
Abstract
Pleurotus djamor is an edible mushroom that has medicinal properties. This study aimed to assess the in vitro activity of P. djamor extracts and fractions against Haemonchus contortus eggs and exsheathed infective larvae (L3). Crude hydroalcoholic extracts were obtained by maceration and fractions were obtained through chromatography. Metabolite identity was determined using gas chromatography/mass spectrometry (GC-MS) analysis. The results showed that P. djamor extracts had no significant activity against eggs at the concentrations used. However, the extract showed 98.7% and 77% larval mortality 72 h postconfrontation at 320 and at 160 mg/mL, respectively. The chromatography analysis resulted in 23 fractions that were eventually grouped into three fractions (E1, E2, and E3). These fractions showed the following egg hatching inhibition percentages: E1 = 100, E2 = 38.7, and E3 = 5.5 at 10 mg/mL concentration 72 h postexposure. Likewise, larval mortality percentages after this period were 90.6, 100, and 0.44 at 40 mg/mL (P < .05), respectively. The GC-MS showed five major compounds in E1 fraction, including four fatty acids: (i) pentadecanoic, (ii) hexadecanoic, (iii) octadecadienoic, (iv) octadecanoic acid, and one terpene identified as β-sitosterol. We concluded that the edible mushroom P. djamor possesses nematicidal metabolites, which could be used as an alternative anthelmintic treatment.
Introduction
T
Various metabolites from edible mushrooms with nematicidal activity have been previously identified: xanthonone, pleurotin, leucopleurotin, p-anisaldehyde, p-anisyl alcohol, and so on. 28 Likewise, other genera/species of edible mushrooms can have nematicidal activity. The mushroom Coprinus comatus produces spiny structures that secrete nematode-immobilizing toxins that cause cuticular damage in the bacteriophagous nematode Panagrellus redivivus. 29,30 Nematicidal activity has been found in 23 edible mushroom species of the genus Pleurotus. 11 However, so far, there are no reported nematicidal metabolites produced by P. djamor. So, this study was based on the hypothesis that the edible mushroom P. djamor produces metabolites with nematicidal activity with potential as natural biocompounds for controlling H. contortus as one of the most pathogenic nematodes affecting the sheep industry.
This study aims to identify metabolites produced by P. djamor with nematicidal activity against H. contortus infective larvae and eggs.
Materials and Methods
Obtaining P. djamor basidiomata
Basidiomata obtained from P. djamor (strain): ECS-0123 were isolated from Talquián village at Union Juárez, Chiapas, Mexico. Taxonomic identification was performed by using the internal transcribed spacer (DNAr) and recorded at GenBank (accessing number: GU722265). 31 Laboratory basidiomata were produced at the Tropical Mushroom Laboratory of El Colegio de la Frontera Sur Unidad Tapachula, Chiapas, Mexico. Mushrooms were produced following the methods in Avendaño-Hernández and Sánchez. 32
Extract preparations
Fresh basidiomata (2.205 kg) were chopped into small fragments (1 cm diameter) and put into three Erlenmeyer flasks. Then, 5 L of a hydroalcoholic mixture (ethanol/water 60:40) was added to each flask for the maceration process. Fungi material was allowed to incubate for 24 h at room temperature (18–25°C).
After maceration, the crude extract was filtered through Whatman paper (#4) and concentrated using a rotavapor (Heidolph 6, Germany) until the liquid residue was eliminated. The hydroalcoholic extract (HE) was lyophilized using a Heto Drywinner DW3 lyophilizer. The dried HE was kept at −4°C until use. Mushroom material yield was 48 g. 33,34
HE chromatographic fractioning
Thirty grams of HE was adsorbed using 30 g of silica gel (70–230 mesh) and put into an open chromatography column (34 × 700 mm) containing 50 g of silica gel (MERCK; 70–230 mesh) and eluted into a gradient system in the mobile phase. The system started using 100% dichloromethane and its polarity was increased with 10% methanol. Five 500 mL samples were collected from each eluted system. Resulting fractions were analyzed and biologically assessed following the methodology described by Osuna-Torres et al. 35
Obtaining H. contortus eggs
Haemonchus contortus eggs were recovered from feces of an egg-donor sheep artificially infected with 350 H. contortus infective larvae (L3) per kilogram of body weight. After a 21-day prepatent period, fecal samples were collected directly from rectum of this animal. Fresh feces were crushed and mixed with tap water in a plastic bowl to form a homogeneous paste and sieved through meshes from different open sizes (35, 100, 200, and 400 U.S. standard sieve) and then washed with tap water. The obtained filtrate was centrifuged at 366.51 rad/s for 5 min in a 40% sucrose density gradient to separate detritus from the nematode eggs. The nematode eggs were obtained by micropipette aspiration of the white phase formed after. Eggs were rinsed with sterile water ( × 4) to remove sucrose residues. The number of eggs per gram of feces (EPG) was estimated using the McMaster technique. 36,37
Obtaining H. contortus infective larvae (L3)
Haemonchus contortus (L3) were obtained from coprocultures of egg-containing feces incubated in plastic bowls at room temperature (18–25°C) for 5 days. 38 Infective larvae exsheathment was achieved by exposure of larvae to 6% sodium hypochlorite for 10 min. Larvae were then washed with distilled water by centrifugation for 1 min at 261.79 rad/s ( × 4) to eliminate the sodium hypochlorite. Exsheathed larvae were recovered from the pellet formed after centrifugation. 38
Bioassays
In vitro assessment of P. djamor HEs against H. contortus eggs and infective larvae (L3)
Nematode/extract confrontations were carried out in 96-well microtitration plates (MTPs).
Two bioassays were carried out: one to assess HE activity against nematode eggs and the other to evaluate HE activity against larvae. Fifty microliters of HE and 50 μL of an aqueous suspension containing either 100 eggs or 200 exsheathed larvae were put in each well (n = 4). Final HE concentrations in the nematode egg bioassay were 4.8, 9.7, 19.5, 39, 78, 100, 156, 312, and 625 μg/mL. HE concentrations in the larval assay were 0.312, 0.625, 1.25, 2.5, 5, 10, 20, 40, 80, 160, and 320 mg/mL. A negative control of distilled water and a positive control of commercial ivermectin at 5 mg/mL were used to set a baseline. The MTPs were covered with foil and incubated at 28°C (±1) (Rios Rocha; EC-33). The egg hatching inhibition (EHI) was assessed at 48 and 72 h postconfrontation. Larval mortality was assessed at 24, 48, and 72 h postconfrontation. The results of both bioassays were determined by counting the number of larvae hatched from eggs (egg-hatching inhibition) in ten 5 μL aliquot drops placed on a glass slide. Larval mortality was assessed using the slide method previously described. 37
Selection of the proper dissolvent for HE fractions
An assay to identify the best dissolvent for the fractions was established as follows: One milligram of each fraction to be assessed was deposited on a slide and a 50 μL drop of each dissolvent was put on the fraction sample and mixed with a metal needle to see if the fraction samples were properly dissolved. The following dissolvents were used: (i) distilled water, (ii) 5% ethanol, and (iii) 2.5% dimethyl sulfoxide (DMSO).
Assessment of P. djamor fraction activity against H. contortus eggs and larvae
In both eggs and larvae bioassays, 1.5-mL conical microtubes (Eppendorf) were used. For each assessed fraction, the following mixture was used: 2 μL fraction, 10 μL water, and 88 μL of an aqueous suspension containing either 100 eggs or 200 exsheathed larvae, to give a final volume of 100 μL. The egg fraction concentration was 10 and 40 mg/mL for larvae. DMSO (2.5%) was used as a negative treatment control, and 1.2 mg/mL commercial ivermectin was used as a positive control (n = 3). Microtubes were covered with foil and incubated at 28°C (±1). A single time point at 72 h postconfrontation was considered. The results of both egg and larval bioassays were evaluated using the criterion previously described. 37 Both eggs and larvae were evaluated under magnification post-HE exposure for possible alterations to body integrity.
The percentage results of the EHI and larval mortality experiments were estimated based on the following formula
39
:
Statistical analysis
A completely randomized design was used. 40 An analysis of variance was made, followed by a means comparison by Tukey's test (α = 0.05). The statistic program SAS 9.2 was used. 41
Gas chromatography/mass spectrometry analysis
The fractions showing the highest nematicidal activity either against eggs or larvae were the selected ones to be analyzed by gas chromatography/mass spectrometry (GC-MS) using gas chromatograph equipped with a quadrupole mass detector in electron impact mode at 70 eV. Volatile compounds were separated with an HP 5MS capillary column (25 m long, 0.2 mm ID, 0.3 μm film thickness). The oven temperature was set at 40°C for 2 min, then programmed from 40°C to 260°C at 10°C/min, and maintained for 20 min at 260°C. Mass detector conditions were as follows: interphase temperature 200°C and mass acquisition range 20–550. The temperatures of the injector and detector were set at 250°C and 280°C, respectively. The splitless injection mode was carried out with 1 μL of each fraction (3 mg/mL). Helium at a flow rate of 1 mL/min was used as the carrier gas. Comparing experimental mass spectra with those of the National Institute of Standards and Technology 1.7 Library did identification of volatiles. In addition, a standard solution of C7–C40 alkanes was used to obtain the retention index of compounds and for literature comparison purposes. 37,42
Results
The P. djamor HE yield was 2.2% from 48.2 g of vegetative material with respect to the fresh mushroom weight. The HE chromatography fractioning resulted in 23 fractions; these were initially grouped in 12 reunions. From these groups, reunions 1–7 were eventually regrouped into three fractions that were named as E1, E2, and E3 fractions for their assessment (Table 1). The crude HE with the highest larval mortality (98.7%) was 320 mg/mL, 72 h postconfrontation (Table 2); however, it had almost no hatching inhibition (Table 3). E1, E2, and E3 fractions showed 100%, 38.7%, and 5.5% hatching inhibition at 10 mg/mL against H. contortus eggs (Table 4). After exposure to P. djamor fractions, morphological changes were observed in both eggs and larvae of the first evolutive stage that succeed in hatching (Fig. 1). In this Figure, a normal egg and a normal larva (negative control in DMSO) are shown. In larval bioassay, E1, E2, and E3 fractions showed 90.6%, 100%, and 0.44% mortality, respectively, at 40 mg/mL concentration (Table 5). Morphological changes associated with the assessed fractions are shown in Figure 2.
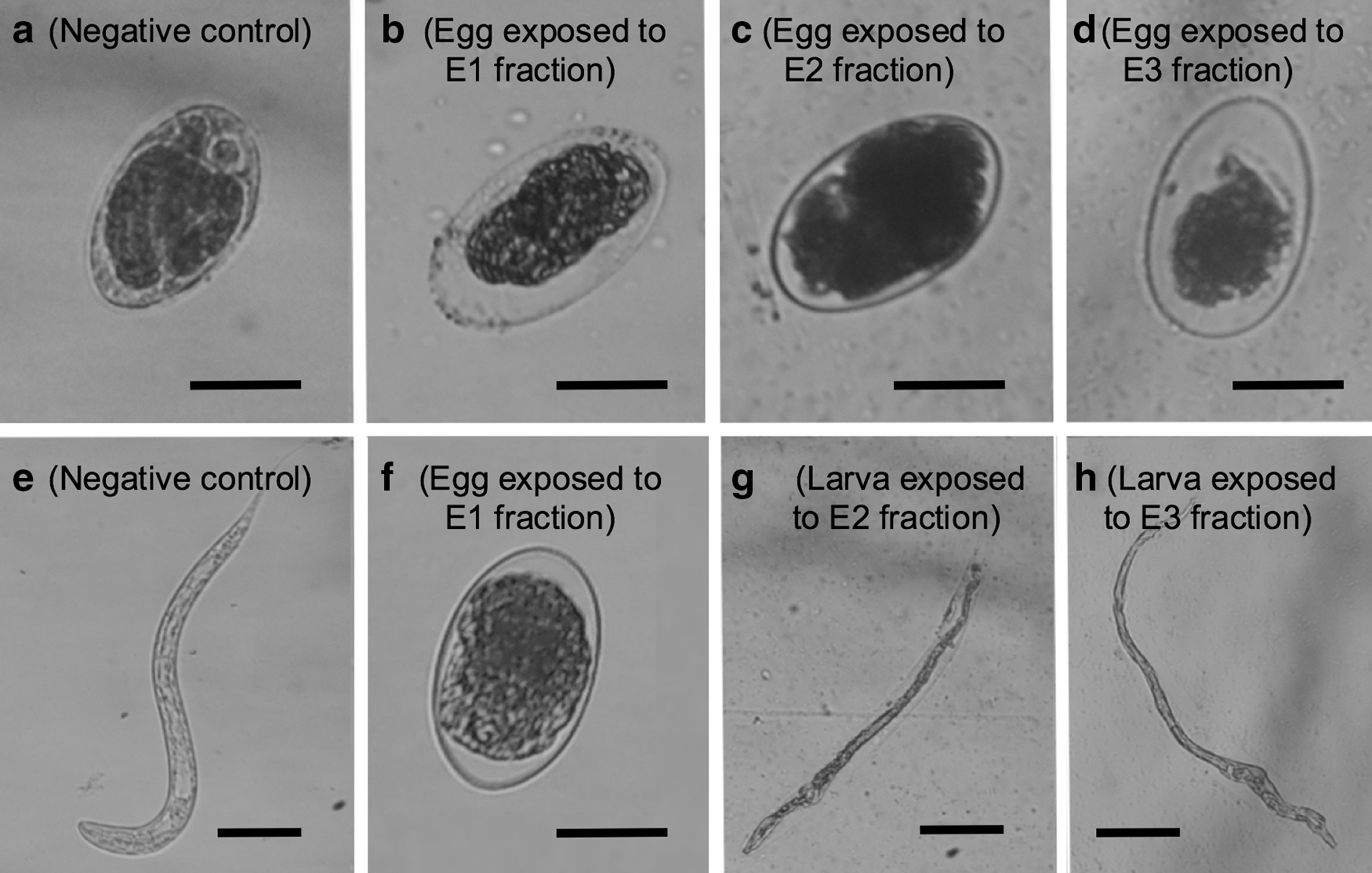
Haemonchus contortus eggs and larvae after 72-h exposure to Pleurotus djamor E1, E2, and E3 fractions.
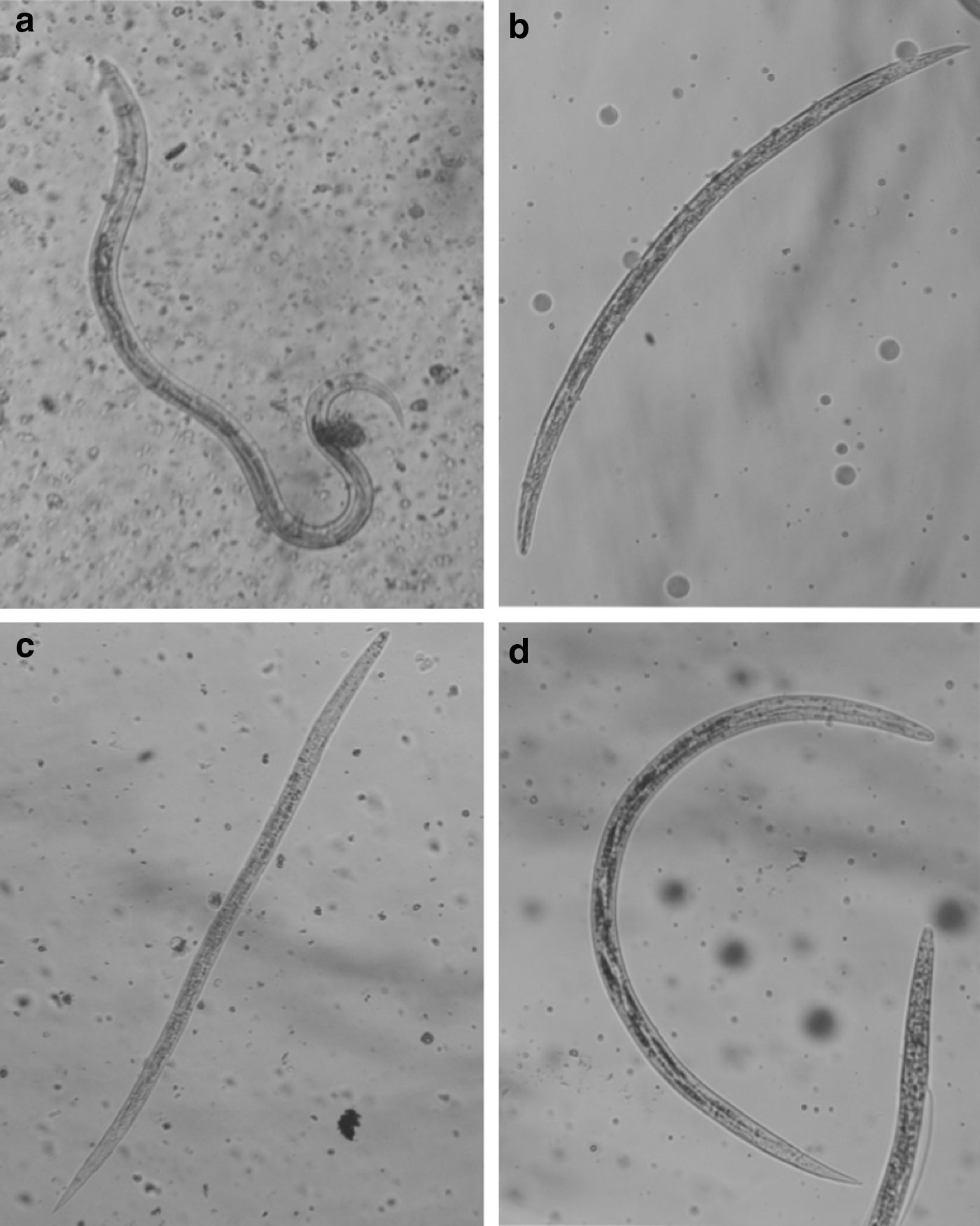
Haemonchus contortus exsheathed infective larvae (L3) after 72-h exposure to Pleurotus djamor E1 and E2 fractions.
n = 4 (wells); 10 aliquot-drops per well. Note: The adjusted larval mortality was obtained considering the total amount of living larvae in the control group as 100% viability.
P ≤ .05. a–fThe same letter in the same column indicates that the values are not significantly different (Tukey test).
n = 4 (wells); 10 aliquot-drops per well.
P ≤ .05. abThe same letter in the same column indicates that the values are not significantly different (Tukey test).
n = 3 (wells); 10 aliquot-drops per well.
P ≤ .05. abcThe same letter in the same column indicates that values are not significantly different (Tukey test).
DMSO, dimethyl sulfoxide.
n = 3 (wells); 10 aliquot-drops per well.
The adjusted larval mortality was obtained considering the total amount of living larvae in the control group as 100% viability.
P ≤ .05. abcThe same letter in the same column indicates that the values are not significantly different (Tukey test).
GC-MS revealed that the most active fraction, E1, contained the following fatty acids: pentadecanoic acid (32.025 min), hexadecanoic acid (34.219 min), 9,12 octadecadienoic acid (37.693 min), octadecanoic acid (37.917 min), and a terpene identified as β-sitosterol (55.225 min) in 4.602%, 21.984%, 62.092%, 5.778%, and 0.974%, proportions, respectively (Fig. 3).
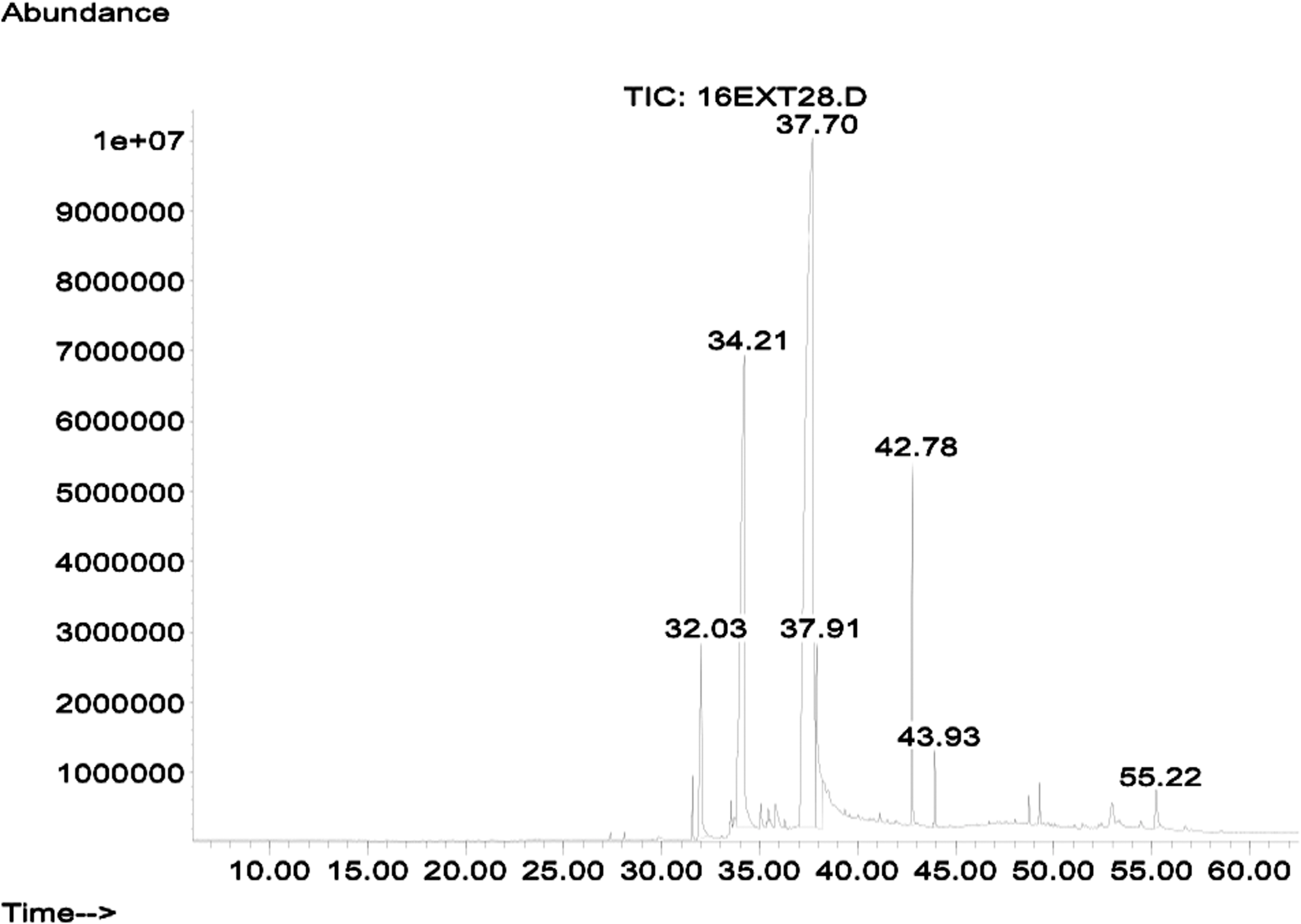
Chromatogram of E1 fraction (from gas chromatography/mass spectrometry analysis) showing the retention times of compounds.
When eggs and larvae were examined under the microscope after confrontation with E1, E2, and E3 fractions, the authors found changes in larval body integrity. Most of the eggs exposed to the E2 fraction showed embryos with a loss of the inner cellular structure, giving a dark, diffuse appearance (Fig. 1c). Eggs exposed to the E1 and E3 fractions showed a small, dark, and diffuse mass in the middle of an otherwise empty egg. Larvae exposed to E2 and E3 fractions showed a substantial reduction in their thickness and a loss of their intestinal cell structure, giving an appearance of a small segment of a plant root (Fig. 1g, h).
Discussion
Mushrooms are the main producers of biologically active metabolites that have potentially therapeutic effects in mammals. 43 –45 These metabolites include more than 200 bioactive compounds with nematicidal activity. 11 Most of edible mushrooms from Lentinula edodes, Agaricus bisporus, and P. ostreatus have been studied to identify their potential medicinal properties. 46 Particularly, the genus Pleurotus spp. has been explored as a possible tool against parasitic nematodes of agriculture and livestock importance. 47,48 However, there is so far, no information about the possible nematicidal activity of P. djamor strains against either phytoparasites or animal parasitic nematodes.
The results of this study suggest that the following fatty acid compounds possess either ovicidal or larvicidal activity against the sheep parasitic nematode H. contortus: pentadecanoic acid, hexadecanoic acid, 9,12 octadecadienoic acid, octadecanoic acid, and one terpene called β-sitosterol. This is in contrast to previous evidence of nematicidal activity associated with a fungal stress response metabolite (produced in the fruiting bodies after damage) identified by Stadler and Sterner. 49 This work showed a higher nematicidal activity in physically damaged mushrooms than in undamaged ones. The present study shows a wide variety of potentially nematicidal compounds in E1, E2, and E3 fractions of P. djamor HE. In fact, these compounds' abundance was the reason why we decided to assess these fractions for nematicidal activity.
In a similar vein, HEs obtained from P. ostreatus (ECS-1123 strain) caused 49% mortality against exsheathed H. contortus (L3) at 80 mg/mL concentration after 72 h postconfrontation. 50 This author also found that using a butanolic fraction from P. ostreatus extracts had a 73% mortality rate against sheathed H. contortus (L3) 48 h postconfrontation. In another study, the HE from P. ostreatus fruit bodies and mycelia at 240 mg/mL showed 82.7% and 96.8% mortality, respectively, against H. contortus eggs and L3 larvae. Meanwhile, in another assay, Valdez 51 reported a hatching inhibition range between 90% and 100% when using HE obtained from P. ostreatus mycelia at different concentrations (from 80 to 240 mg/mL) at 48 and 72 h postconfrontation; similarly, a total EHI using the spent mushroom substrate (SMS) was recorded after 72 h postconfrontation at 3.2 and 6.4 mg/mL concentrations.
Likewise, four P. djamor strains (ECS-0123, ECS-0127, ECS-0142 y, ECS-0143) were assessed to identify their in vitro larvicidal activity against H. contortus (L3) and were found to have different larvicidal activities. Activities of 96.1% and 93% against sheathed larvae were obtained with the ECS-0127 and ECS-0142 strains. In addition, this author recorded 95% larvicidal activity with the ECS-0123 strain. The other strains showed a lower larvicidal activity (<50%). 52
In another study, six HEs obtained from P. djamor fruit bodies produced in different substrate mixtures were assessed against exsheathed H. contortus (L3). Five of these strains showed a larval mortality greater than 95% at 72 h postconfrontation when applied at a 200 mg/mL concentration. The remaining strain showed 75.5% larval mortality. 53 These records are evidence that P. djamor and P. ostreatus contain compounds with nematicidal activity. It is interesting to remark that in most records, the HE from fungal mycelia and SMS showed the highest nematicidal activities. The two highest HE concentrations (160 and 320 mg/mL) used against larvae showed the highest larval mortality (higher than 77% and 98%; respectively). This fact can be considered as successful, however, since the point of view of such a high HE concentration could be little practical. In the case of the ovicidal activity (EHI), the results obtained with E1 fraction, only a little concentration (10 mg/mL) was sufficient to achieve 100% activity. It is important to get a correlation between yield and the concentration necessary to obtain the highest biological activity. This could be a limitation in future practical applications as an anthelmintic product; however, the compounds identified as P. djamor metabolites in the present study could be explored to be used as potential anthelmintics.
This study seems to be the first record about the HE fractions obtained from P. djamor fruit bodies against any nematode of importance in the sheep industry. So, there are no records available for comparison of our results.
On the contrary, the records published about the nematicidal activity from Pleurotus species showed evidence about active compounds in these species present not only in the fungal fruiting body and mycelia but also in the SMS. 50 –53
Other edible mushrooms have also shown nematicidal properties. For example, P. ferulae showed in vitro nematicidal activity against P. redivivus and Bursaphelenchus xylophilus. 54,55 Likewise, P. levis showed in vitro activity against P. redivivus. 54 Similarly, P. corticatus showed lethal activity against P. redivivus. 56 In a recent study, aqueous extracts obtained from the basidiomycetes Agaricus blazei Murril showed a high nematicidal activity (100%) against H. contortus in sheep. 57
On the contrary, one of the most promising uses of SMS is the fact that after fungal growing, this material could content biomolecules with nematicidal activity. This could be beneficial, since such a low-cost substrate could be used to produce natural anthelmintic against ruminant parasitic nematodes. A number of important applications have been identified for the SMS, including agriculture fertilizers, bioremediation tools for heavy metal contamination, and as agents of pesticide bioremediation. The SMS from several edible mushrooms has been found to be nematicidal against nematodes of importance in the agriculture industry, such as the root-knot nematode Meloidogyne incognita, a parasitic nematode that devastates economically important crops and vegetables. 58
In our EHI and larval mortality assays, we observed changes in the egg and larval body structures; such changes were likely the cause of death and can be attributed to the effects of the compounds in the assessed fractions. Similar findings were reported in other studies using molecules, that is, cysteine proteases, obtained from plants such as Ananas comosus, Actinidia deliciosa, 59 and the aqueous extract from the leguminous plant Acacia mearnsii. 60
A number of metabolites obtained from edible mushrooms with nematicidal activity have been reported. In 1992, Kwok et al. 61 reported the presence of trans-2-decanoic acid in P. ostreatus, a compound that paralyzes 95% of the bacteriophagous nematode Panagrellus redivivus when applied at 300 μg/mL.
In another study, six new compounds with nematicidal activity against the bacteriophagous nematode Caenorhabditis elegans were isolated from P. pulmonarius submerged cultures. These compounds were identified as (i) S-coriolic acid, (ii) linoleic acid, (iii) p-anisaldehyde, (iv) p-anisyl alcohol, (v) 1-(4-methoxyphyenyl)-1,2-propanediol, and (vi) 2-hydroxy-(4′-methoxy)-propiophenone. The highest nematicidal activity was recorded with S-coriolic acid and linoleic acid with 5–10 ppm as a mean lethal dose (LD50). 62
The compounds obtained in the present study represent a potential tool for the control of sheep gastrointestinal nematodes.
Conclusion
This study shows evidence of compounds with a high EHI (>96%) against eggs of H. contortus using E1 fractions of P. djamor. E1 and E2 fractions possess compounds with a high larvicidal activity (>90%). In contrast, E3 fraction does not show nematicidal activity against either eggs or larvae. The results obtained with the fruiting bodies offer new possibilities in the use of edible mushroom industry with possible antiparasitic activity.
Footnotes
Acknowledgment
This research received financial support from two projects: 9442232005 and 834432984 (Proyectos Fiscales Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias). Publication costs were partially supported by Red Temática Farmoquímicos from CONACYT. This study was part of the bachelor degree thesis performed by Jesús Antonio Pineda Alegría to obtain the grade of Engineer in Biotechnology by the Universidad Politécnica del Estado de Morelos, Mexico.
Author Disclosure Statement
No competing financial interests exist.