Figs. 4 and 5). Analyses of the OFEO and OFEO+GSC groups show that OFEO has antiatherogenic properties; there were no atheromatous plaques in the vascular endothelium. This can be accounted for by the ability of OFEO to lower LDL-cholesterol; LDL is the key lipoprotein in early atherogenesis process. The GSC+SIN and DW animals did not form atherosclerotic plaques. 41,42
Conclusion
In this study, we found that OFEO reduces total cholesterol and LDL-cholesterol and prevents the atherogenic process in rats with Cocos nucifera L. saturated fat-induced hyperlipidemia. Thus, we propose that OFEO has antihypercholesterolemic and antiatherogenic properties.
Footnotes
Acknowledgment
This study was supported by National Council for Scientific, Technological Development–CNPq (Biotec—Process 402332/2013-0), and Universidad Autônoma de México – Programa de Estancias de Investigación (PREI).
Author Disclosure Statement
No competing financial interests exist.