
Abstract
Seed oils from oleaginous plants are rich in fatty acids (FAs) that play important roles in the health of the consumers. Recent studies indicate that FA also can play an important role in communication and regulation of virulence in bacteria. Nevertheless, evidence demonstrating protection against bacterial infections mediated by their quorum sensing inhibition (QSI) activity is scarce. In this study, sunflower, chia, and amaranth oils, were assayed for their QSI capacity by inhibiting violacein production and alkaline exoprotease activity of Chromobacterium violaceum. In vitro assays revealed that the oils exhibited QSI activities, whereas in vivo they delayed death of mice inoculated intraperitoneally with the bacterium. Gas chromatography coupled with mass spectrometry analysis of the oils indicated the presence of saturated FA (SAFA) and unsaturated FA as main components. Through a structure–activity relationship study of free FAs, bactericidal effect was identified mainly for polyunsaturated FAs, whereas QSI activity was restricted to SAFA of chains 12–18 carbon atoms in length. These data correlate with a possible interaction suggested by molecular docking analysis of lauric, myristic, and stearic acids with the CviR protein. Our study highlights the antiquorum sensing potential of SAFA, which may be future antivirulence therapeutic agents for the treatment of bacterial infections.
Introduction
E
Indiscriminate use of antibiotics has produced an alarming increase in the number of resistant pathogenic bacteria. 5 Hence, emergence of resistance has become a threat and has been declared a public health problem worldwide. 6 However, the development of new antibiotics has been insufficient to cope with the dramatic increase of resistance selection. 7 Therefore, new approaches to combat bacterial infections are required 8 ; one of these is to primarily inhibit virulence rather than growth, for example by attenuating bacterial cell–cell communication known as quorum sensing (QS). 9 QS systems (QSS) are used by many microorganisms to modulate several processes, including the expression of virulence factors and biofilm formation. 10 –14
QS inhibition (QSI) has been proposed as an alternative strategy that aims to counteract bacterial resistance. 13,14 QSI, then, represents a promising therapeutic approach with lower risk of selection for development of resistance than treatments with traditional antimicrobials. 15
In this respect, the QSS dependence on N-acyl homoserine lactones are the main regulators of virulence factor expression and biofilm development in several Gram-negative pathogenic bacteria. 16 Hence, their inhibition represents a very important novel approach for the design of new therapies. 15,17
In this regard, some FA that have shown bactericidal efficacy, particularly those that are unsaturated and have medium-long chains exhibit the best activities. 18,19 In addition, it has recently been reported that certain food extracts from ground meat and poultry have FAs that are able to inhibit certain QSS. 20
Chromobacterium violaceum is an aquatic bacterium that can infect humans causing abscesses and bacteremia. 21 Although reported cases of human infection by this bacterium are considered low, relative to other bacteria of medical importance, C. violaceum infections have a major impact on public health due to their high mortality rate. 22 This microorganism activates QS through the C6-homoserine lactone (C6-HSL) autoinducer 23 to regulate violacein production and biofilm formation as well as chitinase and alkaline protease activity. In fact its violacein production is widely used as an indicator of the efficiency of molecules with potential QSI. 23,24
In this study, we analyzed the QSI activity of three seed oils, as well as some free FA, on the QSS of C. violaceum. Also, the effect of consuming seed oils on survival of mice infected with bacteria was determined.
Materials and Methods
Extracting seed oils and gas chromatography coupled with mass spectrometry analysis
The sunflower seeds (Helianthus annuus L.), INSHELL® sourced from Argentina, were purchased from Mexicana de Víveres MSV, S.A de C.V. “Black” chia seeds (Salvia hispanica L.) were obtained from a production center in San Mateo Coatepec, Atzitzihuacán, Puebla, and amaranth seeds (Amaranthus hypochondriacus L. cv. PQ2) were provided by Dr. Sergio Barrales Domínguez of the Universidad Autónoma Chapingo (UACh).
The seeds were pulverized in a blender (Hamilton Beach Mod. 600-6) and deposited in paper bags (Whatman No. 4). Samples were extracted in a Soxhlet system with 300 mL of hexane (JT Baker) for 8 h. The solvent was evaporated in a rotary evaporator (Buchi-R114, Switzerland) at 40°C, and samples were stored in amber bottles in a cool place.
Derivatization: for each 10 μL of sample, 1 mL 0.5 mol L−1 NaOH was added and allowed to react at 75°C for 30 min. The mixture was cooled at room temperature and 1 mL methanol boron trifluoride (14% methanolic solution) (Sigma-Aldrich) was added and allowed to react at 75°C for 20 min. Finally, 1 mL distilled water and 2 mL hexane (JT Baker) were added. The mixture was stirred vigorously and the organic phase was stored at 2°C. 25
Chromatographic separation was carried out using the Agilent 5975T low thermal mass/mass selective detector (LTM/MSD) system 26 equipped with a split/splitless injector autosampler. A fused silica capillary column HP-SMS (30 m × 0.25 mm i.d. and 0.25 μm film thickness) was used. Helium was used as the carrier gas. LTM oven: 70°C 1 min to 10°C/min to 200°C, 5°C 1 min to 280°C, 33 min. Electron energy was 70 eV. The results were expressed as the relative percentage area.
Fatty acids
Saturated fatty acids (SAFAs) (EC10A-1KT Supelco: caproic, caprylic, capric, lauric, myristic, palmitic, stearic, arachidic, behenic, and lignoceric acids, 73982 Fluka: pelargonic acid) and unsaturated fatty acids (UFAs) (UN10-1KT, Supelco: palmitoleic, petroselinic, elaidic, oleic, linoleic, linolenic, arachidonic, erucic, cervonic, and nervonic acids) were obtained from Sigma-Aldrich.
Determination of growth and violacein production
The effect of the oils and FA on the QS-controlled production of violacein was determined using the wild-type pigment-producing strain of C. violaceum 553, whereas the potential toxic effects were determined using the nonpigmented CV026 strain (mini-Tn5 mutant with the autoinducer synthase cviI gene interrupted) to avoid inaccuracy due to light scattering of violacein at 660 nm. 27 All bacterial strains were stored in 10% glycerol stocks at −70°C and later cultured in Luria-Bertani (LB) broth and incubated at 28°C.
For each assay a fresh overnight culture with an OD660 adjusted to 0.1 (∼1 × 105 cfu/mL) was used. The experiments were conducted in 96-well flat-bottom plastic microplates (Corning® Costar®). To each well 0.2 mL of culture was added, plus 5 μL oil (62.5, 125, 250, and 500 μg mL−1) or FA (0.001, 0.01, 0.1 and 1 mmol L−1) and incubated for 48 h. The vehicle dimethylsulfoxide (DMSO; Sigma-Aldrich) was used as negative control and 500 μg mL−1 of anacardic acid mixture (AAm) as a positive control. 27 Assays were performed in triplicate with three independent replicates per treatment.
Violacein extraction was achieved by taking 200 μL from each culture and adding the same amount of ethyl acetate. The mixture was then shaken at maximum speed by a vortex apparatus for 2 min and centrifuged at 6000 g for 1 min. The organic phase was recovered and absorbance was read with a spectrophotometer at a wavelength of 575 nm. To calculate the percentage of inhibition, absorbance of the controls was considered to be 100% production of violacein.
Exoprotease activity
The wild-type strain was cultured in 5 mL of LB medium at 28°C and 200 rpm shaking ( IKA® HS Shaker, Model KS 250) until the cells reached an OD600nm of ∼1.0; 100 μL of each oil or DMSO as negative control were then added and the cells were incubated for 24 h. After that, aliquots of 1.5 mL of each culture were taken, and the supernatants were obtained by centrifugation at 6000 g for 1 min, then 150 μL of each supernatant was mixed with 10 mg of hide remazol blue powder (Sigma-Aldrich) and suspended in 1 mL of buffer Tris 20 mmol L−1 CaCl2 1 mmol L−1 at pH of 8.0. The mixture was incubated at 37°C and 200 rpm shaking for 1 h and centrifuged at 6000 g for 5 min to remove the hide remazol blue powder that was not hydrolyzed. The blue pigment solubilized in the supernatants (proportional to the exoprotease activity) was quantified with spectrophotometry by recording absorbance at 595 nm. Activity was normalized by growth of the cultures (OD600nm). 28 Experiments were done with four independent cultures.
Molecular docking analysis
Crystal structure of CviR ligand-binding domain bound to the native ligand C6-HSL was obtained from the protein data bank (accession no. 3QP1). The three-dimensional models of the FA used in the study were generated and optimized using Argus Lab 4.0.1 software (
Animal infection model
Male CD1 mice (4–5 weeks old) were purchased from Facultad de Estudios Superiores, Iztacala-UNAM and were housed at Colegio de Postgraduados-Montecillo following the Official Mexican Norm (NOM-062-ZOO-1999) in compliance with technical specifications for production, care, and use of laboratory animals.
For the bacterial inoculum, 5 mL of 24-h cultures (28°C with shaking at 200 rpm) were adjusted to OD660nm = 0.5–0.6, centrifuged to 6000 g, and washed with phosphate-buffered saline (PBS). The bacterial suspension was diluted in saline solution and 0.3 mL was used for intraperitoneal (i.p.) inoculation of the mice. The cfu was determined by counting colonies in LB plates and the animals were observed daily for 2 days to evaluate survival.
Effect of oral administration of seed oils on survival of C. violaceum-infected mice
Groups of 10 animals were fasted for 6 h with free access to water. Two hours before bacterial inoculation (∼8.6 × 1010 cfu), mice were administered 0.2 mL of each oil orally through a metal cannula. As negative control, 0.2 mL of PBS was administered. Throughout the experimental phase (48 h), all groups were fed commercial pellets (Purina®). Three independent replicates were performed and survival was recorded every 2 h.
Statistical analysis
Statistical analysis was performed with SigmaPlot 10 (Systat Software, Inc., San José, CA, USA). Values shown represent the mean ± standard deviation. The assessment of statistical significance of differences between mean values was performed by analysis of variance and the Dunnett's test (Statistix 9, Analytical Software, Tallahassee, FL, USA) and two-tailed Student's t-test. The log-rank (Mantel–Cox) test was applied to Kaplan–Meier survival data to evaluate differences between groups.
Results
Seed oils showed QSI activity
To determine the inhibitory activity of seed oils on the QS of C. violaceum, violacein production and the activity of alkaline exoprotease were evaluated. Interestingly, the three oils showed QSI activity; they inhibited pigment production without affecting growth at concentrations of 250 and 500 μg mL−1. The magnitude of inhibition was similar to that shown by the positive control (AAm) at 500 μg mL−1 (Fig. 1). For inhibition of alkaline exoprotease, the effects of sunflower oil (SFO) and chia oil (CO) were also significant; concentrations of 2% inhibited 65% of the exoprotease activity (Fig. 2).
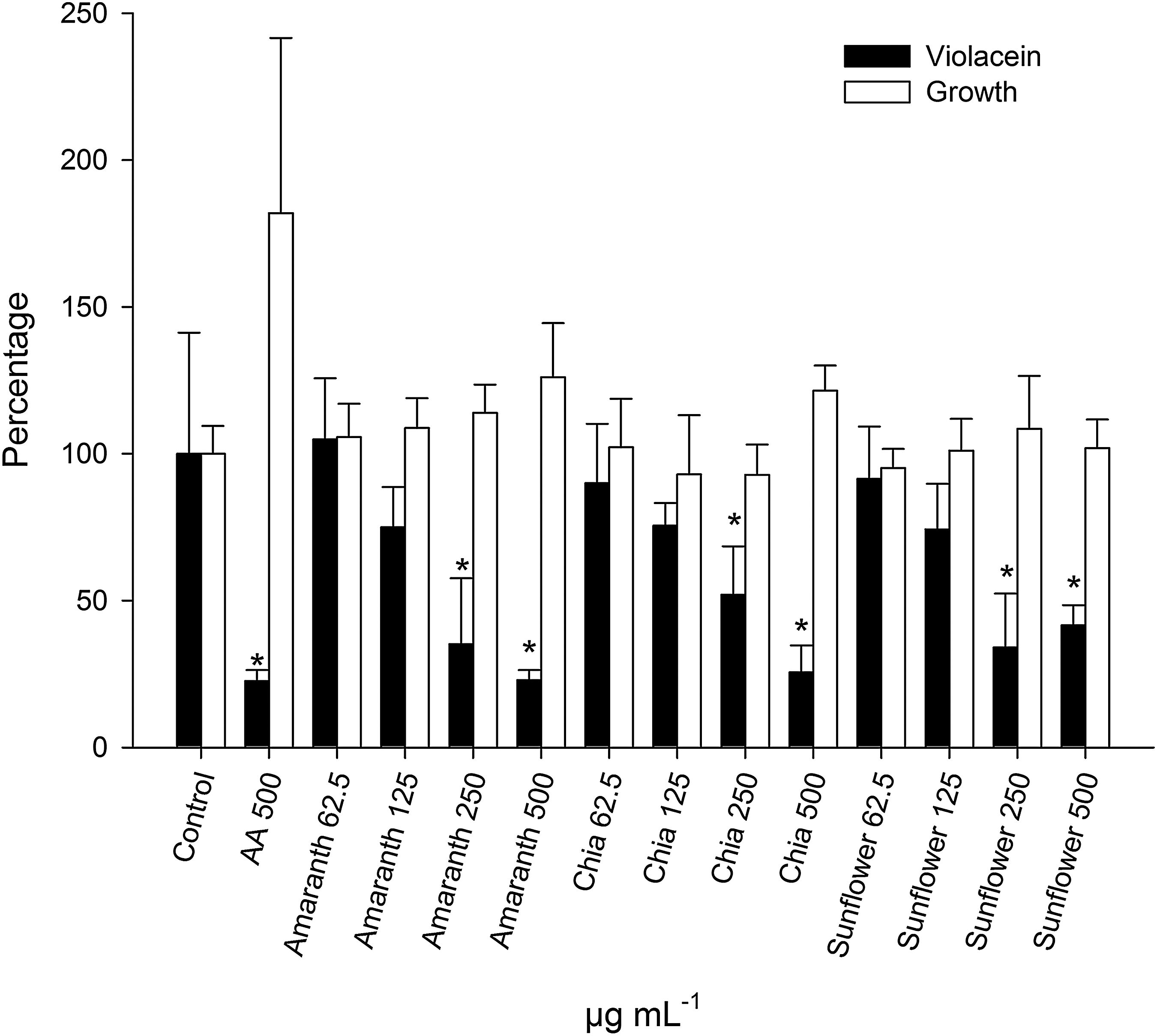
Effect of AO, CO, and SFO on violacein production (black bars) and growth (white bars) of Chromobacterium violaceum. Different concentrations of the oils diluted in DMSO (negative control) were evaluated. AAm was used as a positive control. *P < .05. AAm, anacardic acid mixture; AO, amaranth oil; CO, chia oil; DMSO, dimethylsulfoxide; SFO, sunflower oil.

Effect of AO, CO, and SFO on alkaline protease activity of C. violaceum. *P < .05, two-tailed Student's t-test.
FA composition of seed oils
To analyze the differences in composition of these FA in oils, methylated derivatives were obtained and analyzed by gas chromatography coupled with mass spectrometry (GC/MS). The three oils were similar in type of FA present, but there were differences in proportions (Table 1).
AO, amaranth oil; CO, chia oil; FA, fatty acid; MUFA, monounsaturated fatty acid; PUFA, polyunsaturated fatty acid; SAFA, saturated fatty acid; SFO, sunflower oil.
As can be observed, SFO had the highest percentage of UFA (83.3%); of these, oleic (40%) and linoleic (39%) were more abundant. The total amount of SAFAs was 19.7%; palmitic acid (15.1%) was more representative.
In the case of the amaranth oil (AO), the proportion of UFA was the lowest of the three samples with 55.8%; however, it was the oil with the highest amount of SAFA (44.14%), the main component being palmitic acid with 38.8%.
CO had intermediate concentrations of UFA (76.8%), SAFA (23.1%), and palmitic acid (19.1%) (Table 1). In the case of stearic acid, the proportion was similar in the three oils; however, myristic acid was identified only in SFO and AO.
SAFAs exhibit QSI activity
To determine the involvement of SAFA in the QSI activity exhibited by oils, its effect on violacein production and bacterial viability was assessed. The FA that showed QSI activity were lauric, myristic, and stearic acids, which have long aliphatic chain lengths of 12–18 carbon atoms (SAFA: 12, 14, 18). These inhibited pigment production by 60–84% at 1 mmol L−1, which was the highest concentration tested (Table 2). Palmitic acid showed a mild effect on violacein production, but it was not significant. Likewise, no QSI or bactericidal activity was recorded for the other UFA evaluated. Erucic acid was the only monounsaturated fatty acid (MUFA) with QSI activity, inhibiting violacein production by 47% without affecting growth (Table 2).
Length of the aliphatic chain: unsaturated.
P < .05.
—, no activity; nd, not determined.
Molecular docking analysis
To investigate whether QSI activity of FA was related to their binding and inhibition of the QS receptor CviR, molecular docking analysis was performed using the Autodock simulation program. Conformations of the CviR-FA complex selected corresponded to the lowest values of dissociation constants (Kd) (Table 2). As a reference model, the natural autoinducer C6-HSL was docked in the ligand-binding domain of CviR (Fig. 3). The results of molecular docking analysis suggested that the QSI activity exhibited by seed oils could be related to the binding of SAFA of 12–18 long chains of carbon to CviR.
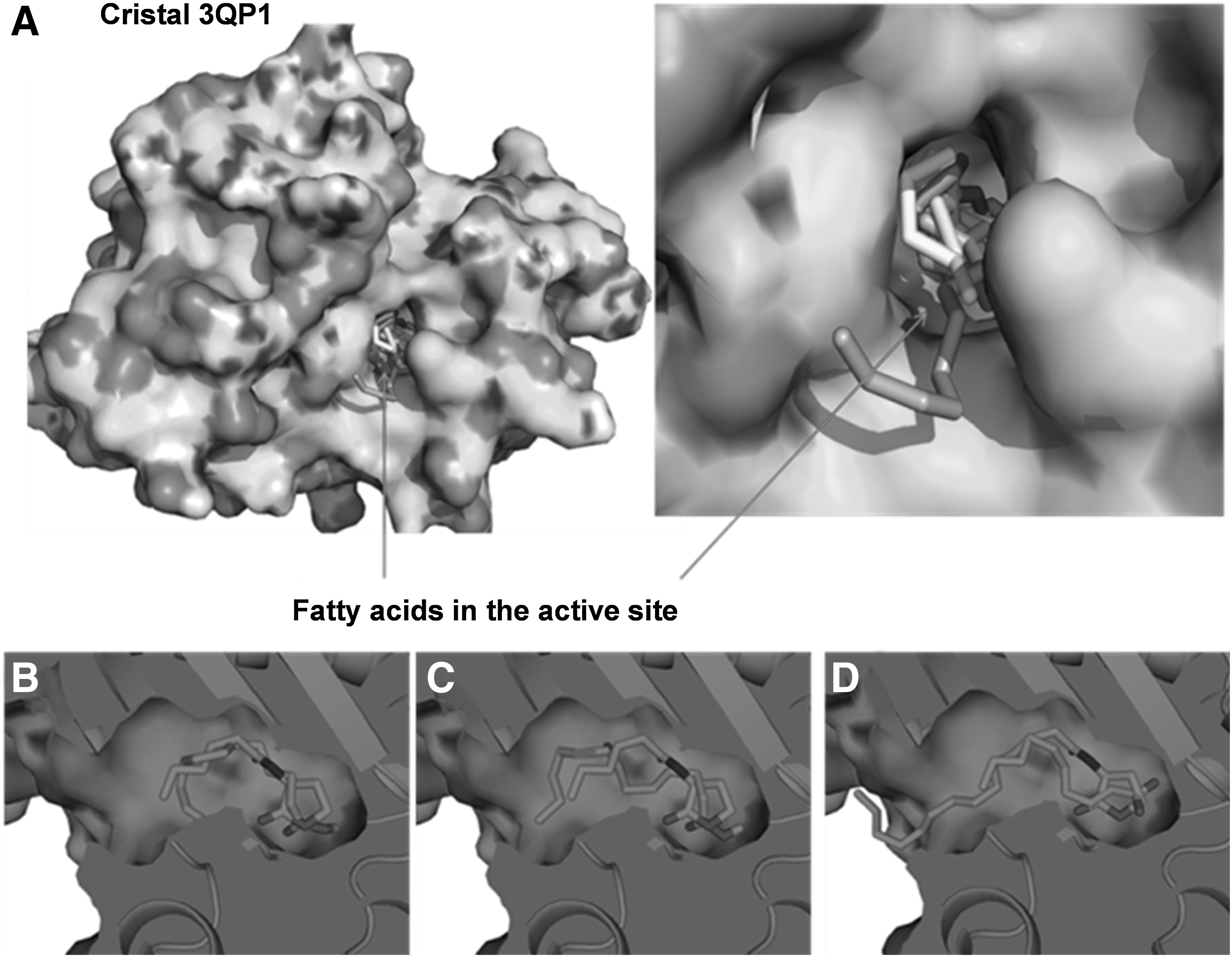
The docking analysis of SAFA 12–18 in the CviR active site rendered Kd values of 251.74–879.72 μmol L−1 (Fig. 3 and Table 2). Of these, lauric, myristic, and stearic acids had the lowest Kd values of the most favored conformation, indicating a higher binding affinity to the active site of the CviR protein. In contrast, lignoceric acid (with a longer hydrophobic chain) and caproic acid (with a shorter hydrophobic chain) did not show stable arrangement inside the active site of the protein (Table 2). In addition, MUFA (elaidic, oleic and erucic acids) were docked in the active site with Kd values in the range of 1060–8390 μmol L−1 (Table 2).
UFAs exhibit antibacterial activity
The MUFA and PUFA that showed significant bactericidal activity were those that have 18–22 carbon chain lengths, such as oleic, linoleic, linolenic, arachidonic, and cervonic acids (Table 2). Interestingly, their antibacterial activity was directly proportional to the degree of unsaturation; cervonic was the most active inhibiting growth by 92.5% (Table 2).
Oral administration of oil delays death of mice infected with C. violaceum
Survival of the mice was analyzed by Kaplan–Meier curves (Fig. 4). The results of the survival analysis were expressed through median survival (MS), which is defined as time elapsed, until death of 50% of mice was observed.
Effect of oral administration of PBS (), AO ( ), CO (
), CO ( ), and SFO (
), and SFO ( ) on the survival of mice after intraperitoneal challenge with C. violaceum (∼8.6 × 1010 cfu/mice). The data correspond to the averages of three independent experiments, with n = 10. The animals were fasted for 6 h. Two hours before the infection, 0.2 mL of oil was administered orally. Significance of difference of cumulative survival rates was calculated using the Kaplan–Meier method, with a log–rank test (Mantel–Cox test). P < .01. PBS, phosphate-buffered saline.
) on the survival of mice after intraperitoneal challenge with C. violaceum (∼8.6 × 1010 cfu/mice). The data correspond to the averages of three independent experiments, with n = 10. The animals were fasted for 6 h. Two hours before the infection, 0.2 mL of oil was administered orally. Significance of difference of cumulative survival rates was calculated using the Kaplan–Meier method, with a log–rank test (Mantel–Cox test). P < .01. PBS, phosphate-buffered saline.
The SFO-treated group had the highest MS (18 h, 15.55–20.45), compared with the groups treated with CO (16 h, 14.29–17.72), AO (14 h, 12.67–15.33), and PBS (10 h, 8.63–11.37). Only significant differences (determined by Log-Rank Test [Mantel–Cox ]) in survival curves were found between the groups administered with oil and treated with PBS, confirming that the oils provide protection against C. violaceum infection.
However, in this assay all infected animals (regardless of the experimental condition) showed reduced movement and did not consume any food or water. Hence, to avoid unnecessary suffering of animals, the tests were ended after a maximum of 2 days (Fig. 4).
Discussion
In this study, QSI activity of seed oils from oleaginous plants against C. violaceum bacteria was identified by in vitro assays. Our results indicate that QSI activity was restricted to lauric, myristic, and stearic acids (SAFA: 12–18). Previous reports suggest the potential of FA in regulating (inhibition/activation) QS in certain bacterial systems. In this regard, Abd-Alla and Bashandy reported that production of myristic and linolenic acids in the onion bulb was a response to infection by Pseudomonas aeruginosa. 34 The authors suggested that myristic acid acts as part of the plant's defense system and this may be related to its QSI activity, because myristic acid in vitro inhibited pyocyanin production and exoprotease, lipase, and polygalacturonase activity. 34 Also, it has been reported that FA present in extracts or fractions obtained from ground meat and poultry meat, exhibit QSI activity on the AI-2 system of Vibrio harveyi and counteracts biofilm formation in Escherichia coli K-12. 20,35 The authors suggest that oleic, linoleic, palmitic, and stearic acids are involved in these QSI effects.
In our work, molecular docking analysis correlated well with the QSI activity found by in vitro assays, suggesting a structural relationship between the sizes of the SAFA: 12–18 and the CviR receptor of C. violaceum. However, for the case of palmitic acid, despite having a low Kd value and a chain of 16 carbon atoms, the QSI activity recorded in the in vitro assays was not significant and was the only one that did not show a dose–response effect on QSI (data not shown).
Moreover, the bactericidal activity on C. violaceum was identified only in the UFA (oleic, linoleic, linolenic, arachidonic, and cervonic acids), whose efficiency is directly proportional to the degree of unsaturation. In general, UFAs with a higher degree of desaturation tend to be more effective as antibacterials. 36 In their free form, UFAs can impair cell membrane properties, inhibit enzyme activity, and promote bacterial cell damage through auto-oxidation or peroxidation products. 37 Similarly, our molecular docking analysis is correlated with their in vitro activity, in which FA with unsaturated chains do not exhibit good affinity for the receptor CviR. However, it has been reported that linoleic, oleic, stearic, and palmitic acids from other sources may have bactericidal activity against Staphylococcus aureus, Klebsiella pneumoniae, and species of the genus Vibrio. 18,19
Moreover, the oleic acid produced by rhizosphere bacteria (Stenotrophomonas maltophilia BJ01) exhibit QSI activity in C. violaceum and antibiofilm activity in P. aeruginosa. 38 In contrast, in our assays significant inhibition of C. violaceum growth by oleic acid (22.9%) was recorded.
FA protection from bacterial infections has been reported in other animal models. Using Daphnia magna an increase in survival and reproduction due to arachidonic acid (ARA) intake during Pseudomonas spp. infection was registered. 38 However, the authors indicate that the mechanism is not due to antimicrobial activity and suggest that it may be because of ARA-derived eicosanoids, which are involved in the defense mechanisms of the crustacean. 39 Similarly, the cis-2-dodecenoic acid factor signal of Burkholderia cenocepacia (BDSF) increases survival of zebrafish infected by P. aeruginosa. The authors attribute the protective effect to virulence attenuation by QSI and to interference with type III secretion system. 40
In C. violaceum, the production of violacein pigment and virulence factors, such as chitinase 41 and alkaline exoprotease activity, are regulated positively by QS. 24 It also increases formation of biofilm, swarming, H2O2 stress tolerance, 26 and controls expression of type VI secretion system. 42 However, it should be noted that, in this study, the role of QSI in delaying animal death is not clear since the infective capacity of the CV026 mutant strain does not decrease (data not shown). It is known that i.p. infection of C. violaceum induces fulminant hepatitis in mice through the T3SS encoded in Cpi-1/-1a. 43 It was also observed that mutant strains lacking the Cpi-1/-1a-encoded T3SS do not produce death in infected mice. 43 In this regard, salts of oleic, myristic, and palmitic acids have recently been reported to repress transcription of hilA, a transcriptional activator of the T3SS structural genes in Salmonella enterica serovar Typhimurium. 44 Although in vitro studies indicate that the QSI and bactericidal activities are attributed to certain types of FA, it is not yet clear how they participate in the protective effect observed in our in vivo studies. We suggest that protection by seed oils during pathogen challenge may be due, in part, to a combination of antimicrobial effects of FA (QSI and bactericide activity). In this respect, it has been recently reported that the combination of FA and antibiotics potentiates the susceptibility of bacteria, increasing the efficiency of antibiotics against several bacterial genera. 40
In conclusion, this study complements previous work related to the beneficial effects of the ingesting FA and the protection they provide against bacterial infections. Also, our study suggests that this effect may be the result of a combination of FA with QSI and bactericidal activities, providing a starting point for the development of new strategies to protect animals and humans against acute bacterial infections.
Footnotes
Acknowledgments
This work was supported by grants from Scientific Development Projects for Solving National Problems/CONACyT Mexico No. 2015-01-402. M.P.-L. thanks CONACyT for her master scholarship. I.C.-J. was supported by Fideicomiso-COLPOS 167304 and Cátedras-CONACyT program. The authors thank E. Ibarra-Estrada for GS-MS analysis. R.G.-C's research funding was by the PAPIIT UNAM RA201116 Grant. J.S.R.-Z's research funding was by the SEP/CONACyT-México Grant No. 166463. Patent application in process: IMPI: MX/E/2015/007607-MX/E/2015/042763.
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationship that could be construed as a potential conflict of interest.