
Abstract
Salvia officinalis, which has a high phenolic acid and flavonoid content, is a powerful antioxidant and anti-inflammatory herb. Inflammation plays an important role in the pathophysiology of many diseases and could cause damage by means of oxidative stress. The aim of this study was to investigate the anti-inflammatory and antioxidant activity of S. officinalis formed lipopolysaccharide (LPS)-induced experimental inflammation model. Four- to five-month-old 42 female Wistar albino rats were divided into six groups. Three groups were administered intraperitoneally 1 mg/kg LPS. Twenty-four hours after injection of LPS, 10 and 30 mg/kg S. officinalis extract were given orally to treatment groups. Pulmonary and hepatic 18F-fluoro-deoxy-D-glucose (18F-FDG) uptake was calculated to determine the status of inflammation by 18F-fluoro-deoxy-D-glucose-positron emission tomography (18FDG-PET) scan. Antioxidant enzyme activities and nitric oxide (NO) and malondialdehyde (MDA) levels were determined. Nuclear factor-kappa B (NF-κB) and tumor necrosis factor-alpha (TNF-α) levels were also detected in serum. As a result, lung and liver 18F-FDG uptake was found to be higher in the inflammation group than control group. MDA levels in erythrocyte and all tissue samples (liver, lung, and kidney) were found to be significantly higher compared to treatment groups. Superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase activities of the inflammation group in the liver, lung, kidney tissues, and erythrocyte SOD and CAT activities were determined to significantly lower than groups treated with S. officinalis. Increased NO, NF-κB, and TNF-α levels were found in the inflammation group. S. officinalis has been observed to have useful effects on LPS-induced inflammation and oxidative stress in rats.
Introduction
I
Many inflammation models have been formed by the action of bacterial endotoxins. Lipopolysaccharide (LPS) is an endotoxin that is an important component of the cell wall of gram-negative bacteria. 1 LPS administration induces receptors in monocytes and macrophage cell membranes and initiates a series of reactions. The stimulated receptors provide for the activation of transcription nuclear factor-kappa B (NF-κB) and protein kinases. Thus, the release of inflammatory mediators and free oxygen radicals from plasma, leukocytes, and vascular cells results in a response to inflammatory agents. 3,4
Cytokines that are polypeptide constructs secreted by activated macrophages and helper T cells (CD4+) are stimulated by bacterial products, toxins, physical agents, and the immune system. 5 The interleukins (especially interleukin [IL]-1, IL-6, and IL-8) and tumor necrosis factor-alpha (TNF-α) are important cytokines that are effective in the inflammatory process. TNF-α has an important role in regulating cell growth and proliferation, as well as proinflammatory effects. On the other hand, IL-1 has been shown to play a role in the synthesis of nitric oxide (NO) with prostaglandin as it activates cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS). 6,7
At the molecular level, free radicals and aldehydes (such as malondialdehyde [MDA]), formed during inflammation, cause DNA damage and posttranslational modifications of some proteins related to many diseases such as cancer, atherosclerosis, diabetes, arthritis, and osteoporosis. 1,8
The harmful effects of free oxygen radicals are reduced or eliminated by antioxidants. Enzymes involved in the redox cycle, such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx), are endogenous enzymatic antioxidants and called elements of the primary antioxidant defense system. 9,10
The ability to scavenge free radicals of Salvia officinalis (Lamiaceae), which has a high flavonoid and phenolic content, has been shown in various investigations. Studies have reported that S. officinalis eliminates free radicals in vitro and inhibits the development of cancer cells. The data obtained from studies indicate that S. officinalis is an important antioxidant and anti-inflammatory agent because it repairs DNA damage caused by free radicals and acts as a barrier to chain reactions leading to lipid peroxidation. 11,12
In our study, we investigated the effects of S. officinalis extract on inflammation and antioxidant system in an experimental inflammatory animal model induced by LPS. The results of the study are important for prevention of the inflammatory process, which plays an important role in the pathogenesis of cancer and many other diseases, use of new natural compounds in therapy, and for further studies.
Materials and Methods
Animals
Four- to five-month-old Wistar albino rats weighing 250–300 g were used in this study. The rats were housed in a room, which had a 12-h dark/12-h light cycle, temperature of 22 ± 2°C, and humidity of 45–50%, and fed on a standard pellet laboratory diet. All experimental procedures were approved by the Eskisehir Osmangazi University Medical Faculty Animal Use and Care Committee (Date: February 18, 2015, No: 79/402-2).
Study design and treatment
In the study, the inflammation model induced by LPS that was previously described was used. 13 Single dose of LPS (Escherichia coli, serotype 055-B5, 1 mg/kg) was intraperitoneally administered to the inflammation groups. Salvia officinalis (Bellasoft)® liquid extract of aerial parts was given to treatment groups orally (10 and 30 mg/kg) every 8 h for 2 days after 24 h of intraperitoneal LPS injection. Applied substances to control and treatment groups are shown in Table 1.
ig, intragastric; ip, intraperitoneal; LPS, lipopolysaccharide; SD, standard diet.
18F-fluoro-deoxy-D-glucose-positron emission tomography scanning
Rats were anesthetized with ketasol (1 mL/kg) to determine 18F-FDG uptake in the liver and lung tissues by 18F-fluoro-deoxy-D-glucose-positron emission tomography (18FDG-PET) scanning. 0.8 mL/kg 18F-fluoro-deoxy-D-glucose (18F-FDG) was injected intracardially into the rats. 18F-FDG uptake was determined 1 h after 18FDG injection using a PET scanner (Biograph 6 Hires PET/BT Siemens, Knoxville, TN).
Sample preparations
Blood samples were taken into tubes containing EDTA to prepare erythrocyte hemolysates. Blood samples were centrifuged at 1500 g for 5 min and the plasma was separated. Erythrocytes were washed three times with saline and erythrocyte packets prepared. Erythrocyte hemolysates were maintained at −80°C until MDA levels, and SOD and CAT enzyme activities were determined. Also, blood serum was separated for determination of TNF-α and NF-κB levels. After the animals were sacrificed, lung, liver, and kidney organ samples were removed and homogenized in potassium chloride (1%) with an ultrasonic homogenizer to determine SOD activies and MDA levels. To prepare the homogenates to be used in CAT and GPx measurements, homogenate preparation steps indicated in the CAYMAN enzyme determination kit were followed (CAYMAN CAT Assay Kit No: 707002, CAYMAN Gpx Assay Kit No: 703102). Erythrocyte hemoglobin levels were determined according to the oxyhemoglobin method and tissue total protein amounts were calculated according to the Sigma, Total Protein Kit (Product code: TP0100).
Determination of SOD activity
SOD activity was determined with the Sigma SOD Determination Kit (Cat. No. 19160) based on the WST (water-soluble tetrazolium salt) reaction. The calculated SOD activities were expressed as % inhibition. 14
Determination of CAT activity
Small-molecular-weight alcohols serve as electron donors for the peroxidative activity of CAT. The CAT activity was determined by the “CAYMAN CAT Assay Kit (707002)” based on the reaction of the enzyme with methanol at the appropriate hydrogen peroxide (H2O2) concentration.
Determination of GPx activity
While GPx shows its activity, it reduces hydroperoxides using glutathione (GSH) as electron donor. Oxidized glutathione (GSSG) is converted to the reactive GSH, which is catalyzed by the glutathione reductase enzyme. During the reduction of GSSG, nicotinamide adenine dinucleotide phosphate (NADPH) is also oxidized to oxidized nicotinamide adenine dinucleotide phosphate (NADP+). GPx activity was measured by the “CAYMAN Glutathione Peroxidase Assay Kit (703102),” based on the relationship between GPX activity and oxidation of NADPH to NADP+.
Determination of MDA levels
The method based on the color reaction of MDA, one of the lipid peroxidation products, with thiobarbituric acid (TBA) was used. 0.1 mL of homogenate, 3 mL of 1% phosphoric acid, 0.5 mL of distilled water, and 1.0 mL of 0.6% 2-TBA were mixtured and boiled for 45 min. After addition of 4.0 mL of n-butanol/piridin, homogenate and hemolysate MDA levels were measured spectrophotometrically at 532 nm. 15
Determination of NO levels
After homogenization of tissues in phosphate buffer, 0.5 mL of homogenate, 2 mL of NaOH, and 2.5 mL of ZnSO4 were added for deproteinization and left for 10 min. Copper-coated cadmium granules with 1 mL glycine buffer, 1 mL deproteinized sample, and 2 mL deionized water were incubated in room temperature for 90 min. 2.5 mL of deproteinized sample, 1 mL of water, 1 mL of sulfanilamide solution, and 1 mL of N-naphthylethylenediamine solution were added after 1 h of incubation. Absorbance was determined at 545 nm. The nitrite concentrations were calculated as μmol/mg protein for the tissue samples, considering the dilution factor that was constructed by using sodium nitrite as a standard. 16
Determination of TNF-α and NF-κB levels
TNF-α and NF-κB levels were determined according to the protocol “Yh Biosearch Laboratory ELISA Kit Cat. No. YHB0794Ra and Cat. No: YHB1098Ra.”
Statistical analysis
Data analysis was used with IBM SPSS 21. Normality of quantitative variables was evaluated with the Shapiro–Wilk test. Descriptive statistics of quantitative variables are given as mean ± standard deviation. One-way analysis of variance was used for overall comparison of groups, while pairwise comparisons were made with Tukey's test. Results that were obtained as P < .05 were found to be statistically significant.
Results
Effect on 18F-FDG uptake
Results in Tables 2 and 3 show that there were significant differences between 18F-FDG uptake of inflammation groups compared with control groups in the liver and lung homogenates, whereas S. officinalis treatment decreased standard uptake value (SUV) maximum levels (Fig. 1).
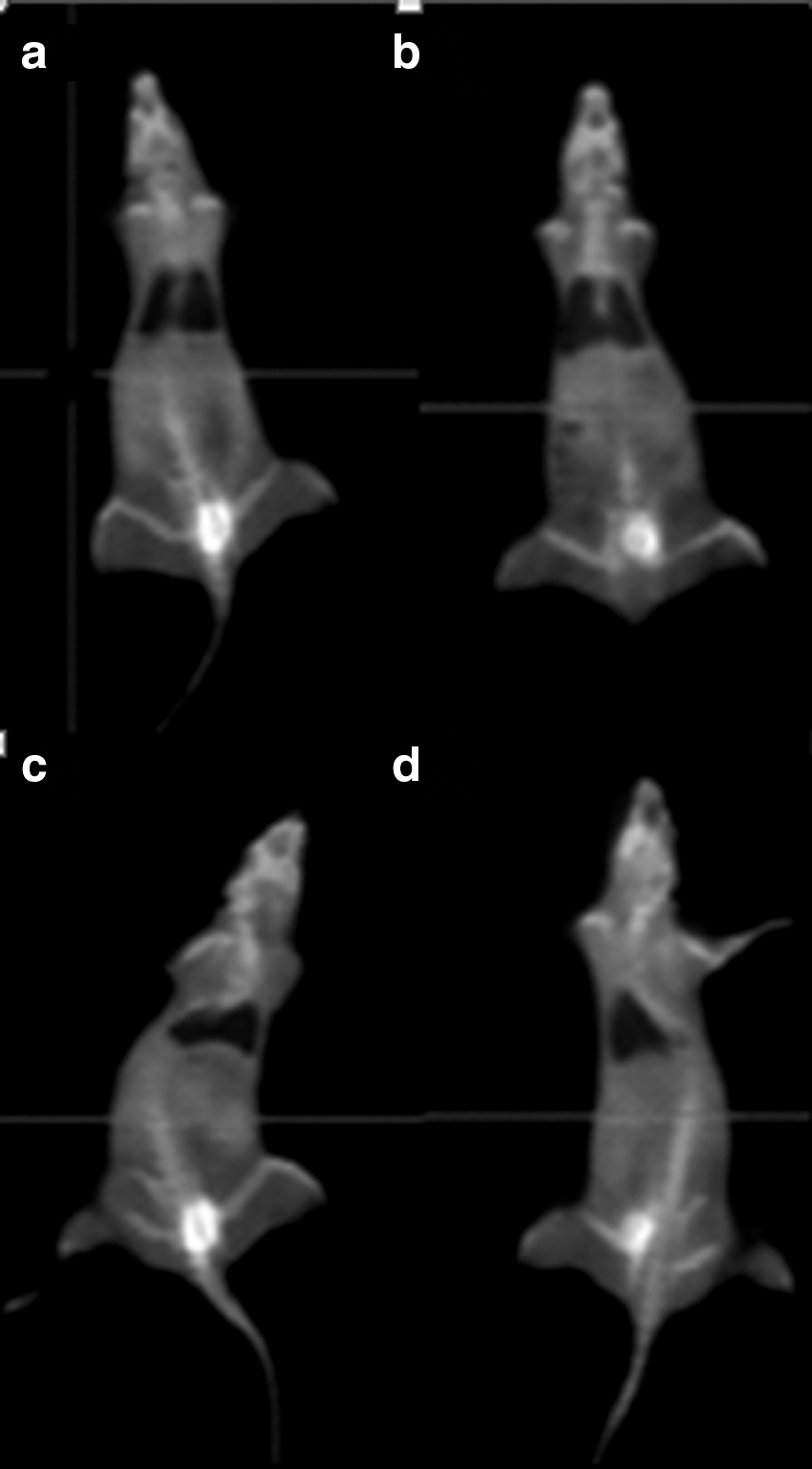
18FDG-PET scanning of
Data are presented as mean ± standard deviation (n = 7).
Significant differences were found: *** P < .001 compared with control group.
F, test statistics of ANOVA; ns, nonsignificant; SUV, standard uptake value.
Data are presented as mean ± standard deviation (n = 7).
Significant differences were found: * P < .05, *** P < .001 compared with control group.
Effects on MDA levels
MDA levels of inflammation groups in erythrocyte hemolysate and all tissue samples were found to be significantly higher than control groups. Besides treatment groups (inflammation + S. officinalis 10 and 30 mg/kg) MDA levels decreased compared to inflammation groups in all samples (Table 4).
Data are presented as mean ± standard deviation (n = 7).
Significant differences were found: ** P < .01, *** P < .001 compared with control group; # P < .05, ## P < .01, ### P < .001 compared with inflammation group.
MDA, malondialdehyde.
Effects on SOD activies
As shown in Table 5, SOD activities of inflammation groups decreased significantly compared to control groups in erythrocyte hemolysate and all tissue samples. In addition, S. officinalis treatment significantly increased SOD activities in hemolysate and lung and liver homogenates. S. officinalis treatment reversed LPS-induced decrease in SOD activites near control. Results demonstrated a significant effect of S. officinalis on increasing SOD levels in hemolysate and lung and liver tissues.
Data are presented as mean ± standard deviation (n = 7).
Significant differences were found: * P < .05, ** P < .01, *** P < .001 compared with control group; ## P < .01, ### P < .001 compared with inflammation group.
SOD, superoxide dismutase.
Effects on CAT activities
LPS administration significantly reduced CAT activity compared to control groups in the kidney, lung tissues, and hemolysate. These values were reversed by treatment with S. officinalis (inflammation + S. officinalis 10 and 30 mg/kg). On the contrary, we found that CAT activities of S. officinalis 10 and 30 mg/kg groups were significantly higher than that of inflammation groups in all tissue samples (Table 6).
Data are presented as mean ± standard deviation (n = 7).
Significant differences were found: * P < .05, ** P < .01, *** P < .001 compared with control group; # P < .05, ## P < .01, ### P < .001 compared with inflammation group.
CAT, catalase.
Effects on GPx activies
Similar to SOD and CAT, GPx activities of inflammation groups were significantly lower compared with those of control groups and increased activities of treatment groups were observed in all tissue samples. In addition, GPx activities of S. officinalis 10 and 30 mg/kg groups were significantly higher than that of inflammation groups in all tissue samples (Table 7).
Data are presented as mean ± standard deviation (n = 7).
Significant differences were found: * P < .05, *** P < .001 compared with control group; ### P < .001 compared with inflammation group.
GPx, glutathione peroxidase.
Effects on NO, TNF-α, and NF-κB levels
In the liver and kidney, NO levels of inflammation groups were observed to be significantly higher compared with control groups. Besides we determined decreased NO levels of treatment groups (inflammation + S. officinalis 10 and 30 mg/kg) significantly in the kidney. In addition, 30 mg/kg S. officinalis treatment reduced NO levels in the liver (Table 8). An increase was observed when TNF-α and NF-κB levels of inflammation groups were compared with those in control groups, but this increase was not significant (Table 9).
Data are presented as mean ± standard deviation (n = 7).
Significant differences were found: * P < .05, *** P < .001 compared with control group; # P < .05, ## P < .01, ### P < .001 compared with inflammation group.
NO, nitric oxide.
Data are presented as mean ± standard deviation (n = 7).
NF-κB, nuclear factor-kappa B; TNF-α, tumor necrosis factor-alpha.
Discussion
LPS leads to initiation of an immunological response by providing stimulation of receptors such as CD14 and TLR-4 in the membrane of monocytes, lymphocytes, and especially macrophages. 17,18 Induction of these receptors activates the nuclear transcription factor NF-κB and the mitogen-activated protein kinase (MAPK) pathway, enabling activation of a number of signaling pathways within the cell. As a result, many proinflammatory cytokines such as TNF-α, IL-1β, IL-2, IL-8, and IL-6 are released and an increase in expression of iNOS occurs. 19 At the end of this inflammatory response, products such as prostaglandins, which are formed by COX-2, NO, and free oxygen radicals, are triggered. 1
Concentrating on plants that are thought to be medically useful in recent years has enabled them to discover many biochemical properties. Studies have shown that plants rich in flavonoids, polyphenols, and phenolic acids are powerful antioxidants. The idea that S. officinalis may also have free radical scavenging properties suggests that it can be used as an important antioxidant, antibacterial, antimutagenic, hypoglycemic, and anticarcinogenic agent. 20
Our study suggests that S. officinalis prevents the inflammation induced by LPS, probably through increasing the activity of antioxidant enzymes and regulating molecules in the inflammation pathway.
18F-FDG, a radioactive glucose analogue, is usually used in PET imaging. 18F-FDG is introduced into the cell via transport proteins (especially GLUT-1) in cell membrane and converted to glucose-6-phosphate by hexokinase. The rapid metabolism of glucose in cancer cells causes more accumulated 18F-FDG in these cells. 18F-FDG uptake is not only associated with increased glucose levels in cancer cells, in addition, 18F-FDG uptake has also been shown to increase in many infectious diseases associated with increased rate of glucose uptake in inflammatory cells. 21,22 According to our findings, 18F-FDG uptake of inflammation group significantly increased in lung and liver tissues compared to the control group. This result indicated increased inflammation in the liver and lung. Furthermore, 18F-FDG uptake of treatment groups in the lung and liver was significantly higher than the uptake of the control groups. Thus, S. officinalis has been shown to reduce the inflammatory response. In addition, the fact that inflammation + S. officinalis 30 mg/kg group had less 18F-FDG uptake than inflammation + S. officinalis 10 mg/kg group suggested that the 30 mg application was more effective in the lung.
MDA, a lipid peroxidation product, has been identified as an indicator of oxidative stress in a number of studies. 23,24 Alkan et al. showed that MDA, an oxidative stress marker induced by cyclophosphamide, was decreased with the treatment of S. officinalis. 20 Moreover, it was reported that different antioxidants reduced MDA levels and prevented harmful effects of oxidative stress. 24 In the present study, levels of MDA in the inflammation group were found to be significantly higher than the control group, indicating that LPS triggered oxidative stress in all tissue samples. It was observed that S. officinalis used in treatment groups decreased MDA levels and prevented membrane damage.
Antioxidants inhibit the damage caused by oxidation of cellular components due to reactive structures of free radicals. Enzymes involved in the redox cycle, such as SOD, CAT, and GPx, naturally form enzymatic antioxidants in the organism. 25 SOD is an important enzyme that catalyzes the conversion of reactive oxygen species (superoxide, H2O2, etc.) to molecular oxygen. It has also been noted that SOD is easily inactivated by lipid peroxides or reactive oxygen species. 26 CAT is an endogenous enzyme that catalyzes the conversion of H2O2 to water and molecular oxygen. 27
Several studies have reported that LPS administration significantly reduces SOD, CAT, and GPx enzyme activities, and induces oxidative stress in the lung, kidney, and liver. 1,28 As previous studies described, S. officinalis significantly increased activities of antioxidant enzymes such as SOD and CAT and lowered lipid peroxidation products. 20,29 We demonstrated that LPS triggered formation of free oxygen radicals, which adversely affected the antioxidant system and reduced the activity of endogenous antioxidants CAT, SOD, and GPx in the lung, kidney, liver, and erythrocyte. The high antioxidant capacity of S. officinalis increased endogenous antioxidant enzyme activities in treatment groups.
NO is a very stable gas in an oxygen-free environment. However, peroxynitrite, the product of superoxide and NO reaction, shows strong oxidant properties. Reactive nitrogen species react with lipids, DNA, thiols, amino acids, and metals, destroy enzyme functions, and disrupt membrane integrity. 30 LPS administration has been shown to stimulate iNOS and trigger NO production as a cause of inflammation. 1,30 In the present study, NO levels of inflammation groups increased in the liver and kidney significantly. Treatment with S. officinalis (inflammation +10 and 30 mg/kg) reduced levels of NO in the kidney, and decreased NO levels in S. officinalis treatment group (inflammation + S. officinalis 30 mg/kg) were also demonstrated in the liver markedly.
Current information shows that NF-κB is a transcription factor that controls cytokine production and plays an important role during inflammation. The NF-κB pathway is activated via TLR-4 receptors induced by LPS. The activation of NF-κB causes the release of many inflammatory cytokines such as TNF-α, and is a key inflammatory mediator for the spread of the inflammatory process. 31,32 In many studies using LPS-induced models, TNF-α and NF-κB levels have been shown to increase in association with inflammatory response. 33 In our study, we indicated that levels of serum TNF-α and NF-κB were increased in the inflammation group compared to the control group. However, this increase was not statistically significant. Likewise, S. officinalis application reduced serum TNF-α and NF-κB levels in treatment groups, yet these findings were not statistically significant either.
In conclusion, suggesting that the possible mechanism of the therapeutic effect of S. officinalis in the liver, kidney, lung tissues, and hemolysate versus LPS-induced inflammation and oxidative stress may be due to the inhibition of reactive lipid peroxidation and/or the consumption of antioxidants by scavenging reactive oxygen radicals. S. officinalis seems to be highly effective in the therapy of many inflammatory diseases, including cancer.
Footnotes
Acknowledgment
This study was funded by the Commission of Scientific Research Project Support (BAP) Eskisehir Osmangazi University (Turkey).
Author Disclosure Statement
No competing financial interests exist.