
Abstract
Menopause induces a loss of bone as a result of estrogen deficiency. Despite pharmaceutical options for the treatment of osteopenia and osteoporosis, many aging women use dietary supplements with estrogenic activity to prevent bone loss and other menopausal-related symptoms. Such supplements are yet to be tested for efficacy against a Food and Drug Administration (FDA) approved medication for menopausal bone loss such as zoledronic acid (ZA). The postmenopausal rat model was used to investigate the efficacy of various synergistic phytochemical blends mixed into the diet for 16 weeks. Retired-breeder, Fischer 344 rats were randomly assigned to sham or ovariectomy surgery and 4 treatment groups: ZA; genistein supplementation; and a low dose and high dose blend of genistein, resveratrol, and quercetin. Ovariectomy resulted in a loss of both trabecular and cortical bone which was prevented with ZA. The phytochemical blends tested were unable to reverse these losses. Despite the lack of effectiveness in preventing bone loss, a significant dose–response trend was observed in the phytochemical-rich diets in bone adipocyte number compared to ovariectomized control rats. Data from this study indicate that estrogenic phytochemicals are not as efficacious as ZA in preventing menopausal-related bone loss but may have beneficial effects on bone marrow adiposity in rats.
Introduction
T
Partly as a result of safety concerns with hormone replacement therapy and side effects of pharmaceuticals, the number of postmenopausal women taking botanical and dietary supplements has increased in recent years 3 ; 60–80% of peri- and postmenopausal women take dietary supplements. 3,4 Numerous studies report the potential of resveratrol and genistein to combat against bone loss in animal models supporting a mechanism of protection mediated by anti-inflammatory signaling. 5 –7 Furthermore, there is growing evidence that quercetin also may improve bone turnover markers 8 and bone mineral density 9 in animal models. While clinical trials of resveratrol and quercetin are lacking, clinical trials with the soy isoflavones have largely been inconclusive, demonstrating both efficacy and inefficacy to protect against bone loss in menopausal and postmenopausal women. For example, genistein supplementation improved both femoral and lumbar bone mineral density in postmenopausal women. 10 Soy isoflavones did not prevent a loss in bone density in both a Taiwanese postmenopausal population 11 and in the United States. 12 One such hypothesis for the inability of isoflavones to demonstrate explicit protection in clinical trials is that the human equivalent doses necessary to replicate the rodent models are high, increasing the number of daily pills one must ingest and reducing study compliance. 13
Phytochemicals such as genistein, resveratrol, and quercetin have demonstrated synergistic properties. 14 In 3T3–L1 adipocytes a combination of these phytochemicals decreased adipogenesis and increased apoptosis compared to singular treatment with any of the individual compounds and their calculated additive response. 14 Furthermore, this combination, along with vitamin D, improved bone density and decreased bone marrow adipocyte accumulation in ovariectomized rats. 15 While the role of bone marrow adipocytes on bone loss and fracture risk is contested, 16 both aging and osteoporosis are positively associated with bone marrow fat. Furthermore, adipocytes and osteoblasts share a common mesenchymal stem cell lineage. Various signals, including runt-related transcription factor 2 (RUNX2) and bone morphogenetic proteins (BMP-2/4), can stimulate osteoblast differentiation, whereas CCAAT-enhancer-binding protein-β (C/EBPβ) and peroxisome proliferator-activated receptor-γ (PPARγ) can induce an adipocyte commitment. 17 Unsurprisingly, treatment with phytochemicals has demonstrated osteoblast differentiation in various in vitro models 18 –20 and also show potential for synergism in bone. 21 Such phytochemicals appear to act on distinct intracellular signaling pathways and further directly interact with the metabolism of each other. For example, quercetin prevents phase II metabolism of resveratrol thereby increasing its bioavailability. 22 Hence, synergistic phytochemicals may be useful to combat menopausal bone loss. However, their efficacy against that of a pharmaceutical drug is yet to be explored.
In the current study, we tested the hypothesis that a prolonged duration of dietary intake of a combination of vitamin D, resveratrol, quercetin, and genistein, which has demonstrated efficacy in vivo and in vitro to reduce bone loss and adiposity, would be comparable to currently available therapies in the menopausal-induced osteoporotic population. We investigated the efficacy of a low- and high-dose phytochemical blend compared to a nonsynergistic mix of vitamin D and genistein, a patented combination marketed as a prescription medical supplement, and zoledronic acid (ZA), a Food and Drug Administration (FDA)-approved bisphosphonate that inhibits osteoclast function and maturation.
Materials and Methods
Animal care
Aged, retired breeder Fischer 344 rats (∼12 months of age; n = 12/group) were purchased from Taconic and Charles River Laboratories National Institute of Aging colony (Germantown, NY, USA and Wilmington, MA, USA, respectively). Rats were randomized by body weight, and bilateral ovariectomy or sham surgery was performed under 2.5% isoflurane anesthesia while applying sterile technique. One week following surgery, rats assigned to the ZA group received an intravenous injection (20 μg/kg body weight) of ZA (Novartis Pharmaceuticals, East Hanover, NJ, USA), and dietary treatments began. Both food and water were available ad libitum. Following 16 weeks of dietary phytochemical supplementation, rats were fasted for 2 h and euthanized by cervical decapitation under 2.5% isoflurane anesthesia. All experimental procedures described herein were approved by the Institutional Animal Care and Use Committee at the University of Georgia before initiating the study.
Diets
The diets used in this study were selected based on a prescribed antiosteoporotic medicinal food product containing genistein and from experiments performed in our laboratory that have demonstrated synergistic capabilities to promote antiadipogenic properties in vitro. 14 The powdered basal diet (AIN-93M; TestDiet, St. Louis, MO, USA) was provided to the rats randomized into sham, ovariectomy, and ovariectomy+ZA groups. A modified AIN diet containing supplemented vitamin D at 2.4 (IU/g diet) was used in the three phytochemical diets (TestDiet) as reported in Table 1. Vitamin D was supplemented in the dietary treatment groups as serum 25(OH)D deficiency in patients at risk or in treatment for osteoporosis is common (52% in North America). 23 Hence, vitamin D supplementation would be recommended in this population, in addition to other forms of therapy (i.e., pharmacologic). All compounds were purchased from Spectrum Chemicals (Gardena, CA, USA). Food intake was monitored daily.
Phytochemicals (Spectrum Chemicals, Gardena, CA, USA) were mixed into the AIN-93M diet from TestDiet, St. Louis, MO, USA, with or without added vitamin D3.
Daily intake of genistein was equivalent to the concentrations of the genistein compound found in a prescribed medicinal food product for treatment of osteoporosis (∼25 mg/day).
OVX, ovariectomized.
Dual X-ray absorptiometry
Bone mineral density was measured using the Lunar PIXImus. X-ray densitometer for small animals (GE Lunar Corp., Madison, WI, USA) was measured at weeks 4 and 16 postovariectomy. Rats were anesthetized under isoflurane (2–2.5%) throughout the scan. Because the field of the image was not large enough to scan the entire size of a rat, bone mineral density was determined utilizing a method previously described. 24 Samples with poor image quality were removed from analysis.
Micro-computed tomography
Excised rat femurs (right) were fixed in neutral buffered formalin for 24 h and transferred to 70% ethanol before shipment. Imaging and analysis were performed at the University of Alabama at Birmingham Small Animal Phenotyping Laboratory by a technician blinded to the treatment groups. Samples were scanned using the Scanco mCT40 desktop cone-beam micro-computed tomography (μCT) scanner (Scanco Medical AG, Brüttisellen, Switzerland). The femur was placed inverted in a 16-mm diameter scanning holder and scanned at the following settings: 16 mm resolution, 70 kVp, and 114 μA with an integration time of 200 msec. Scans were automatically reconstructed into two-dimensional slices, and all slices were analyzed using the μCT Evaluation Program (v5.0A; Scanco Medical AG). For the cortical bone, the midshaft of the bone was determined, and the scan started 0.4 mm above the midpoint and ended 0.4 mm below the midpoint for a scan of 50 slices. The region of interest was drawn on every image slice and fitted to the outside of the cortical bone, to include all the bone and marrow. The threshold for bone was set at 400. The three-dimensional (3D) reconstruction was performed using all the outlined slices. For the trabecular bone, the scan was started distal to the growth plate and consisted of 312 image slices. The region of interest started at the point on the scan where the epiphysis ended. Two hundred slices were outlined from this point, on the inside of the cortical bone, enclosing the trabecular bone and marrow. A threshold for trabecular bone was at 259, and the 3D analysis was performed on the 200 slices.
Hematoxylin and eosin Y staining
A representative sample of an n = 5–6/treatment group was used for further histology study. The right proximal tibia was removed at necropsy and was preserved in 10% formalin for 24 h. Samples were then washed briefly in water and stored at 4°C in 70% ethanol before decalcification in 10% EDTA. Following decalcification, samples were dehydrated, cleared in xylene, and embedded in paraffin before sectioning at 4–5 μm thickness. Hematoxylin and eosin Y (H&E) staining was performed using the CV Select H&E Kit (BBC Biochemical, Mt. Vernon, WA, USA) following manufacturer's protocols. Images were taken at 5 × on a Zeiss Axioplan microscope mounted with a Zeiss HRc camera (Carl Zeiss AG, Oberkochen, Germany). Bone adipocytes for each sample were counted over a 0.1 mm2 region (bone area) and averaged from three independent regions on each slide using Image-Pro Plus (Media Cybernetics, Inc., Rockville, MD, USA).
Tartrate-resistant acid phosphatase assay
Slides were prepared of the tibia as described in the previous section. Following hydration, slides were incubated for 1 h (37°C) in the dark in a solution of 44 mL water, 2 mL acetate solution, 2 mL naphthol AS-BI phosphoric solution, and 2 mL tartrate solution (Sigma-Aldrich, St. Louis, MO, USA). Slides were washed and then stained in acid hematoxylin (Sigma-Aldrich). Section images were captured at 200 × , and osteoclasts on each bone surface (BS) were counted in replicates of five over a 0.1 mm2 region. Surface lengths were measured using Image-Pro Plus (Media Cybernetics, Inc.) and were expressed as osteoclast number per bone parameter (mm; N.Oc/B.Pm).
Statistics
Sample size was determined to be an n = 6 at 0.90 power to obtain statistically significant differences in trabecular microarchitecture as a result of ovariectomy. Between-treatment group differences were measured by Tukey HSD (honest significant difference) post hoc testing or Dunn's test in the case of nonparametric data using GraphPad Prism (version 6.04). To determine if a phytochemical dose-dependent effect existed in trabecular adipocyte trend analysis was performed in GraphPad Prism. The trend analysis quantifies the linear relationship between increasing phytochemical treatment and adipocyte count. Significance was set at P < .05.
Results
Effect of ovariectomy on food intake, body, and uterine weight
As reported previously, 25 food intake was significantly affected by ovariectomy (P < .05 for analysis of variance). Despite lack of significant between-group differences, all ovariectomized groups, other than rats fed the low-dose phytochemical diet, had near significant reductions in daily food consumption (13.6 ± 0.25 g, sham, vs. 12.9 ± 0.15 g, ovariectomized). Body weight at 16 weeks postovariectomy was not different between groups (311.7 ± 6.2 g, sham, vs. 315.4 ± 4.0 g, ovariectomized), and no effect of phytochemical treatments was measured. As expected, ovariectomy resulted in a 67% reduction in uterus weight compared to sham rats (0.48 ± 0.02 g vs. 0.16 ± 0.01 g, P < .001). Dietary supplementation with phytochemicals did not increase uterine weight relative to ovariectomized rats.
Dual X-ray absorptiometry measurements
At the 4-week baseline measurement, no differences existed in total femur, diaphysis, or metaphysis bone mineral density between most groups (Table 2). However, a significant difference between ZA-treated and the low-dose phytochemical group was observed (P < .01), which may be explained by the ZA injection that occurred 2 weeks before the first Dual X-ray absorptiometry (DXA) measurement. The percent change in bone mineral density was measured at 4 and 16 weeks postovariectomy. In the total femoral density, ovariectomy resulted in significant bone loss compared to sham rats (P < .05; Table 2). This loss was entirely reversed with ZA treatment and treatment with the low-dose phytochemical blend (P < .05). There was no effect of the genistein or high dose treatment in femoral density. Both the diaphysis and metaphysis of the femur were also measured; however, due to the large variation within each treatment no differences were observed.
BMD of the femur was performed 4 and 16 weeks following ovariectomy by dual X-ray absorptiometry in rats treated with ZA, genistein, or a combination of genistein, resveratrol, and quercetin. Percent change in BMD was calculated from the difference between 16 and 4 weeks. Groups: Sham (n = 12), OVX (n = 9), OVX+ZA (n = 11), OVX+genistein (n = 10), OVX+low dose (n = 12), OVX+high dose (n = 10). Data are presented as mean ± SE. Groups with differing letters are statistically different as determined by Tukey's HSD posttest at P ≤ .05. P value for each one-way ANOVA is reported in the final column for each measurement.
ANOVA, analysis of variance; BMD, bone mineral density; HSD, honest significant difference; SE, standard error; ZA, zoledronic acid.
Overall, there were little between-group differences in baseline lumbar spine measurements at 4 weeks (Table 3). Similar to what was observed in the femur, a significant difference between the ZA and low dose groups in L4 was found (P < .05). Aging resulted in a loss of lumbar spine mineral density in all groups (Table 3). Total lumbar spine bone mineral density remained unaffected due to ovariectomy; however, rats fed the high-dose blend diet had greater reductions in lumbar density compared to ZA-treated rats (P < .05). While there were no between-group differences in L4 density, ovariectomy induced greater reductions in L5 density compared to sham rats which were preventable with ZA treatment (P < .05). Rats fed a high-dose blend diet also experienced greater reductions in L5 bone mineral density compared to sham and ZA-treated rats (P < .05).
BMD of the lumbar spine was performed 4 and 16 weeks following ovariectomy by dual X-ray absorptiometry in rats treated with ZA, genistein, or a combination of genistein, resveratrol, and quercetin. Percent change in BMD was calculated from the difference between 16 and 4 weeks. Groups: Sham (n = 9), OVX (n = 8), OVX+ZA (n = 12), OVX+genistein (n = 9), OVX+low dose (n = 12), OVX+high dose (n = 9). Data are presented as mean ± SE. Groups with differing letters are statistically different as determined by Tukey's HSD posttest at P ≤ .05. P value for each one-way ANOVA is reported in the final column for each measurement.
L, lumbar vertebrae.
Cortical and trabecular bone measurements
Representative images of femoral cortical bone are presented in Figure 1A, and reported means are described in Table 4. Cortical bone volume (BV) was not impacted by ovariectomy. However, ZA increased BV compared to sham controls (P < .01). Phytochemical treatments did not change BV compared to ovariectomized controls. Bone volume density (BV/TV [total volume]) was decreased due to ovariectomy (P < .01). This effect was entirely prevented with ZA treatment (P < .001), whereas phytochemical treatments failed to protect from this loss. A similar result was found in measures of cortical thickness; ZA treatment protected from ovariectomy-induced loss (P < .05) which was not observed in rats fed the phytochemical-rich diets.

Femoral bone architecture following ovariectomy.
Cortical bone was measured by μCT in sham or OVX rats following 16 weeks of treatment with ZA, genistein, or a combination of genistein, resveratrol, and quercetin. Quantitative measurements were performed at the University of Alabama at Birmingham. Groups: Sham (n = 12), OVX (n = 8), OVX+ZA (n = 11), OVX+genistein (n = 11), OVX+low dose (n = 12), OVX+high dose (n = 11). Data are presented as mean ± SE. Groups with differing letters are statistically different as determined by Tukey's HSD posttest at P ≤ .05. P value for each one-way ANOVA is reported in the final column for each measurement.
μCT, micro-computed tomography; BV, bone volume; TV, total volume.
Representative images of femoral trabecular bone are presented in Figure 1A, and reported means are described in Table 5. Ovariectomy reduced trabecular BV (P < .0001) and bone volume density (BV/TV; P < .0001) compared to sham controls, which were prevented by ZA treatment (P < .05). Phytochemical-rich diets failed to protect from this loss; however, both the low- and high-dose phytochemical treatments were not significantly different from ZA-treated rats. This may indicate that the phytochemicals were having a positive effect on some bone parameters, but were not able to completely reverse the loss in BV and bone volume density. BS (P < .0001), surface-to-volume ratio (BS/BV; P < .001), trabecular number (Tb.N.; Fig. 1B; P < .0001), trabecular thickness (Tb.Th.; P < .01), trabecular spacing (Tb.Sp.; Fig. 1C; P < .0001), and degree of anisotropy (DA; Fig. 1D; P < .0001) were all reduced compared to sham controls as a result of ovariectomy. ZA treatment prevented reductions in all aforementioned measurements other than Tb.Th., which was also reduced in ZA-treated rats (P < .05 vs. sham controls). Similarly, all aforementioned measurements were also reduced in phytochemical treatment groups (P < .01 vs. sham controls), indicating that dietary phytochemical supplementation failed to reverse losses in trabecular bone.
Cortical bone was measured by μCT in sham or OVX rats following 16 weeks of treatment with ZA, genistein, or a combination of genistein, resveratrol, and quercetin. Quantitative measurements were performed at the University of Alabama. Groups: Sham (n = 12), OVX (n = 8), OVX+ZA (n = 9), OVX+genistein (n = 10), OVX+low dose (n = 12), OVX+high dose (n = 9). Data are presented as mean ± SE. Groups with differing letters are statistically different as determined by Tukey's HSD posttest at P ≤ .05. P value for each one-way ANOVA is reported in the final column for each measurement.
BS, bone surface; BS/BV, bone surface-to-volume ratio; DA, degree of anisotropy; Tb.N, trabecular number; Tb.Sp., trabecular spacing; Tb.Th, trabecular thickness.
Trabecular adipocytes and osteoclasts
Adipocytes in the right tibia were counted in triplicate for each rat (Fig. 2A). There was no effect of ovariectomy or ZA treatment on the number of adipocytes in the bone marrow (P = .07; Fig. 2B). However, a significant trend to decrease bone adipocytes was found between the phytochemical treated rats compared to ovariectomized (P = .008) rats.
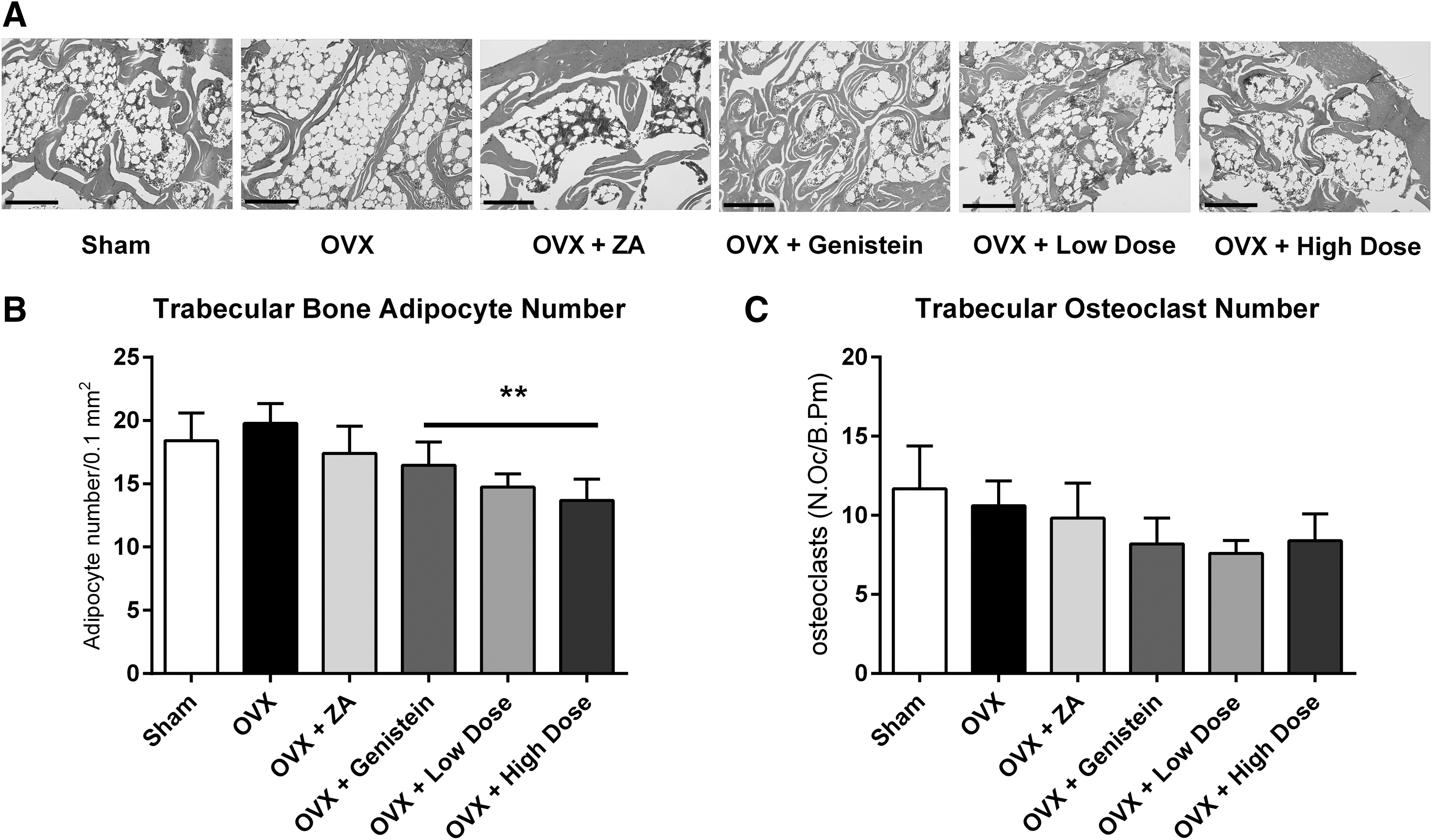
Trabecular bone adipocytes and osteoclasts following ovariectomy.
Osteoclasts were stained using the tartrate-resistant acid phosphatase (TRAP)-based assay, counted and normalized to bone length (mm). No differences in osteoclast number were measured between groups (P = .51; Fig. 2C).
Discussion
In this study, the FDA approved treatment for osteoporosis; ZA prevented the bone loss induced by ovariectomy in trabecular and cortical bone in aged, retired breeder rats. ZA's success as an anti-osteoporotic drug is well established, and it is currently approved for treatment of menopausal bone loss. However, high-dose bisphosphonate use is associated with osteonecrosis of the jaw and bone fractures. 26,27 These adverse effects may lead women and practitioners to seek alternative therapies such as phytochemical supplementation. In this study, the selected phytochemical blends, some of which were shown in a previous 8-week study to reduce bone loss, 15 were unable to prevent menopausal-related bone loss measured by DXA and μCT compared to ZA in a 16-week study in aged rats. Similarly, ovariectomized rats treated with the genistein and vitamin D diet at similar concentrations of a medicinal food product that is prescribed as an antiosteoporotic failed to exert beneficial protection against bone loss in the current study.
The lack of efficacy of phytochemicals to protect against bone loss is not novel. Both animal 28 –30 and randomized clinical control trials 11,12 have reported limited effects of estrogenic dietary supplements in bone. Despite these negative findings, other studies in both rodents 31 and humans 32 report bone promoting effects. It is important to note that we utilized an aged ovariectomy rat model, one similar to menopause and treated with the dietary phytochemicals for a prolonged period of time (16 weeks). Many studies that investigate the efficacy of phytochemicals do so in younger virgin rats and/or in much shorter treatment periods (under 10 weeks) similar to our previous study. 15,33 These differences may account for the inability of phytochemicals to protect against bone loss in this study.
Bone loss following ovariectomy has a triphasic response with a rapid loss for the first 100 days, a period of stabilization for another ∼170 days, followed by a final slow rate of bone loss at 270 days postovariectomy. 34 In the current study, it is feasible to believe that the rats had progressed into a stabilization phase, whereas most studies investigating ovariectomy-induced osteoporosis are comfortably within the rapid phase of bone loss. Thus, it is reasonable to hypothesize that the longer the phytochemical intervention is in an ovariectomized rat, hence the longer hormonal withdrawal, the more bone loss that must be overcome. As maximal bone loss due to ovariectomy had been achieved in this study, the relatively low dose of phytochemicals may not have induced a robust enough effect to prevent or reverse the dramatic loss of trabecular bone induced by the deficiency of ovarian hormones. This study highlights the importance of timing and length of intervention as important factors in the bone sparing effects of phytochemicals.
While the dietary phytochemical blends used in this study failed to exert protective effects on bone density, a dose-dependent trend to reduce bone adiposity was observed compared to ovariectomized controls. This finding is similar to what was observed in a shorter 8-week intervention with phytochemicals. 15 Unlike what was reported in the aforementioned study, osteoclast numbers were not changed due to ovariectomy or treatment in this study. We are not the first to demonstrate a discoordination of bone adiposity with femoral bone loss in an ovariectomized model, 35,36 and thus, these results suggest that the relationship between the two may be more dynamic than previously suspected. Furthermore, the role of aging and prolonged estrogen loss may disrupt trabecular structure through unique mechanisms beyond that of bone marrow adiposity, 37 which may not be rescued by phytochemicals alone.
While the reduction of bone marrow adiposity did not accompany protection against bone loss in the current study, bone marrow adipocytes are related to whole body metabolism. It has been suggested that bone adipocytes may serve as a source of fatty acids in the marrow microenvironment of highly metabolic cells, including osteoblasts and osteoclasts, and for hematopoietic differentiation. 38 Hence, the reduced bone adiposity observed in our phytochemical treatment groups may reflect less metabolic demand of adjacent cells. However, as we did not observe reduced bone marrow adiposity in the ovariectomized group, reduced metabolism does not appear to be a plausible explanation. As mentioned previously, the phytochemicals used in this study have lipolytic properties in vitro. 14 Hence, we additionally investigated changes in systemic lipid shuttling and fatty liver in the rats from the current study. 25 In this work we reported reduced lipid accumulation in the retroperitoneal adipose depot with all three phytochemical treatments, increased serum-free fatty acids, and increased hepatic lipid storage compared to both sham and ovariectomized controls. Thus, the trend to reduce bone marrow adiposity with phytochemicals may reflect a systemic lipolytic effect with treatment. However, it is not clear if the reduction in bone marrow adipocytes in the phytochemical treatment groups had any effect on bone quality in our model.
In summary, our data demonstrate that the phytochemical blends used in this study, selected for their relatively low dose and synergistic properties, were insufficient to protect bone loss triggered by long-term estrogen withdrawal in the rat. Furthermore, our data show that neither the combination of phytochemicals nor the previously defined medicinal food product (i.e., genistein and vitamin D) were as efficacious in preventing bone loss as was ZA in this model. It is hypothesized that the observed lack of efficacy may be related to the prolonged study duration (16 weeks) and the dynamic nature of bone loss following estrogen withdrawal, which requires further study. However, because of the findings of this study, along with the relatively inconclusive clinical control trials investigating bone-protective effects of phytochemicals, patients at risk of menopausal-related osteopenia or osteoporosis should explore the use of pharmaceutical options such as estrogen replacement and ZA for preventing bone loss.
Footnotes
Acknowledgment
This research was funded by a USDA SBIR grant awarded to AptoTec, Inc. (USDA-NIFA-SBIR-003497).
Author Disclosure Statement
Drs. Clifton A. Baile and Mary Anne Della-Fera served as the Chief Executive Officer and Chief Scientific Officer, respectively, and had no role in performing the experiment, assays, or data analysis. Dr. Suresh Ambati was employed by AptoTec, Inc. during the experimental phase of this study. None of the additional authors report a conflict of interest with the company.