
Abstract
The prevalence of obesity is expanding rapidly worldwide, making the disease a global burden with limited treatment options. The current obesity drug development trends suggest the possibility of reducing weight and reverse metabolic disturbances of obesity by controlling appetite. In this study, we screened more than 8000 plants from our plant library for the cannabinoid (CB1) receptor antagonists and identified Morus alba as a lead medicinal plant. Kuwanon G and Albanin G were isolated and identified from root-barks of Morus alba with 92% and 96% CB1 receptor ligand binding inhibitory activity, respectively. The bioflavonoid standardized extract was tested in the acute food intake study in rats at oral doses of 250 and 500 mg/kg for its appetite suppression activity. Diet-induced obesity in the C57BL/6J mice was used to evaluate the long-term food intake reduction activity and effect on the weight loss administered orally at 250 and 500 mg/kg for 7 weeks. Statistically significant and dose-dependent reduction in food intake was observed in both acute and long-term studies for the extract. Food intake reductions of 58.6% and 44.8% at 250 mg/kg and 50.1% and 44.3% at 500 mg/kg were observed at 1 and 2 h postfood provision, respectively. A 20% reduction in daily calorie intake was observed in the long-term study. Obese mice treated with the high dose of Morus root-bark extract showed 10.4 g (22.5%) and 7.1 g (16.5%) loss in body weight compared with the vehicle-treated obese animals (at week 7) and baseline, respectively. Statistically significant reductions in biochemical markers and visceral fat deposit were also observed. These results demonstrated that Morus alba extracts enriched in Kuwanon G, and Albanin G could be used alone to control appetite, manage body weight, and improve metabolic syndromes.
Introduction
Obesity is a consequence of an imbalance between abundant food consumption and limited energy expenditure. The prevalence of obesity is higher in industrialized countries due to easier access, bigger portions, and cheaper price of high calorie foods and beverages. According to the recent OECD (Organization for Economic Co-operation and Development) survey that included a population aged 15 years and older, the United States was found to be the most obese nation in the world, with a 38.2% obesity prevalence rate. 1 Unless there is a change to the present sedentary lifestyle and easy access of high calorie diets, this obesity rate is expected to reach 47% by the year 2030.
Obesity and obesity-related health issues are rapidly expanding globally at an alarming rate. The reality is that there are very limited treatment options. Among the existing prescribed or over-the-counter weight loss drugs on the market, none are considered completely safe and as a cure for obesity. Predominantly, the current clinically effective drugs aim at a reduction in food intake as the primary target for a successful management of obesity. For example, Lorcaserin (Belviq, selective 5-HT2C serotonin receptor agonist, FDA approved in 2012), Phentermin/Topiramate (Qsymia, Sympathomimetic/GABA and glutamate modulator, FDA approved in 2012), and Naltrexone/bupropion (Contrave, Dopamine-norepinephrin reuptake inhibitor/opioid receptor antagonist, FDA approved in 2014) are all marketed as appetite suppressants with limited acceptance. The maximum placebo subtracted weight loss achieved from these drugs was between 3.3% and 8.8% after 1 year of treatment. 2 A reduction as low as 5% in body weight has been shown to improve the comorbidities of obesity. 3 However, for morbidly obese patients, these reductions may not seem significant, especially taking the associated side effects of the prescription drugs into account. To date, Lorcaserin and Phentermin/Topiramate are still not approved by the European Medicines Agency in Europe because of their unwanted cardiovascular side effects. As a result, there are strong needs for other safer and effective alternatives. Some dietary supplements with defined mechanisms of action coupled with a proven record of safety and efficacy supported by scientific evidence may bridge the gap to fulfill the unmet demands.
Various targets have been evaluated for weight loss intervention. Among them, the cannabinoid receptor-1 (CB1) inverse agonists and neutral antagonists have shown some success with limitations. 4 Rimonabant and its class of drugs are good examples. 5 For many years, Cannabis sativa (marijuana) has been well known to stimulate food consumption through the action of its active component, Δ-9-tetrahydrocannabinol (THC), an exogenous cannabinoid. This phenomenon led to an investigation of the role of the endocannabinoid system in the regulation of obesity and associated metabolic syndrome targeting its two specific receptors known as CB1 and CB2. These receptors belong to the G-protein coupled receptor family characterized by seven trans-membrane loop domains. Both receptors belong to Gi/0 subclass and signal by negatively regulating cyclic AMP levels. CB1, the cannabinoid receptor involved in feeding behavior, is widely expressed in both brain and peripheral tissues, including adipose tissue, skeletal muscles, liver, and gastrointestinal tract. CB2 receptor is present in immune cells and is not involved in regulation of food consumption. With this understanding in this study, we screened more than 8079 plant extracts for CB1 receptor antagonists and identified Morus alba as the lead plant.
M. alba L. (Family: Moraceae), the mulberry or white berry plant, is native to northern China, and it has been cultivated and naturalized elsewhere, from India through the middle east to Southern Europe, and recently to North America. The root-bark of M. alba that is used in traditional medicine is known as Sang Bai Pi or Cortex Mori (Pharmacopoeia of the People's Republic of China, 2005). This herb is also known as Pong-na-moo in Korean and Sohakuhi in Japan. In contemporary pharmacological research, M. alba root-bark has been reported to have antibacterial, 6 antioxidant and hypoglycemic, 7,8 hypolipidemic, neuroprotective, antiulcer, analgesic, 9 –11 and anti-inflammatory activities. 12 Some of the prenylated flavonoids and stilbenoids such as morusin and mulberroside A are unique to Morus plants. 13
Previously, M. alba was used in combination with Yerba mate and Magnolia officinalis to give a standardized composition known as UP601. UP601 was tested in acute food intake, high fat diet (HFD)-induced and high-fat- and high-fructose-induced obesity disease models. In these models, administered at oral doses as low as 300 mg/kg of UP601 for 7 weeks in the long-term studies and as low as 230 mg/kg of UP601 in the acute food intake model, UP601 resulted in a significant decrease in acute and long-term food consumption, marked reduction in body weight, reduced fat mass without affecting lean body mass, improved nonalcoholic steatohepatitis scores, lowered blood glucose, total cholesterol, low density lipoprotein (LDL) and triglyceride, and reduced insulin and leptin while increasing ghrelin. 14,15 These significant improvements in body weight and metabolic markers are believed to be partially due to the biological effects of the bioflavonoid standardized extract from root-barks of M. alba.
In this study, M. alba root-bark-derived bioflavonoid extract was evaluated in acute food intake and HFD-induced animal models. The CB1 receptor antagonist data for the lead plant and its active compounds are also included.
Materials and Methods
Material preparation
Dried root-bark of M. alba was collected from Chongqing, China and identified by professor Shou-Yuen Zhao from Si-Chuan Chinese Traditional Medicine Research Institutes. A voucher of specimen of M. alba (P00329) was deposited at the plant library of Unigen (Seattle, WA, USA). When re-collected, M. alba root-barks were always characterized and confirmed in comparison with the original voucher specimens. M. alba root-bark extract was produced by 70% ethanol extraction of the ground root-bark powder with no less than 10% total bioflavonoids, including kuwanon G, albanin G, and morusin.
CB1 receptor binding activity
Cannabinoid receptor binding assays were used as a primary screening method to identify CB1 antagonist compounds. The assays were performed by using methods adapted from Reggio et al.
16
(CB1 receptor assay) and Munro et al.
17
(CB2 receptor assay). Briefly, (1) CB1 inhibition: human cannabinoid receptor-1 protein bound to Chem-1 membrane was used in modified HEPES (pH 7.4) buffer. A 10-μg aliquot of Chem-1 membrane, radioactively labeled [
3
H] CB1-ligand SR141716A (2 nM) (CB1 receptor antagonist), and test extracts or positive control R(+)-WIN-55,212-2 (10 μM) (CB1 receptor agonist) were incubated in Incubation Buffer (50 mM HEPES [pH 7.4], 5 mM MgCl2, 1 mM CaCl2, 0.2% bovine serum albumin [BSA]) for 90 min at 37°C. After incubation, the membranes were filtered and washed four times; then, the filters were counted to determine the amount of [
3
H] WIN-55,212-2 specifically bound to the CB1 membrane. The organic extracts and aqueous extracts obtained from different plant species and plant parts of M. alba such as root-barks, fruits, leaves, stem-barks, and stem were tested. Once activity was confirmed, the active plant extract was further fractionated and tested for inhibition of CB1-ligand binding activity by using the same method. Subsequently, active compounds were isolated, purified, identified, and tested for inhibition of CB1-ligand activity. The purified compounds Kuwanon G and Albanin G were tested at concentrations ranging between 0.04 and 20 μg/mL, to obtain a dose-response curve for each compound. The sample concentration was plotted against the percent inhibition, and the IC50 (defined as the concentration at which 50% inhibition of receptor binding activity is achieved in relation to the control) was determined. Inhibition of CB2 receptor-ligand binding activity of the purified Kuwanon G and Albanin G was also examined by using methods similar to those described in Example 2 for the CB1 receptor, with some modifications. (2) CB2 inhibition: Briefly, human cannabinoid CB2 receptor protein expressed in CHO-K1 cells was used in modified HEPES buffer (pH 7.0). A 30-μg aliquot of CB2 membrane was mixed with tritium-labeled nonspecific CB1 agonist [
3
H] WIN-55,212-2 (2.4 nM) and test samples of Kuwanon G and Albanin G, or just the nonspecific ligand R(+)-WIN-55,212-2 (10 μM) (positive control) were incubated in incubation buffer (20 mM HEPES [pH 7.0], 0.5 mg/mL BSA) for 90 min at 37°C. After incubation, the membranes were filtered and washed; the filters were then counted to determine the amount of radiolabeled [
3
H] WIN-55,212-2 that was specifically bound to the CB2 membrane. (3) α-Glucosidase inhibition: Inhibition of rice α-glucosidase activity by each of the purified compounds Kuwanon G and Albanin G was measured as follows. Rice α-glucosidase inhibition assay: The test compound or vehicle was preincubated with 77 mU/mL enzyme rice α-glucosidase in 2-(N-morpholino)ethanesulfonic acid buffer pH 6.3 for 15 min at 25°C. The reaction was initiated with addition of 2 mM p-nitrophenyl α-
Acute food intake study
Male Sprague-Dawley rats (Koatech, Korea), 8 weeks of age at the beginning of the experiment, were used in this study. During acclimation periods, the animals were maintained on a regular rat chow diet (2018S; Harlan, USA). The rats were housed in a climate-controlled room maintained on a 12 h/12 h reverse light/dark cycle. The rats were administered 0.5% CMC (carboxymethyl cellulose) aqueous solution as vehicle or in combination with caffeine at 40 mg/kg and M. alba extract at 250 and 500 mg/kg 30 min before the start of the dark-phase feeding cycle. Experimental testing commenced at the onset of the 12 h dark cycle. Before initiation of a test cycle, the rats were fasted overnight (less than about 16 h) to enhance their hunger. Otherwise, the animals had unlimited access to 45% HFD (Harlan) and tap water. Food intake was measured at 0, 1, 2, 4, 6, 8, 10, and 24 h from the start of the experiment to determine the acute food intake of each animal in a study. Eight animals per group were used for each treatment.
Diet-induced obesity induction and intervention
C57CL/6J mice aged 4–6 weeks (Korea Research Institute of Bioscience & Biotechnology, Ohchang, Korea) were housed in Polycarbonate cages (five mice per cage) in a room with a 12 h/12 h light/dark cycle and an ambient temperature of 24°C. All the mice were fed a commercial chow diet for 1 week after arrival in the animal facility. The mice were then divided into normal and obesity groups and fed with normal diet (ND) and HFDs, respectively. Once induction was confirmed, the HFD group was divided into (1) normal control + vehicle (N = 10, ND, research diet D12450B), (2) HFD + vehicle (N = 10), (3) HFD + Orlistat (N = 10, 40 mg/kg), (4) HFD + M. alba (N = 10, 250 mg/kg), and (5) HFD + M. alba (N = 10, 500 mg/kg); then, fractionated dosage administrations were initiated orally twice per day to give the full daily dosages for each group and sustained for 7 weeks. The vehicle-treated animals received 0.15% xanthan gum +0.5% Tween 80 only in both studies. The positive control Orlistat (Lipidown Cap 120 mg, Lot #12003; Hanmi, Korea), N-Formyl-
Statistical analysis
All nondiscrete data from clinical chemistry, body weights, and food consumption were represented as mean ± standard deviation and were analyzed by using SigmaPlot (version 11.0). Statistical significance between groups was calculated by means of single-factor analysis of variance followed by a paired test. P-values less than or equal to 0.05 (P ≤ .05) were considered significant. When normality tests failed, data for nonparametric analysis were subjected to Mann–Whitney sum ranks for test and Kruskal–Wallis one-way analysis of variance on ranks for ANOVA.
Results
CB1 antagonist activity
As seen in Table 1, the greatest inhibition of CB1-ligand binding activity was found in an organic extract obtained from root-barks of M. alba, with 91% inhibition at a testing concentration of 100 μg/mL and 82% at a testing concentration of 10 μg/mL (Table 1). These data show that organic extracts of M. alba root-bark contained moderately high levels of active compounds that block the binding of the labeled CB1-ligand to a human CB1 receptor protein. In contrast, other extracts from M. alba's different plant parts (fruits, leaves, stem bark, and stems) were tested in this experiment and showed undetectable levels of inhibition of CB1-ligand binding to the CB1 receptor. After subsequent activity-guided fractionation, isolation, and purification, the active compounds from the root-barks of M. alba were identified as Diels-Alder adduct of a chalcone and prenylphenyl moiety called Kuwanon G and Albanin G (Fig. 1 and Table 2) with 92% and 96% CB1-ligand binding inhibition activity, respectively. IC50 for the CB1/CB2 was found as 10.1/0.1 and 28.9/6.6 μM for Kuwanon G and Albanin G, respectively. There was no effect of α-glucosidase activity by the Kuwanon G, whereas the Albanin G showed 66% inhibition of this enzyme (Table 2).
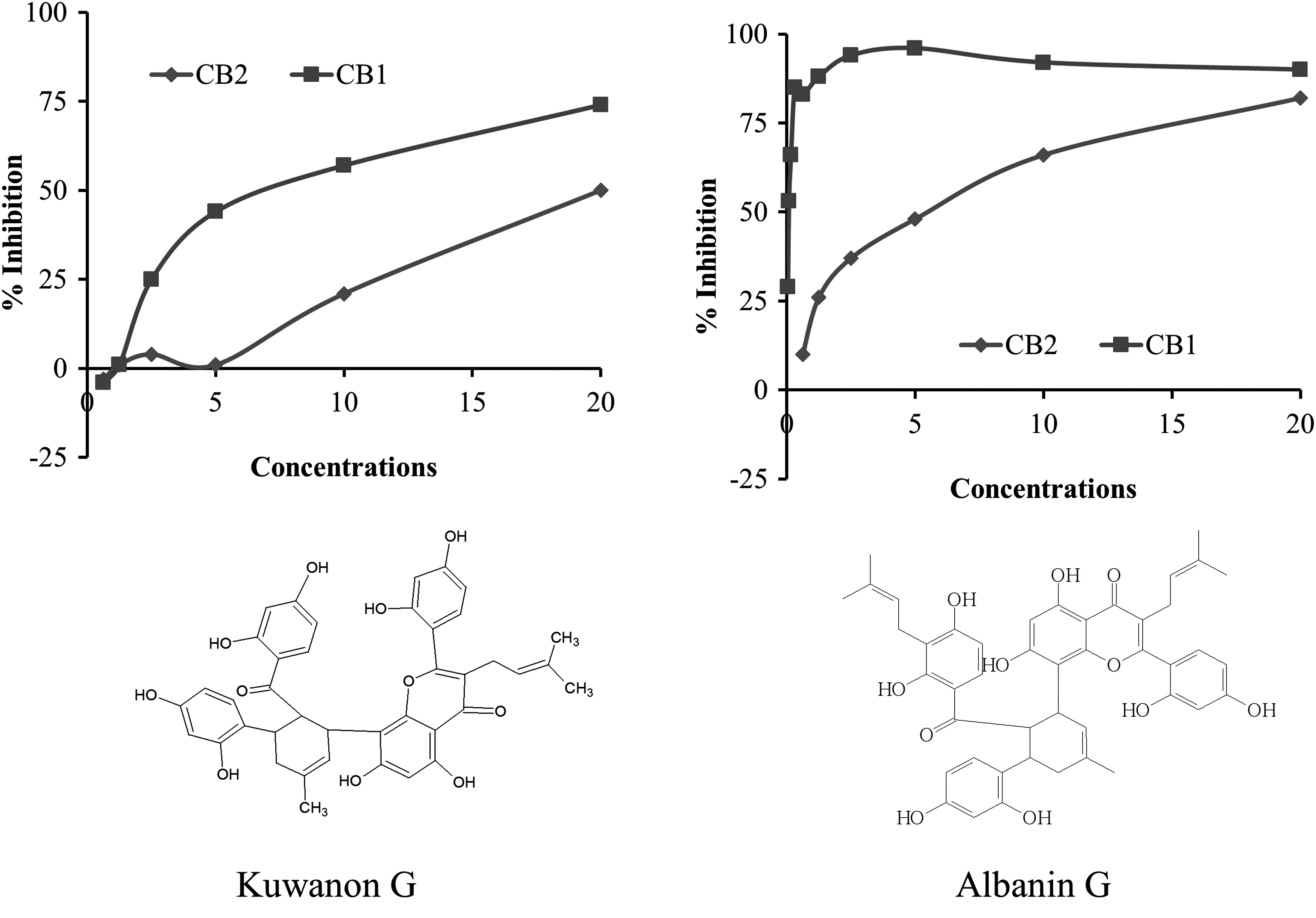
Active compounds from the root-barks of Morus alba and their CB1-ligand binding inhibition activity.
Inhibition of CB1-Ligand Binding Activity by Extracts from Various Parts of Morus alba Plant
Results are expressed as a percent inhibition relative to control radiolabeled SR141716A ligand.
OE means ground plant material extracted with Methanol:Dichloromethane (1:1 volume ratio).
AE means OE-extracted plant material that was air dried and extracted once with ultrapure water, filtered, and freeze-dried to provide aqueous extract material.
EE means that the ground plant material was extracted with only ethanol.
ME means that the ground plant material was extracted with only methanol.
Inhibition of CB1, CB2, and α-Glucosidase by Kuwanon G and Albanin G Purified from Morus alba Root-Bark Extracts
Food intake inhibition activity from acute model
Rats treated with M. alba extract showed dose-dependent reduction in food consumption. When cumulative food intakes were compared against the control animals treated with the vehicle, there were statistically significant reductions in food intake for Morus extract at both 250 and 500 mg/kg groups for the first 2 h (Fig. 2). These reductions were 58.6% and 44.8% for the 250 mg/kg and 50.1% and 44.3% for the 500 mg/kg (Fig. 2), respectively. It seemed that for the first hour the lower dosage performed better than the higher dosage of Morus root-bark extract. However, starting from the fourth hour postfood provision, the reductions in food intake were higher for the 500 mg/kg treatment group. Reductions of 40.1%, 45.5%, 40.3%, 39.1%, and 25.8% were found for rats treated with the 500 mg/kg of M. alba extract at 4, 6, 8, 10, and 24 h, respectively, after food provision (Fig. 2). In comparison, the 250 mg/kg treated animals showed 32.9%, 36.4%, 34.1%, 27.4%, and 10.0% reductions in food intake for the same time points monitored, respectively (Fig. 2).

Acute food intake inhibition activity of M. alba in Sprague-Dawley rats. Food intake was measured at 0, 1, 2, 4, 6, 8, 10, and 24 h. Eight animals per group were used for each treatment. *P ≤ 0.05, **P ≤ 0.001, † P ≤ 0.0001.
Effect of Morus root-bark extract on body weight in diet-induced obesity model
A progressive and stable body weight increase was observed when C57BL/6J mice (five mice/cage) were provided with a 60% Kcal HFD ad libitum for 6 weeks. The mice were transferred to a “minicage” that houses a single mouse per cage as of week 6 for additional 2 weeks of induction and the duration of treatment on the same diet. After 8 weeks on the HFD, a 52.1% increase in body weight gain (28.2 ± 2.0 g vs. 43.4 ± 2.4 g mean body weight of mice before and after HFD induction) was observed and deemed mice were ready for randomization and treatment interventions (Fig. 3). As depicted in Figure 3, as expected, a rapid drop in body weight was observed for mice treated with 40 mg/kg/day of obesity drug Orlistat for the first 2 weeks of the treatment period followed by a moderate body weight gain compared with the vehicle-treated HFD group. A similar trend was observed for mice treated with the 250 mg/kg/day of M. alba root-bark extract. A body weight decrease for the first 3 weeks and slight increases in body weight for the rest of the duration of the treatment were observed in mice treated with the 250 mg/kg of Morus extract. On the other hand, after 3 weeks of oral treatment, the body weight gains of obese mice treated with 500 mg/kg/day of Morus extract were on hold for the rest of the treatment duration. Compared with vehicle-treated HFD obese mice, body weight gains for mice treated with the 500 mg/kg/day of Morus root-bark extract were significantly lower as of week 1 of treatment and these differences remained statistically significant for the rest of the treatment duration (Fig. 3). The 500 mg/kg treatment group showed superior reduction in body weight gain compared with any of the other treatment groups, whereas the 250 mg/kg group showed comparable reduction in body weight gain compared with that of the positive control Orlistat. The changes in body weight gains at week 7 were found to be 0.45 ± 0.93, 2.87 ± 2.50, −1.42 ± 1.26, −0.28 ± 2.76, and −7.07 ± 1.58 g for lean mice under normal control diet, for obese mice under HFD, 40 mg/kg Orlistat, 250 mg/kg Morus extract, and 500 mg/kg Morus extract-treated animals, respectively (Table 3).
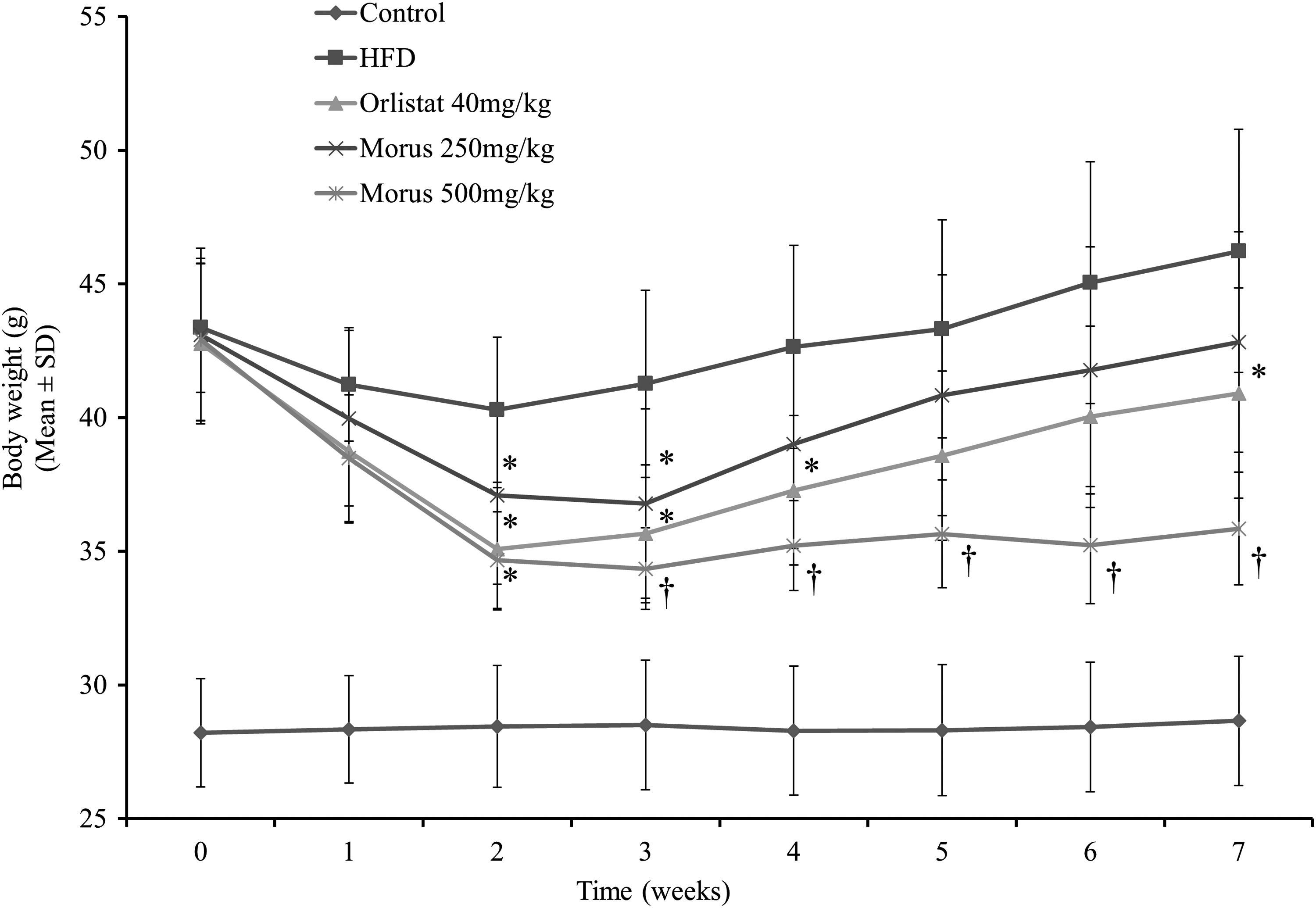
Effect of Morus root-bark extract on body weight in diet-induced obesity model. Mice (N = 10) were treated with the extract at oral doses of 250 and 500 mg/kg for 7 weeks. *P ≤ 0.05, † P ≤ 0.0001.
Effect of Morus alba Root-Bark Extract on Changes in Mice Fed a High Fat Diet in Body Weight Gain in Grams in Diet-Induced Obesity Model
HFD, high fat diet; ND, normal diet; SD, standard deviation.
Effect of Morus extract on food intake and efficiency in diet-induced obesity
The data depicted in Table 4 showed that both the M. alba root-bark extract treatment groups (250 and 500 mg/kg) and the positive control treatment group (Orlistat 40 mg/kg) showed statistically significant decreases in average body weight gain per day when body weight gain was analyzed on a per day basis. In agreement to the acute food intake data, mice treated with the 500 mg/kg Morus extract showed a statistically significant decrease in the average amount of food intake per day when compared with the HFD-induced obese group. Feed efficiency ratio [feed efficacy ratio = body weight gain (g/day)/food intake (g/day)] was statistically significant for both dosages of Morus extract and Orlistat when compared with the vehicle-treated HFD-induced obese animals.
Effect of Morus alba Root-Bark Extract on Food Intake and Efficiency in Diet-Induced Obesity Model
FER = body weight gain (g/day)/food intake (g/day).
P-value: compared with HFD by t-test. * P ≤ .05, ** P ≤ .001, † P ≤ .0001. Data expressed as mean ± SD.
FER, food efficiency ratio.
Effect of Morus extract on absolute organ weight in diet-induced obesity model
Decreases in visceral fat and liver weights were observed for mice treated with the 500 mg/kg Morus extract compared with vehicle-treated HFD-induced obese mice where most of the fat reductions were contributed by low mass in the perirenal fat (Table 5). These percentage reductions were determined as 28.3%, 35.2%, and 55.8% for the epididymal, retroperitoneal, and perirenal fat for animals treated with 500 mg/kg Morus extract in comparison with the vehicle-treated HFD-induced obese group. No differences in liver weight or other tissue weight were observed for the 250 mg/kg Morus extract and the positive control group. Statistically significant differences in liver weight, epididymal fat, retroperitoneal fat, perirenal fat, and total fat were observed for the lean animals under ND compared with the obese animals under HFD (Table 5).
Effects of Morus alba Root-Bark Extract on Absolute Organ Weight in Mice Fed a High Fat Diet
P-value: compared with HFD by t-test, * P ≤ .05, ** P ≤ .001, † P ≤ .0001. Data expressed as mean ± SD.
Effect of Morus extract on the metabolic biomarkers in the diet-induced obesity model
Disturbance in the normal metabolism process was confirmed here again, with a statistically significant increase in fasting serum levels of glucose, total cholesterol, and LDL-cholesterol in the HFD-induced obese animals compared with the normal control lean animals (Table 6). These increases were 101.9%, 80.9%, and 81.6% in fasting glucose, total cholesterol, and LDL-cholesterol, respectively, in the HFD-induced obese group compared with the normal lean control. These changes were significantly reduced as a result of treatment with Morus root-bark extract at an oral dose of 500 mg/kg. Statistically significant reductions in serum total cholesterol and LDL-cholesterol were observed for obese mice treated with Orlistat (Table 6). The low-dose Morus extract showed statistically significant reduction in the serum LDL level when compared with vehicle-treated HFD-induced obese animals (Table 6).
Effect of Morus alba Root-Bark Extract on Biochemistry Parameters
P-value: compared with HFD by t-test, * P ≤ .05, ** P ≤ .001, † P ≤ .0001. Data expressed as mean ± SD.
LDL, low density lipoprotein.
Discussion
From the current obesity drug development trend, it can easily be inferred that it is possible to reduce body weight and reverse the metabolic disturbances of obesity by controlling calories intakes. The cannabinoid (CB1) receptor antagonists such as Rimonabant and its class of drugs incur their impact by blocking the effects of endogenous cannabinoids that are known to stimulate hunger and increase appetite. 18 –20 Unfortunately, the strong binding affinity from the prescription drug also induced psychological side effects that eventually led to withdrawal of Rimonabant from the obesity treatment. However, searching natural CB1 antagonists with less binding affinity and more peripheral than neurological effects may remain as a viable alternative approach to control the epidemic of obesity.
In this study, we depicted data from CB1 antagonist plant library screening, acute food intake model in rats, and HFD-induced obese mice treated with root-bark extract of M. alba. More than 8000 plant extracts from Unigen's Phytologix library were screened for inhibition of CB1-ligand binding activity. Among them, M. alba was determined as one of the most active plants in the library. Various parts (root-bark, fruits, leaves, stem-bark, and stems) of M. alba plant were extracted with organic solvent, aqueous solvent, ethanol, and methanol and those extracts were tested for inhibition of CB1-ligand binding activity. It was found that only the organic extract of root-barks of M. alba showed 91% and 82% inhibition at 100 and 10 μg/mL concentrations, respectively. No inhibition activity was observed for the extracts obtained from other parts of the plant. Organic extract of M. alba root-barks was further investigated by examining high-through-put (HTP) fractions of the organic extract on a reverse-phase C-18 column for inhibition of CB1 binding activity by labeled CB1-ligand. The activity profile of those HTP fractions indicated that more than one component in the fractionated M. alba organic extract contributed to the inhibition of CB1-ligand binding activity. Subsequent isolation and purification resulted in two potent compounds identified as Kuwanon G and Albanin G with CB1-ligand binding inhibition activity IC50 7 μg/mL (10 μM) and 0.08 μg/mL (190 nM), respectively. For comparison, the reference compound Rimonabant had an IC50 of 20 nM in this assay. Both compounds showed selectivity to the CB1 receptor, magnifying their involvement in appetite control. Besides, Albanin G could have some additional benefit in inhibiting the activity of the enzyme α-glucosidase, indicating its further advantage in reduction of calories derived from carbohydrates and, hence, improvement of diabetes similar to the benefit from Acarbose. 21
Significant appetite suppression activity of M. alba root-bark ethanol extract was observed in the acute food intake study. The root-bark extract of M. alba produced dose-correlated reduction in food intake in rats administered at single oral doses of 250 and 500 mg/kg. When cumulative food intake data were compared with the vehicle-treated control animals, rats treated with the 250 mg/kg Morus extract showed 58.6%, 44.8%, 32.9%, 36.4%, 34.1%, 27.4%, and 10.0% reductions in food consumption at 1, 2, 4, 6, 8, 10, and 24 h, respectively, after food provision. For the same durations of monitoring, there were 50.1%, 44.3%, 40.1%, 45.5%, 40.3%, 39.1%, and 25.8% reductions in food intake for the 500 mg/kg Morus extract treatment group. These data showed that at the low dose, Morus extract seemed to have significant appetite suppression impact in the early hours of food provision than the higher dose. However, as time goes by, starting from the fourth hour, efficacies from the higher dose were greater in magnitude for the rest of the duration of the study. Long-lasting appetite suppression activity up to 24 h was observed for rats treated with Morus extract at 500 mg/kg (i.e., 25.8% compared with vehicle control, *P ≤ .01). These results were in accordance with clinically effective appetite suppressants in this model. For instance, the clinically effective obesity drugs such as Sibutramine, Rimonabant, Belviq, Qsymia, and Contrave all reduced food intake when given orally at a single dose to lean rodents. 22 –27 More specifically, when 18-h fasted rats were treated with Rimonabant (selective CB1 antagonist) at oral doses of 10 mg/kg, 55%, 50%, 42.9%, 38.1%, and 14.1% reductions in food intake were observed at 1, 2, 4, 6, and 24 h postfood provision. 28 Similarly, the pharmaceutical anti-obesity drug Lorcaserin, a selective human 5-hydroxytryptamine-2C (5-HT)2C agonist, reduced acute food intake in rats administered orally at doses of 3–24 mg/kg. At the highest dose of Lorcaserin tested, 80%, 65%, 55%, and 25% inhibitions in cumulative food intakes were observed at 2, 4, 6, and 22 h, respectively, after the provision of food. 25 Therefore, based on data depicted in this study, it can be inferred that the appetite suppression activity of M. alba root-bark extract observed in this model could be partially explained by the presence of active components in the extract, with CB1 antagonist activity causing reduced calorie intake.
We also evaluated the effect of M. alba root-bark extract on obesity and its associated metabolic disturbances in HFD-induced obese mice administered for 7 weeks. M. alba root-bark extract significantly decreased the body weight of obese mice in a dose-dependent manner. Data depicted here showed that chronic oral administration of M. alba root-bark extract reduced food intake and weight gain in C57BL/6J obese mice induced by HFD, demonstrating the steady efficacy of M. alba root-bark extract. In this repeated-dosing study, 7.4% and 22.5% reductions in body weight gain were observed for obese mice treated with M. alba root-bark extract at doses of 250 and 500 mg/kg when compared with vehicle-treated HFD obese animals at week 7. Compared with week 0 (baseline or treatment start), there was also a 16.5% (loss of 7.1 g) reduction in body weight for rats treated with the 500 mg/kg M. alba root-bark extract for 7 weeks. The reductions observed as a result of treatment with the high dose of M. alba root-bark extract were statistically significant at each time point monitored with an average body weight loss of 0.154 ± 0.034 g/day for the duration of the study († P ≤ .0001 vs. HFD). In contrast, the vehicle-treated HFD obese animals gained 6.6% body weight with an average body weight gain 0.062 ± 0.054 g/day. The anti-obesity drug Orlistat and the 250 mg/kg M. alba root-bark extract-treated obese animals showed 4.3% and 0.6% reductions in body weight from their baseline, respectively. Interestingly the body weight reduction observed for the high dose of M. alba root-bark extract is comparable to that of the reduction from CB1-specific antagonist, Rimonabant. After 5 weeks of daily oral treatment with Rimonabant at 10 mg/kg/day, obese mice showed a 20% reduction in body weight compared with the vehicle-treated HFD animals (i.e., loss of 8 g, P < .01). 29 In comparison, obese mice treated with the high dose of M. alba root-bark extract had 10.4 g (22.5%) and 7.1 g (16.5%) loss in body weight compared with vehicle-treated HFD obese mice (at week 7) and baseline of the same group of mice, respectively.
In agreement with the acute feed intake data, here again, there was an average daily 20% decrease in calorie intake for obese mice treated with the high dose of M. alba root-bark extract compared with the vehicle-treated HFD obese animals, reflecting the appetite suppression activity of the extract. However, the marked acute hypophagia observed in the acute food intake study tends to weaken over the repeated administration through the course of 7 weeks. It is worth mentioning here that, although there was no difference in calorie intake for the low dose of M. alba root-bark extract compared with the vehicle HFD, obese mice treated with Orlistat showed a 13.4% increase in daily calorie intake. Consistent with our findings, it was reported that Rimonabant administered orally to diet-induced obesity (DIO) obese mice at oral doses of 10 mg/kg/day for 5 weeks resulted in transient reductions in daily calorie intake (48% for the first week compared with vehicle-treated HFD). 29
Moreover, the absolute organ weight data also showed statistically significant reduction in the epididymal, retroperitoneal, and perirenal fat deposits in the obese mice treated with the high dose of M. alba root-bark extract where the largest decrease (55.8%) occurred in the perirenal depots compared with vehicle-treated HFD obese animals. The Orlistat and the 250 mg/kg M. alba root-bark extract groups showed statistically nonsignificant 23.3% and 16.3% reduction in perirenal fat mass, respectively, when compared with vehicle-treated HFD obese animals. Comparable to our data, the weight loss observed in DIO mice treated with Rimonabant (10 mg/kg) for 5 weeks was associated with a 50% depletion of fat stores attainment due to the decrease in both the weight of abdominal fat pads and the total body fat content. 29 These findings augment the decrease in body weight gain observed in these mice treated with M. alba root-bark extract and could mainly be due to the result of reduced visceral fat accumulation.
The liver is one of the vital organs to be easily affected by the comorbidities of obesity, often grossly reflected by its size increase. In this study, there was a statistically significant 65.5% increase in the absolute weight of the liver in the obese mice treated with vehicle (**P ≤ 0.001 vs. normal control). These changes were significantly reduced (26.4% compared with vehicle-treated HFD) by the treatment of M. alba root-bark extract at an oral dose of 500 mg/kg for 7 weeks. These findings were also corroborated by the moderation of the dyslipidemia observed in the vehicle-treated HFD group. Obese mice treated with 500 mg/kg M. alba root-bark extract for 7 weeks showed 25.6%, 29.7%, and 53.8% reductions in fasting serum glucose, total cholesterol, and LDL-cholesterol, respectively, compared with the vehicle-treated HFD obese animals. These decreases from Morus high-dose group were statistically significant for all the biomarkers. Obese mice in the Orlistat group experienced 12.2%, 30.5% (P ≤ 0.05), and 52.7% (P ≤ 0.0001) reductions in fasting serum glucose, total cholesterol, and LDL-cholesterol, respectively, compared with the vehicle-treated HFD obese animals. These findings further give strong support to the in vivo efficacy of M. alba root-bark extract in moderation of metabolic disorders of HFD-induced obese animals.
Substantiating our findings, significant studies have been reported describing the importance of Morus plants in metabolic disorders such as diabetes and obesity. For instance, mulberry (Morus indica L.) leave extracts and M. alba leaf tea for hypoglycemic activities in diabetes patients 30,31 and for improving hyperglycemia and associated complications in diabetic rats, 30,32 ethanolic extract from M. alba leaves for decreasing body weight and adiposity, regulating hepatic lipid accumulation in diet-induced obese mice, 33 mulberry leaf for decreasing expression of white muscle adipocytokines in db/db mice, 34 phenolics from the root bark of M. alba for reducing blood glucose level in alloxan diabetic mice, 35 mulberry leaf and fruit extract for lowering body weight gain, plasma triglycerides, and lipid peroxidation levels, suppressing hepatic fat accumulation and reducing epididymal adipocyte size in HFD-induced obese mice 36 were some of the beneficial effects documented for this medicinal plant. Although this is the first time to report the CB1 antagonist activity as a likely mechanism of action of M. alba root-bark extract, previously, extracts from Mulberry leaf had been shown to increase adiponectin level of adipocytes, increase glucose uptake and GLUT4 translocation in adipocytes, 37 inhibit α-glucosidase activity, 38,39 and inhibit human and rat intestinal disaccharidase. 40 Therefore, collectively, elucidation of the active components in M. alba root-bark as a special type of bioflavonoids with possible mechanisms of action as CB1 antagonist could further provide significant contributions to understanding and using Morus plants for mitigating obesity, obesity-associated comorbidities, and providing other health benefits.
More recently, we have reported that M. alba root-bark extract combined with Y. mate leaf extract and M. officinalis stem-bark extract, yielding a composition known as UP601. 14,15 We observed statistically significant decreases by 9.1%, 19.6%, and 25.6% of body weight for mice treated with UP601 at 300, 450, and 600 mg/kg compared with vehicle-treated high-fat and high-fructose-induced obese animals at week 7, respectively. Similarly, dose-correlated reductions of total cholesterol, triglyceride, LDL-cholesterol, and fasting serum glucose were observed for UP601. In the DEXA scan analysis, body fat distribution was also found to be reduced by 31.6% for the mid-dose (450 mg/kg) UP601 group. In addition, up to an 89.1% reduction in mesenteric fat deposit was observed for UP601-treated rats in relative organ weight measurements; corroborating the observed decreased body weight could actually be a result of reduction in visceral fat. 15 These findings were also substantiated by the HFD-induced obese mice model. 14 Hence, it could be inferred that the appetite suppression, body weight, and metabolism management of UP601 could be partially explained by the presence of M. alba root-bark extract as CB1 antagonist in the standardized formulation.
There were some limitations in this study in that (1) identified pure compounds were not tested for their in vivo activity on calorie intake and body weight reductions, (2) the observed decrease in food intake cannot be determined whether as a result of suppression of appetite or increasing satiety, (3) lack of DEXA scan data for the body composition to support the visceral fat reduction analysis, and (4) liver histopathology data are unavailable to indicate the changes in fat accumulation in the liver as a result of the HFD or Morus extract treatment-related improvements. Nevertheless, promising preclinical results have been documented as a CB1 antagonist of M. alba root-bark extract, indicating its potential usage in appetite suppression and weight loss, which will be further evaluated in human clinical trials.
The current pharmaceutical drugs are considered potential adjunctive treatment to lifestyle modification. However, the lack of sustainable weight loss, cost, and adverse effects render the current pharmacotherapy weight management unsuccessful. Associated side effects such as cardiovascular risk for Sibutramine, psychiatric problems for Rimonabant, and oily spotting and flatulence for Orlistat are a few examples. Therefore, in light of the limited pharmaceutical drug choices and the socio-economic implications of the obesity pandemic, medicinal plants such as M. alba, which has a long history of safe human consumption, can be considered an alternative solution. In addition, the active extracts in Morus may also provide other additional health benefits beyond weight control, suggesting its beneficial application and affordability.
In summary, we have identified two bioflavonoid compounds (Kuwanon G and Albanin G) from M. alba root-bark extract with CB1 antagonist activity. The anorexic activity of Morus extract was demonstrated in the acute food intake model. Data from a long-term study also supported the appetite suppression activity of M. alba root-bark extract, which was found to be effective in reducing body weight, calorie intake and also improving metabolic disturbances in obese mice fed with HFD. These results demonstrated that M. alba root-bark extract enriched in the Diels-Alder adducts of a chalcone and prenylphenyl moiety standardized at bioflavonoids Kuwanon G and Albanin G could be used to control calorie intake, manage body weight, and improve metabolic syndromes.
Footnotes
Acknowledgments
The authors would like to express their gratitude to Dr. Ed Cannon, Dr. Wenwen Ma, Dr. Doug Bradley, Dr. Min Chu, Dr. Eu-Jin Hyun, Mr. Tae-Woo Kim, and the Unigen team for their incalculable support for the completion of this project. The methods and the findings described in this article have been published under the granted U.S. patent (Grant No. 09844576) titled “Compositions and methods for managing weight”
Author Disclosure Statement
All authors are current Unigen employees; therefore, they have competing financial interests.