
Abstract
Obesity is associated with elevated plasma levels of glucocorticoids and reduced levels of thyroid hormones, both known to effect food intake and energy expenditure. Furthermore, tissue specific glucocorticoid metabolism is altered in obesity, increasing insulin resistance and cardiometabolic risk. The goal of this study was to examine whether these metabolic disturbances can be prevented with the isoflavone genistein in the ob/ob mouse, a model that resembles the phenotype in human obesity. Male ob/ob mice, aged 5 weeks, were fed either a genistein-rich diet (600 mg/kg) or a genistein-free diet for 4 weeks. ob/ob mice weighed 70% more than lean controls. While there was no effect of genistein on body weight, food consumption during weeks 3 and 4 was significantly increased in genistein-fed mice. This was associated with increases in body temperature and plasma levels of triiodothyronine (T3), suggesting a thermogenic effect. The hypercorticosteronism observed in the ob/ob mouse was reduced with genistein treatment. This effect was accompanied by a decrease in protein expression of renal 11β-hydroxysteroid dehydrogenase type 2 (11β-HSD2) without changes in hepatic 11β-HSD1. Our results suggest that a diet containing genistein can have beneficial effects on energy expenditure, T3 production, and corticosterone status in the ob/ob mouse model of obesity.
Introduction
T
One approach to healthful eating is to increase the consumption of soy-based foods. Recent evidence indicates that consuming the isoflavone phytoestrogen genistein, abundantly found in soy and other plants, 5 reduces the risk of type 2 diabetes. 6,7 The surrogate endpoints associated with cardiovascular disease, such as fasting insulin, glucose, and blood pressure, are reduced in postmenopausal women on a genistein-rich diet. 8 Structurally, genistein is similar to 17-β estradiol and exhibits similar binding affinities to estrogen receptors, 9 hence its cardioprotective effects. Genistein exerts antidiabetic effects in animal models of diabetes, 10 and we have previously shown that a genistein-rich diet improved gut function and bone structural properties in the ob/ob mice, as well as ischemic tolerance of hearts from ovariectomized rats. 11,12 These findings not only support the benefits of bioactive compounds displaying antiobesity activity but also highlight the benefits of increasing the consumption of leguminous plant-based foods that contain genistein.
The leptin-deficient ob/ob mouse is commonly used to investigate the effects of obesity and type 2 diabetes because of its close phenotypic resemblance to the human condition. 13 These mice are homozygous for the obese spontaneous mutation (Lepob, commonly referred to as ob or ob/ob). Leptin is an adipocyte-derived peptide that exerts a key role in the regulation of energy intake and satiety through central hypothalamic mechanisms, as well as peripheral actions on energy expenditure. 14 In the ob/ob mouse, failure to regulate food intake occurs from leptin deficiency, and the expected sequelae of hypoleptinemia include hyperphagia followed by insulin resistance leading to decreased energy expenditure and severe obesity. 15 Earlier studies have indicated that the decreased energy expenditure in the ob/ob mouse is linked to both low thyroid hormone production and increased endogenous glucocorticoid production from dysregulated hepatic and renal metabolism of corticosterone, 8,9,10 with the latter being a risk factor associated with insulin resistance and hyperglycemia. 16
The effects of genistein on energy expenditure and the obese state in the ob/ob model are poorly understood. With the observation that genistein improves glucose and insulin metabolism in the nonobese diabetic mouse, 17 it increases food intake in chemically induced diabetes, as well as AMP-activated protein kinase and peroxisome proliferator-activated gamma coactivator expression in healthy mice, 18,19 suggesting that energy expenditure is enhanced. Since little research to date has focused on the role of genistein in the ob/ob mouse, we examined the effects of short-term genistein treatment on the thyroid and corticosterone status, food intake, and body temperature.
Materials and Methods
Mouse model of obesity and treatment protocol
All animals were cared for in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals (National Institute of Health, publication number 85-23, 1986) and approved by the Midwestern University Institutional Animal Care and Use Committee. Five-week-old male ob/ob (C57BL/6J background strain; B6.Cg-Lepob/J) male mice were purchased from Jackson Laboratory (Bar Harbor, ME, USA). This strain contains the spontaneous mutation of the Lepob gene, and homozygous mice exhibit early hyperphagia, followed by obesity and insulin resistance as a result of leptin deficiency. This mouse model is also characterized by decreased body temperature and low levels of thyroid hormones. 20 The lean littermates, which possess only one spontaneous deletion of the Lepob gene, served as controls. After 1 week of acclimatization, ob/ob mice were randomly assigned to a genistein-enriched diet (600 mg genistein/kg diet, 600 g; Dyets, Inc., Bethlehem, PA, USA) or a genistein-free diet (0 mg genistein/kg diet, 0 g) for a period of 4 weeks. This diet was selected because it is effective in producing a significant increase in plasma levels of genistein. 21 A description of the diet is found in Table 1. Mice were provided with food and water ad libitum in a room with an alternating 12-h light/12-h dark cycle.
Food intake, measurement of body temperature, and blood sampling
Mice from each group were placed in individual cages for weekly measurement of food intake over a 24-h period. Body temperature using rectal temperatures (Temp JKT; Oakton Instruments, Vernon Hills, IL, USA) was measured and recorded at week 0 (start of the diet) and at weeks 2 and 4. After treatment, mice were euthanized in the morning between 9 and 11 am by asphyxiation in an atmosphere of 100% CO2 followed immediately by surgical thoracotomy (inducing pneumothorax). Blood samples were obtained by cardiac puncture, and the plasma was separated by centrifugation and then stored at −80°C for subsequent measurement of insulin (ALPCO Diagnostics, Salem, NH, USA), corticosterone (Cayman Chemical Company, Ann Arbor, MI, USA), and triiodothyronine (T3) levels (Thermo Fisher Scientific, Inc., Waltham, MA, USA) using standard enzyme-linked immunosorbent assay techniques as described by the manufacturer.
Western blot analysis
Following euthanasia, kidney and liver tissues were immediately snap frozen in liquid nitrogen and stored at −80°C for western blot analysis as previously described. 22 Samples were analyzed for protein content, and run on NuPAGE® 4–12% Bis-Tris gels (Thermo Fisher Scientific, Inc.) at 150 V for 2 h. Transfer was carried out for 2 h at 30 V at 4°C. The blots were incubated with primary antibody 11β-hydroxysteroid dehydrogenase type 1 (anti-HSD11β1 antibody, 1:1000; Abcam Cambridge, MA, USA), 11β-hydroxysteroid dehydrogenase type 2 (anti-HSDIIβ2 antibody, 1:800; Abcam), and GR (1:1000, Glucocorticoid Receptor D6H2L; Cell Signaling Technology, Danvers, MA, USA) overnight at 4°C in 5% milk in Tris-buffered saline solution +0.1% Tween (TBST). To probe for actin (internal control), blots were then incubated with anti-actin primary antibody (1:5000; EMD Millipore, Billerica, MA, USA) for 1 h at room temperature. After washing, blots were incubated with secondary antibody, anti-rabbit immunoglobulin (Ig)G (H+L) DyLight™ 800 Conjugate (1:15,000; Thermo Fisher Scientific, Inc.), for 1 h at room temperature. Images of membranes were obtained, and all proteins of interest were normalized to actin. Band density was analyzed using Odyssey-CLx (LI-COR Biosciences, Inc., Lincoln, NE, USA) and Image Studio Software (LI-COR Biosciences, Inc.).
Statistical analysis
Data are presented as mean ± standard error of the mean (SEM). Group mean differences were determined using analysis of variance followed by a Tukey–Kramer comparison for post hoc analysis. Analyses were performed using GraphPad Software (GraphPad Software, Inc., La Jolla, CA, USA). A value of P < .05 was considered statistically significant.
Results
Effects of genistein on physical characteristics
The effects of genistein treatment on the physical characteristics of ob/ob mice are shown in Table 2. As expected, body weight was significantly increased in ob/ob mice compared to lean mice (49.05 ± 1.13 g vs. 28.37 ± 1.24 g, n = 10, P < .05). The increased body weight of ob/ob mice was primarily associated with significantly higher fat pad and liver weights, since no differences in heart, brain, and spleen were noted between the ob/ob mice and lean mice. ob/ob mice were also insulin resistant, reflected by increased plasma insulin levels (8.45 ± 1.17 ng/mL vs. 1.98 ± 0.32 ng/mL, P < .05) compared to lean control mice. No effect of genistein on body weight of ob/ob mice (49.75 ± 0.97 g vs. 49.05 ± 1.13 g) or plasma insulin levels (8.56 ± 0.59 vs. 8.45 ± 1.17 ng/mL) was observed after 4 weeks of treatment.
Data are expressed as mean ± SEM for 8–10 mice per group.
P < .05, compared to lean control mice.
Effects of genistein on dietary intake, body temperature, and T3 levels
Food intake in control and ob/ob mice is illustrated in Figure 1. As expected, food intake, expressed in grams/day, was higher (∼60%) in ob/ob mice compared to the lean control mice throughout the study, confirming the hedonic feeding behavior of obese mice. Interestingly, obese mice fed a genistein diet consumed a significantly higher amount of food during week 3 (14.1 cal/day vs. 16.34 cal/day) and week 4 (13.6 cal/day vs. 15.6 cal/day) compared to ob/ob mice fed a genistein-free diet, despite maintaining a body weight similar to ob/ob mice.

Effects of obesity and genistein treatment on food intake. Data are expressed as mean ± SEM for 8–10 mice per group. *P < .05, ob/ob effect compared to lean controls. # P < .05, ob/ob genistein effect. SEM, standard error of the mean.
The increase in food intake in genistein-fed ob/ob mice suggests that body temperature increased in response to the isoflavone diet. To detect these potential changes, a rectal thermometer was used biweekly to monitor temperature. As shown in Figure 2A, body temperature was consistently lower in ob/ob mice compared to lean control mice (32.13°C ± 0.29°C, n = 11 vs. 33.6°C ± 0.27°C, n = 10, P < .05). However, following genistein treatment, body temperature was significantly increased in ob/ob mice compared to nontreated obese mice (33.44°C ± 0.28°C, n = 8 vs. 31.43°C ± 0.28°C, n = 11, P < .05). Interestingly, genistein reversed the mild hypothermia associated as seen in this model of obesity, such that body temperature following treatment was the same as measured in lean control mice (Fig. 2B). Whether the effect of genistein on body temperature was hormonally induced, we measured levels of T3. Plasma levels of T3 were decreased in ob/ob mice compared to lean control mice (3.42 ± 0.44 pg/mL vs. 17.42 ± 2.80 pg/mL, P < .05). Genistein treatment produced an increase in circulating T3 levels, suggesting that the increases in body temperature and food intake are associated with improved thyroid hormone status (Fig. 2C). Circulating T3 in ob/ob mice with genistein treatment was similar to levels seen in lean controls (16.53 ± 5.07 pg/mL).
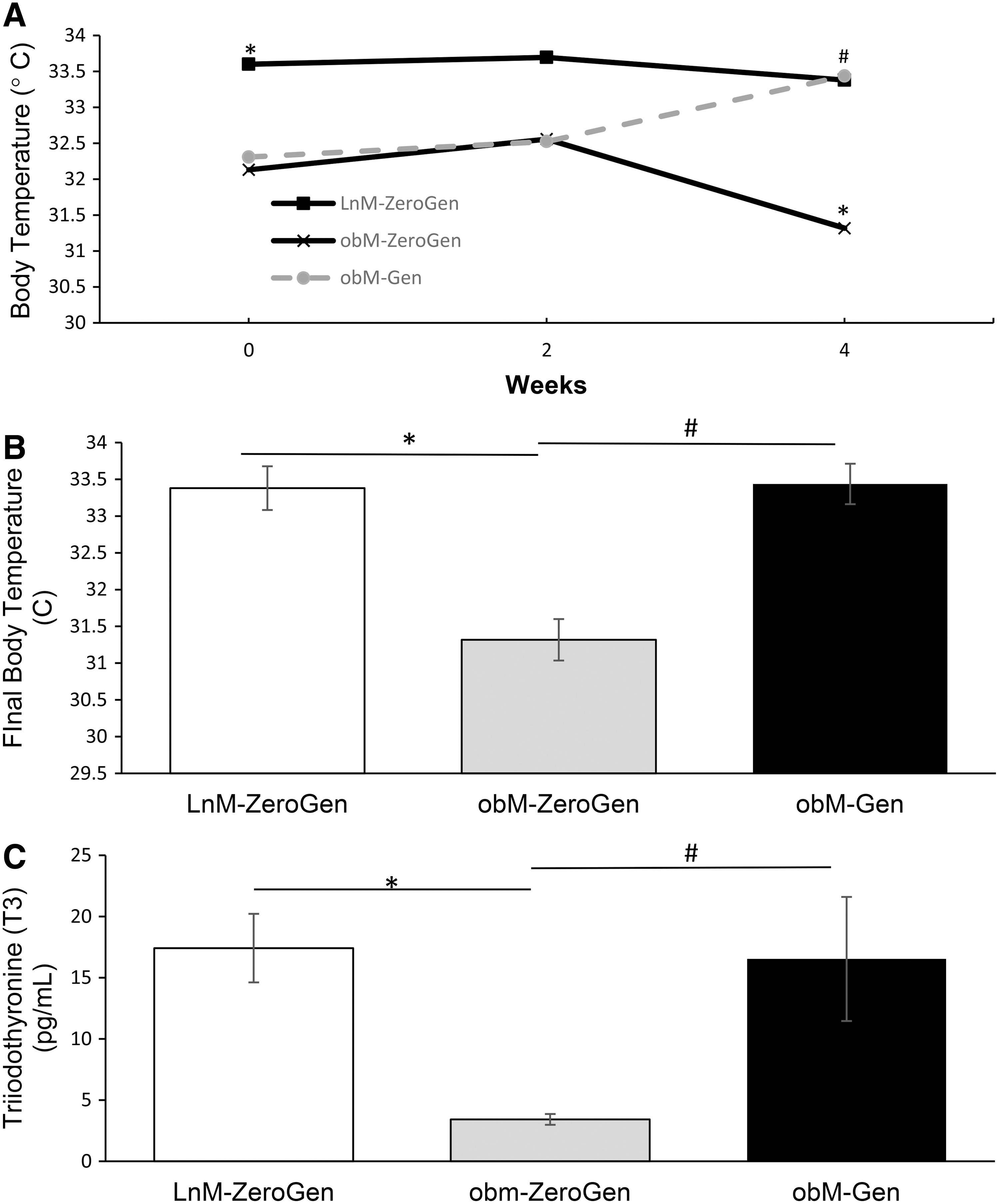
Effects of obesity and genistein treatment on body temperature and T3 levels.
Effects of genistein on corticosterone and protein expression of 11β-HSD1 and 2
Figure 3 illustrates the effects of obesity and genistein treatment on plasma corticosterone levels. Plasma levels of corticosterone were significantly increased in ob/ob compared to lean control mice (398.67 ± 26.09 ng/mL vs. 233.39 ± 32.35 ng/mL, P < .05). Treatment with genistein was beneficial and reduced corticosterone levels of ob/ob mice to comparable control levels (264.24 ± 34.19 ng/mL). We then measured whether the reduction in corticosterone was associated with changes in the central and contrasting HSD enzymes involved in the corticosterone-11-dehydrocorticosterone shunt. Hepatic 11β-HSD1 converts inactive corticosterone to active corticosterone, which can bind to the GR. Consistent with earlier reports, 23 we observed a significant reduction in hepatic 11β-HSD1 protein expression in ob/ob mice compared to lean mice. Genistein had no effect on the expression of this enzyme. Accompanying the decrease in 11β-HSD1 expression was a decrease in GR protein expression in liver of ob/ob mice (Fig. 4). Renal 11β-HSD2, which converts active corticosterone into the inert 11-dehydrocorticosterone form, on the other hand, was significantly increased in ob/ob mice compared to lean controls. Interestingly, expression of renal 11β-HSD2 was decreased in genistein-fed ob/ob mice.
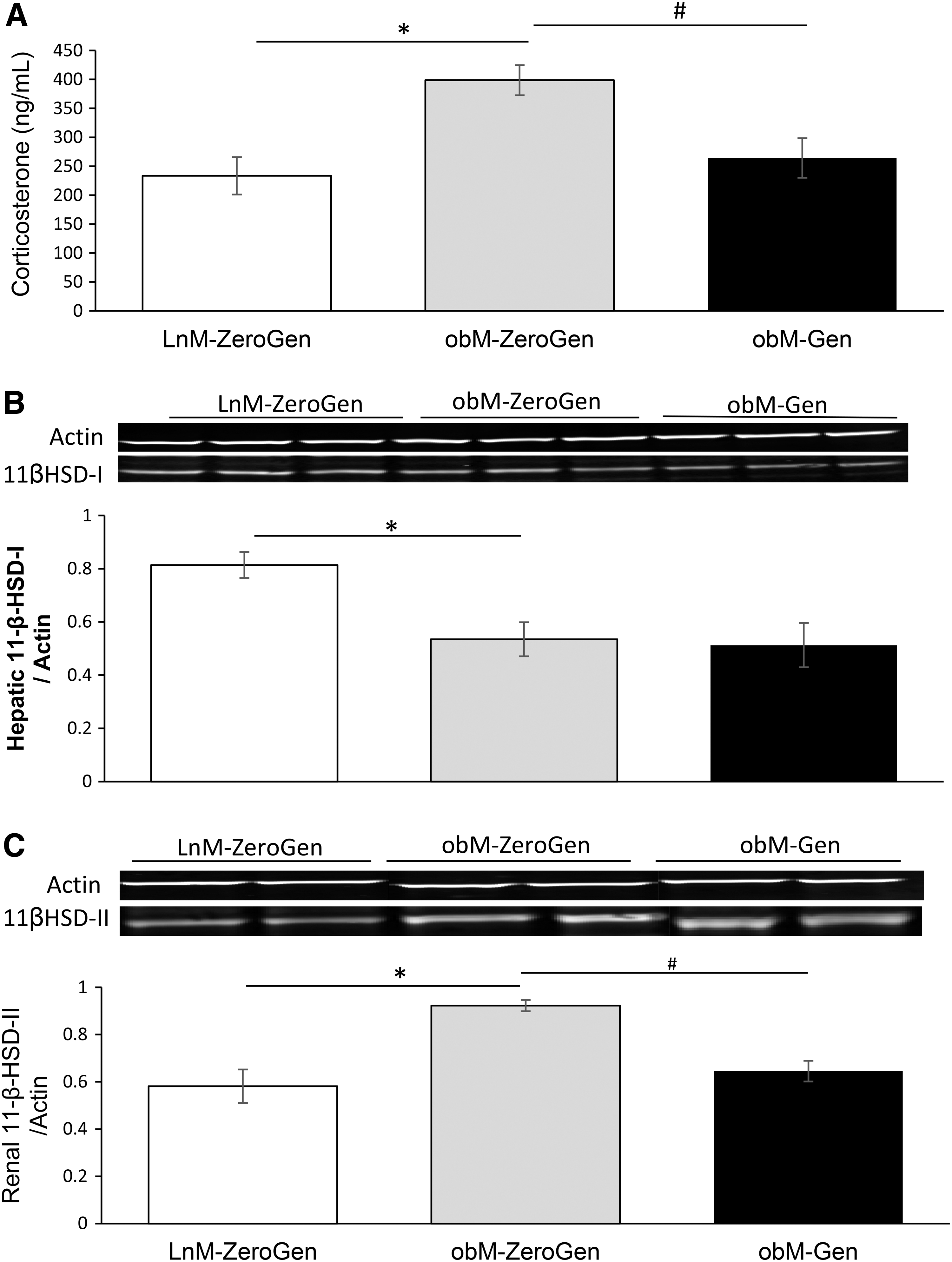
Effects of obesity and genistein treatment on

Effects of obesity and genistein treatment on protein expression of the hepatic GR. Data are expressed as mean ± SEM for 8–10 mice per group. *P < .05, ob/ob effect compared to lean controls. GR, glucocorticoid receptor.
Discussion
The transition from physical activity to sedentary lifestyle has been striking in recent decades, resulting in an alarming increase in the rates of metabolic syndrome, obesity, and diabetes, all of which predispose the patient with these conditions to cardiovascular disorders. The nonpharmacological approach for the management of obesity and diabetes focuses on a healthy dietary intervention. This can improve the obesity and associated risk factors, such as hypertension, hyperlipidemia, impaired glucose tolerance, and hypercortisolemia. 24 In this study, we demonstrate the potential beneficial effects of a genistein-rich diet on the phenotype of obese diabetic mice. We chose the ob/ob mouse because of its close similarity to the human condition of severe obesity and diabetes. A feature of this model is leptin deficiency, causing hyperphagia and obesity, hyperglycemia, insulin resistance, as well as a Cushingoid resemblance. 25 Although leptin deficiency is rarely the cause of these metabolic aberrations of obesity in humans, it is nonetheless detected in cases of severe obesity. 26 Given those characteristics, the goal of our study was to examine how a genistein diet might modulate the phenotype generally associated with this condition. Our results indicate that significant benefits occur from feeding ob/ob mice a diet rich in genistein. Although there was no effect of genistein on the obese state, expressed as fat content and hyperinsulinemia, changes in the hormonal profile and body temperature were observed. Indeed, we show that genistein caused a decrease in the plasma levels of corticosterone in ob/ob mice, suggesting decreased endogenous glucocorticoid production. Our main results indicate that treatment of ob/ob mice improved thyroid hormone T3 levels and this was associated with corresponding increases in body temperature and food intake.
The ob/ob mouse exhibits hypothermia at normal ambient temperatures and an accelerated loss in body temperature when exposed to cold stress. 27 This hypothermia is due to a low metabolic rate, impairment in thermoregulation, 28 and decreased sympathetic tone and thyroid function. 20,28 Recent evidence indicates that the increased susceptibility of the ob/ob mouse to hypothermia is also attributed to leptin deficiency, since leptin administration increases both core temperature and thermal conduction. 29 Our results confirm earlier observations that the ob/ob mouse displays a decreased body temperature compared to their lean counterparts. 20 Introducing genistein into the diet reversed this hypothermia and increased circulating T3 levels, suggesting that genistein offers protection against hypothermia and raises the possibility of a relationship between this isoflavone and increased energy expenditure in the obese mouse. In support of this possibility, recent work indicates that application of genistein to 3T3-L1 adipocytes induced a shift in gene expression consistent with adipocyte beigeing. 30 Induction of adipocyte beigeing by genistein was associated with increased mRNA expression of Cebpβ, which mediates the conversion of white to beige adipocytes 23 and stimulated the expression of uncoupling protein 1 (UCP1), as well as oxygen consumption and greater mitochondrial uncoupling. 30 Furthermore, by altering peripheral thyroid hormone metabolism since there is no impairment in the hypothalamus-pituitary-thyroid axis in the ob/ob mouse, 20 the increase in body temperature and energy expenditure observed in ob/ob mice with genistein may occur by similar mechanisms relating to UCP1 expression in response to increased T3 level. 31 While the role of genistein in thyroid hormone metabolism remains unclear, better understanding of the process of peripheral thyroid hormone conversion and deiodination by this isoflavone will identify mechanisms in inducing these changes.
Accumulating evidence has shown that glucocorticoid excess leads to disordered eating, defects in energy metabolism, and the development of insulin resistance and obesity. The ob/ob mouse is hypercorticosteronemic (hypercortisolemia in humans) as a result of hypertrophy of the adrenal cortex 32 and leptin deficiency causing impaired intrahepatic regulation of 11β-HSD1. 23 Recent evidence indicates that the elevated levels of corticosterone in plasma of obese Zucker rats and ob/ob mice are believed to be due to decreased expression of hepatic 11β-HSD1 that might stimulate the hypothalamic-pituitary-adrenal axis (HPA axis) to increase the production of endogenous corticosterone. 33 Our results are consistent with these earlier findings as we show that liver from ob/ob mice exhibited decreased expression of 11β-HSD1 with corresponding elevated concentrations of corticosterone in plasma. 23 Treating ob/ob mice with genistein normalized corticosterone levels, but failed to reserve the insulin resistance and the obese phenotype. Furthermore, genistein did not influence the expression of hepatic 11β-HSD1, suggesting that the decrease in circulating corticosterone levels may involve other mechanisms. One possibility explaining reduced corticosterone levels may include direct effects on the HPA axis. An inhibitory effect of genistein on basal and ACTH-stimulated secretion of corticosterone was observed in porcine adrenocortical cell 34 and in cultured human postnatal cortical cells and adult adrenocortical H295 cell lines. 35 Others have also reported this effect of genistein in vivo using middle-aged orchiectomized rats. 36 Our findings could suggest that genistein diminished the levels of corticosterone through these mechanisms.
While the role of 11β-HSD1 in liver and adipose tissue has been extensively studied in rodent and human obesity, few studies to date have investigated the effects of obesity and treatment on the expression of 11β-HSD2 in kidney. This microsomal enzyme is abundantly expressed in kidney and converts active corticosterone to the inactive 11-dehydrocorticosterone form, thus preventing corticosterone from binding to the mineralocorticoid receptor. Restricting local glucocorticoids from binding to this receptor prevents their inappropriate activation of mineralocorticoid pathways. 11β-HSD2 is expressed in sites critical in the regulation and blood pressure, and deficiency of this enzyme is associated with electrolyte disturbances and hypertension. 37 We found an increased expression of renal 11β-HSD2 in ob/ob mice compared with lean controls, and these data are consistent with earlier work in the Zucker rat model of leptin resistance 33 and in the high fat–induced model of obesity in rats. 38 Enzyme activity of 11β-HSD2, expressed as the ratio of urinary free cortisone to urinary free cortisol ratio, is also markedly elevated in the severely obese patient. 39 Although increased expression of renal 11β-HSD2 promotes intracellular 11-dehydrocorticosterone formation, representing a defense mechanism against excess fat storage, increased expression of this enzyme better predicts a heightened supply of substrate for adipose tissue 11β-HSD1 activation. 38,40,41 Enhanced activity of 11β-HSD1 in adipose tissue promotes visceral adipose tissue accumulation in obese subjects, 42 correlates with insulin resistance, 39,43 and is previously reported in the ob/ob mouse. 44 Interestingly, the increase in 11β-HSD2 protein expression observed in kidney of ob/ob mice was normalized following genistein treatment, and extrapolating the earlier lessons on the contrasting roles of 11β-HSD to our current work, this could have a metabolic effect of reduced substrate for extrarenal 11β-HSD1.
The prevalence of obesity has increased dramatically over the recent decades and this is contributing to the rise in diabetes and associated cardiometabolic and vascular disorders. Pharmacotherapy options and various weight-loss surgical procedures for the treatment of obesity, although widely available, are expensive and associated with side effects and complications. The results of our study show metabolic benefits of genistein in a commonly used mouse model of obesity, and when considering the results of our previous studies on bone, heart, and gastrointestinal function, 11 genistein appears to have potential as a practical and viable nutritional intervention. The mice in this study were fed genistein for only a period of 4 weeks and based on this short duration, it cannot be excluded that the obesity expressed as fat mass and body weight may have improved with a longer period of treatment. Nonetheless, our results demonstrate that genistein ameliorated circulating thyroid hormone T3 levels, increased body temperature and food intake, and reversed the hypercorticosteronemia. More studies are needed to elucidate the intracellular mechanisms associated with these benefits of genistein in obesity.
Footnotes
Acknowledgments
This work was supported by the Midwestern University Office of Research and Sponsored Programs (L.A., T.L.B.), Diabetes Action and Research Education Foundation (L.A.), and Department of Biomedical Sciences (S.R.).
Author Disclosure Statement
No competing financial interests exist.