
Abstract
Glioblastoma multiforme (GBM) is one of the most aggressive and lethal human brain tumors, and the median survival of patients with GBM is only 14 months. Glioblastoma stem cells (GSCs) are regarded as a main cause of GBM recurrence, because of their self-renewal and drug resistance properties. Therefore, targeting GSCs is an important therapeutic strategy for GBM. In this study, we show the effects of BRM270, a compound from natural plant extracts, on GSCs in vitro and GBM recurrence in vivo. BRM270 induced apoptotic cell death and inhibited cell growth and “stemness” both in vitro and in vivo. Combining BRM270 treatment with concurrent chemoradiotherapy (CCRT) dramatically increased mice survival and tumor growth inhibition. Taken together, our results suggested that BRM270 synergizes with CCRT as a therapeutic agent to target GSCs.
Introduction
G
Natural phytochemicals have recently gained attention as a potential source of anticancer agents. Since existing antitumor drugs target rapidly dividing cells, some normal proliferating cells are also affected, consequently leading to undesired side-effects. However, natural compounds from plants are non-toxic to normal cells and their antitumor effects have been reported in many human malignancies, such as breast cancer, 8 liver cancer, 9 and colorectal cancer. 10 Interestingly, plant extracts are reported to inhibit several signaling pathways that are characteristic of stem cells, such as signal transducer and activator of transcription 3, nuclear factor-kappa B (NF-κB), c-MYC, and Wnt pathways. 11 –14 BRM270 is a compound from natural plant extracts that exhibits anticancer therapeutic effects on osteosarcoma and lung adenocarcinoma cells. 15,16 In these studies, BRM270 effectively inhibited NF-κB signaling in CD133-positive stem-like cancer-initiating cells. 15 However, the antitumor effects of BRM270 on GBM, especially on GSCs, remain undetermined.
In this study, we examined the effect of BRM270 on GSCs both in vitro and in vivo and found that BRM270 suppressed GSC growth and stemness properties in vitro and effectively diminished GBM recurrence after concurrent chemotherapy and radiotherapy in vivo.
Materials and Methods
Cell culture and reagents
Normal human astrocytes (ASCR) obtained from the National Cancer Center were maintained in Dulbecco's modified Eagle medium that was supplemented with 10% fetal bovine serum (Biotechnics Research, CA, USA), 1% penicillin/streptomycin (Lonza, Basel, Switzerland), and 2 mM L-glutamine (Lonza). MD13 GSCs obtained from Dr. Ichiro Nakano were cultured in a neurobasal medium (Invitrogen, CA, USA) that was supplemented with modified B27 (0.04%; Invitrogen, MA, USA), epidermal growth factor (EGF, 20 ng/mL; R&D Systems, MN, USA), and basic fibroblast growth factor (bFGF, 20 ng/mL; R&D Systems). 16 In some cases, ASCRs were also pre-cultured for 12 h in a neurobasal medium that was supplemented with modified B27, EGF, and bFGF before treatment with BRM270. Cells were treated with freeze-dried BRM270 powder that was dissolved in phosphate-buffered saline (PBS) at the indicated concentrations. 15,17
Hematoxylin and eosin staining and immunohistochemistry
Tumor-bearing mice were perfused with PBS and 4% paraformaldehyde. The obtained tumor tissues were embedded in paraffin, sectioned (4 μm in thickness), and placed on glass slides. After deparaffinization and hydration, tissue slides were treated with hematoxylin (1051740500; Merck, Darmstadt, Germany) for 5 min and rinsed with tap water. All slides were incubated in an eosin solution (109844; Merck) for 30 sec and then washed with distilled water.
After deparaffinization and hydration, tissue slides were stained with primary antibodies against Ki67 (NCL-Ki67p; Leica Biosystems, UK), cleaved Caspase 3 (9661; Cell Signaling, MA, USA), CD15 (559045; BD Biosciences, CA, USA), and CD44 (sc-7297; Santa Cruz Biotechnology, TX, USA) for 12 h at 4°C. Slides were incubated with a biotin-conjugated secondary antibody (Vector Laboratories, CA, USA) for 1 h and with VECTASTAIN ABC Reagent (PK-4000; Vector Laboratories) for 30 min at room temperature, respectively. Slides were incubated in peroxidase substrate solution (PK-4100; Vector Laboratories) and counterstained with hematoxylin. Stained images were obtained by using an Epifluorescence and Brightfield microscope (Axioimager M1; Carl Zeiss, MA, USA), pseudo-colored, and quantified with MetaMorph software (Molecular Devices, CA, USA).
In vivo orthotopic implantation and CCRT
For orthotopic implantation, 3 μL of 1 × 102 MD13 GSCs was stereotactically injected into the nude mice brains; coordinates: 2 mm right and 0.5 mm anterior of the bregma. 18 The mice received daily oral administration of BRM270 (1 g/kg) or PBS. For CCRT, 137Cs γ-ray irradiation at a dose rate of 2.04 Gy/min for a total dose of 2 Gy was conducted by using an IBL 437C (CIS Bio-International, Codolet, France). Anesthetized mice were placed in a lead shielding device, and whole brains were exposed to fractionated irradiation (5 × 2 Gy) followed by oral administration of temozolomide (10 mg/kg) or PBS from 10 to 14 days after orthotopic injection.
To analyze the mouse survival, the mice that lost more than 30% of their body weight were sacrificed. Overall survival curves were presented by the Kaplan–Meier method. To compare the tumor histology, all the mice were simultaneously sacrificed when one mouse first showed neurological symptoms. All mouse experiments were approved by the Animal Care Committee of Korea University according to government and institutional guidelines and regulations of the Republic of Korea.
Quantitative reverse transcription-polymerase chain reaction
Total RNA was isolated from cells, using QIAzol lysis Reagent (Qiagen, Hilden, Germany), according to the manufacturer's instructions. RNA (1 μg) pre-treated with RNase-free DNase was utilized as a template for synthesizing complementary DNA (cDNA), using the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, MA, USA), according to the manufacturer's instructions. The quantitative reverse transcription-polymerase chain reaction (qRT-PCR) analysis was performed by using Takara Bio SYBR Premix Ex Taq and CFX096 (Bio-Rad, CA, USA). The expression level of each target gene was normalized to the expression of 18S ribosomal RNA (rRNA).
The primer sequences were human 18S rRNA forward: 5′-CAGCCACCCGAGATTGAGCA-3′, reverse: 5′-TAGTAGCGACGGGCGGTGTG-3′; human CD44 primer forward: 5′-AGAAGGTGTGGGCAGAAGAA-3′, reverse: 5′- AAATGCACCATTTCCTGAGA-3′; human NESTIN primer forward 5′-AGAGGGAGGACAAAGTCCCT-3′, reverse: 5′-CACTTCCTCAGACTGCTCCA-3′.
In vitro limiting dilution assay
For an in vitro limiting dilution assay, sequentially decreasing numbers of cells per well (20, 10, 5, and 1) were seeded in 96-multiwell plates and tumorsphere formation was observed in each well. Stem cell frequency was calculated by using software available at
Annexin-V and propidium iodide staining
MD13 GSCs were treated with PBS or BRM270 (500 μg/mL) for 3 days. Then, cells were collected and washed twice with PBS. The cells were incubated with Annexin-V-FITC (BD Biosciences, MA, USA) and propidium iodide (PI; BD Biosciences) for 15 min at room temperature (19–21°C) in the dark. Incubated cells were analyzed by fluorescence-activated cell sorting (FACS Verse; BD Biosciences).
Cell cycle analysis
For cell cycle analysis, cells (2.5 × 106) were washed with ice-cold PBS, treated with trypsin, and fixed in cold 70% ethanol at 4°C for at least 24 h. Then, fixed cells were washed twice in PBS, and they were incubated in PI and RNase A for 3 h at 4°C. Incubated cells were analyzed by an FACS Verse instrument.
MTS cell viability assay
Suspended cells (100 μL) were seeded at a density of 3000 cells per well in 96-multiwell plates. Then, cells were incubated with or without BRM270 at different doses. The 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay is a colorimetric assay for measuring cell viability. The MTS assay is the improvement assay of traditional 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay as one-step assay. The MTS assay was conducted with an Ez-Cytox Kit (DOGEN, Seoul, Korea) to measure cell viability. EZ-Cytox solution was added to each well and then, cells were incubated for 4 h. The light absorbance was determined at 450 nm by the PowerWaveXS microplate-reader (Bio-Tek Instruments, Inc., VT, USA).
Live cell imaging
Suspended cells (100 μL) were seeded at a density of 3000 cells per well in 96-multiwell plates. Cells were incubated in a humidified atmosphere at 37°C and 5% CO2 and treated with BRM270. The plate was imaged by using an Incucyte Zoom (Essen Instruments, MI, USA) every 2 h for 3 days. The images were obtained under phase-contrast microscopy and pseudo-colored by software installed in Incucyte Zoom.
Statistical analysis
Statistical analysis was performed by a two-tailed Student's t-test. Values of P < .05 (*) or P < .01 (**) were considered statistically significant for different experiments, as indicated in all figures. Data are presented as means ± standard error of the mean (SEM).
Results
BRM270 specifically inhibits viability and growth of GSCs
Since GSCs are known to be a main cause of tumor initiation and recurrence after standard therapy, we evaluated the effects of BRM270 on GSCs in vitro. Using GSC MD13 and ASCR, we performed an MTS cell viability assay on treating cells with different concentrations of BRM270. The results showed that BRM270 inhibited viability of MD13 GSCs more specifically in a dose-sensitive manner than it inhibited control ASCR cells (Fig. 1A). The half-maximal inhibitory concentration (IC50) of BRM270 in MD13 GSCs was 594.9 μg/mL, whereas the IC50 in ASCR cells was 3884 μg/mL (Fig. 1A). Using a live-content imaging system, we also examined changes in cell morphology on treatment with BRM270. Then, MD13 GSCs treated with BRM270 formed smaller tumorspheres than non-treated MD13 GSCs (Fig. 1B and Supplementary Fig. S1; Supplementary Data are available online at
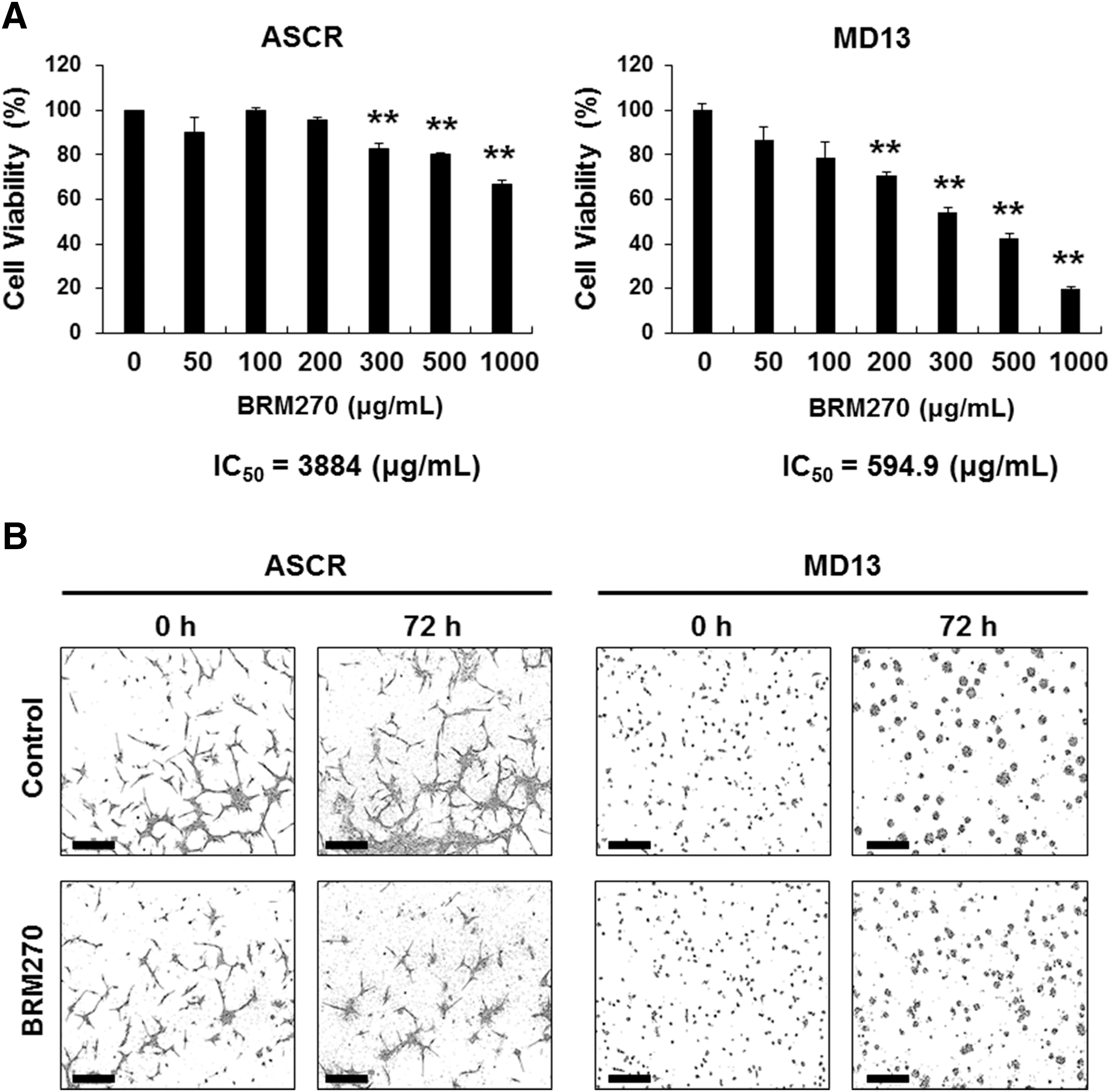
BRM270 inhibits GSC viability and growth.
BRM270 promotes GSCs apoptosis
To identify the cause of growth inhibition, we performed cell cycle analysis in the GSCs treated with BRM270. The results showed that BRM270 significantly increased sub-G0 populations, whereas there were no significant differences in G1, S, or G2/M populations (Fig. 2A). Because the sub-G0 phase indicates dead cell populations, we further determined whether BRM270 induced apoptotic cell death of GSCs. We stained the MD13 GSCs with Annexin-V and PI after BRM270 treatment. FACS analysis showed that BRM270 remarkably promoted apoptotic cell death of MD13 GSCs (Fig. 2B; 15.04% Annexin-V and PI double-positive apoptotic cell death). Taken together, these results indicate that BRM270 induces apoptotic cell death in vitro.
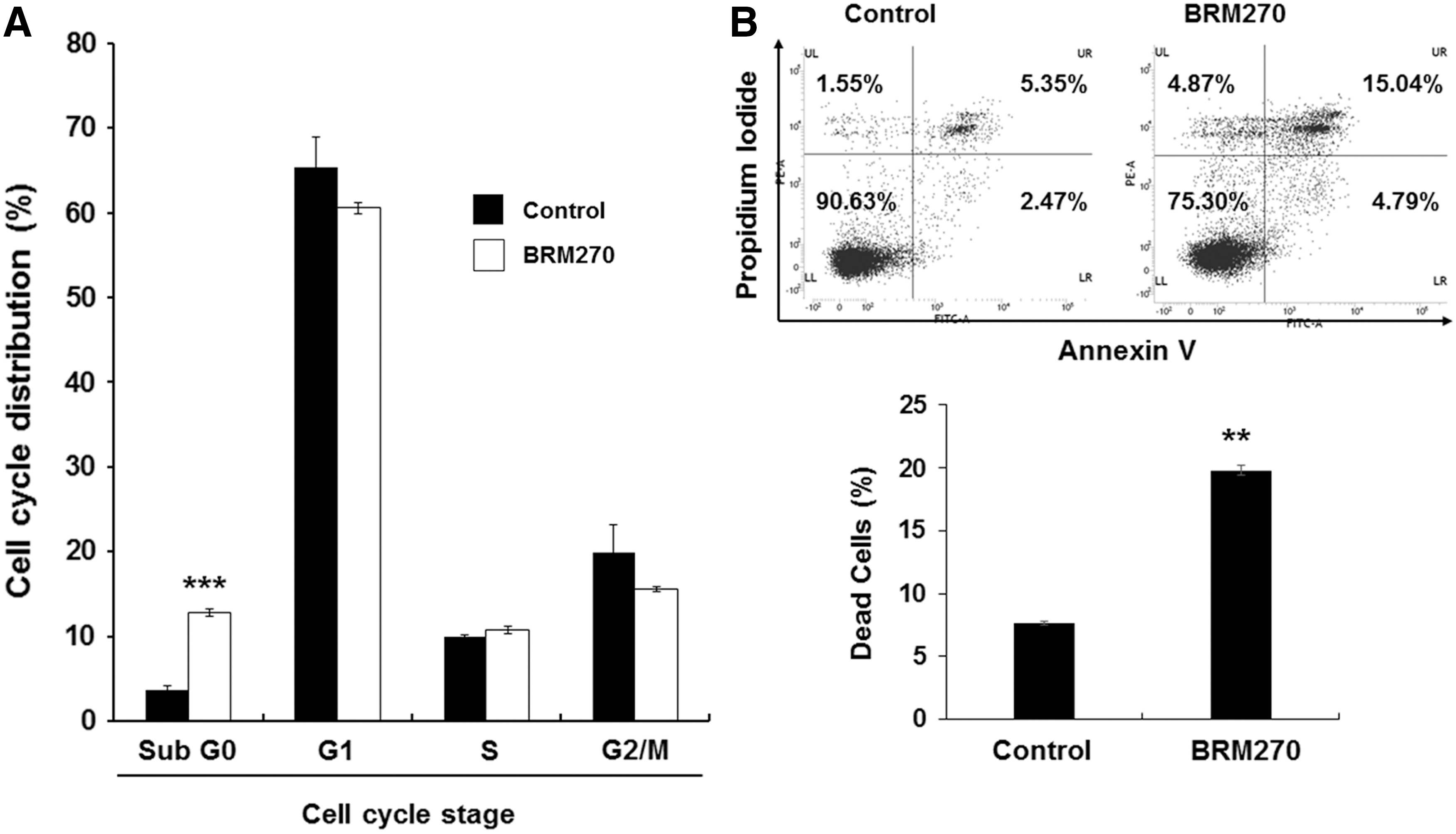
BRM270 promotes GSC apoptosis.
BRM270 suppresses stem cell markers and tumorsphere-forming ability in GSCs
There are several markers available to evaluate GSC stemness. 19 Among them, the detection of stem cell marker and tumorsphere-forming assay have been widely used to identify GSC stemness. Therefore, to evaluate whether BRM270 regulated GSC stemness properties, we examined stem cell markers expression and tumorsphere-forming ability. The results showed that treatment with 300 μg/mL BRM270 daily (D300) or every 72 h (T300) for 7 days dramatically decreased messenger RNA levels of stem cell markers, such as CD44 and NESTIN, in MD13 GSCs (Fig. 3A). In addition, we performed an in vitro limiting dilution assay to investigate the effect of BRM270 on GSC tumorsphere-forming ability. As a result, BRM270 treatment dramatically suppressed tumorsphere-forming ability of MD13 GSCs (Fig. 3B). Taken together, these results show that BRM270 inhibits stemness properties of GSCs.

BRM270 inhibits GSC stemness properties.
Combined treatment of BRM270 with CCRT inhibits GBM recurrence
To determine the antitumor effects of BRM270 in vivo, we generated a GBM xenograft model by injecting MD13 GSCs into brains of nude mice. Then, we treated the tumor-bearing mice with BRM270 (1 g/kg daily) alone, CCRT alone as a standard therapy for patients with GBM, 20 or a combination of BRM270 and CCRT. Unlike the in vitro data, there was no significant difference in survival between mice treated with PBS and mice treated with BRM270 alone (Fig. 4A). Mouse survival was significantly extended by treatment with CCRT alone compared with control or BRM270 treatment (Fig. 4A). Surprisingly, mouse survival was dramatically increased in the group receiving the combined treatment of BRM270 and CCRT compared with other groups (Fig. 4A). To examine differences in tumor growth among the four groups, we sacrificed the mice at the same time (at day 21) when control mice exhibited neurological symptoms. Hematoxylin and eosin (H&E) staining showed that tumors of mice treated with BRM270 presented decreased tumor growth compared with tumors of control mice (Fig. 4B, upper panel). In addition, all mice receiving CCRT exhibited notably reduced tumor growth at day 21. Interestingly, mice receiving combined treatment of BRM270 and CCRT exhibited no tumor recurrence at day 21. In fact, five out of six tumor-bearing mice receiving the combined treatment of BRM270 and CCRT exhibited complete tumor growth inhibition at the end of the animal study (at day 55) (Fig. 4B, bottom panel). Even the one mouse with recurrent GBM after combined treatment of BRM270 and CCRT showed decreased tumor growth compared with tumor growth of mice treated with CCRT alone (at day 30) (Fig. 4B, middle panel).
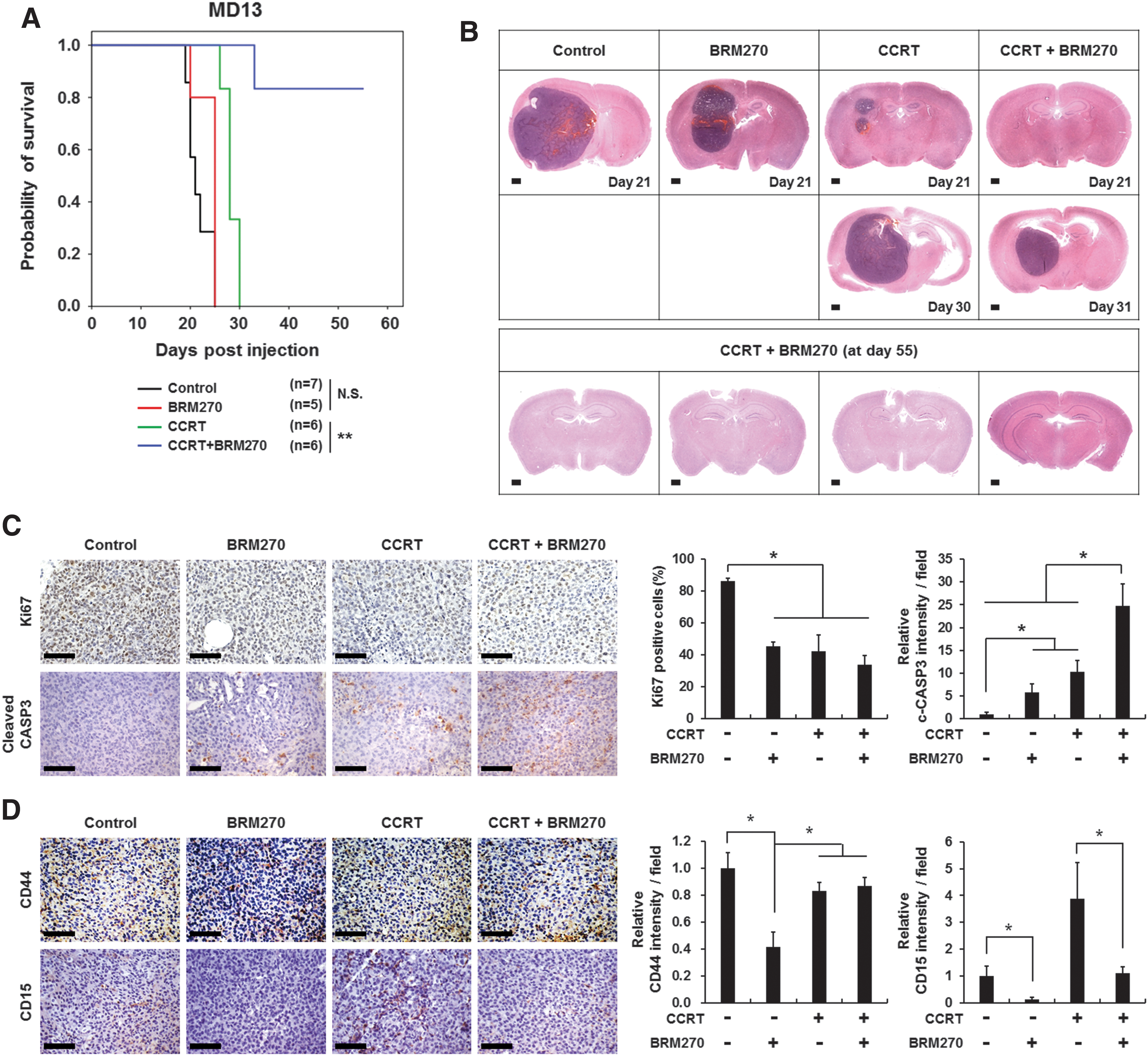
Combined treatment of BRM270 and CCRT suppresses GBM recurrence.
We next examined proliferating cell populations that were positive for Ki67 and apoptotic cell populations that were positive for cleaved Caspase 3 in tumor tissues by immunohistochemistry analysis. Ki67-positive cells were significantly decreased, whereas cleaved Caspase 3-positive cells markedly increased in tumors of mice receiving single treatments of BRM270 and CCRT or the combined treatment of BRM270 and CCRT (Fig. 4C and Supplementary Fig. S2A). However, the combined treatment of BRM270 and CCRT resulted in the most dramatic changes in Ki67 or cleaved Caspase 3 positivity compared with controls and single treatment. These results suggest that BRM270 treatment displays a synergistic effect when combined with CCRT treatment (Fig. 4C and Supplementary Fig. S2A).
Next, we performed immunohistochemistry analysis to determine cell populations expressing the cancer stem cell markers CD44 and CD15. CD44 expression decreased in tumors of mice receiving BRM270 or CCRT alone or the combined treatment of BRM270 and CCRT (Fig. 4D and Supplementary Fig. S2B). However, the most significant decrease in the number of CD44-positive cells was observed in tumors of mice treated with BRM270 alone (Fig. 4D and Supplementary Fig. S2B). BRM270 treatment alone also most significantly reduced the number of CD15 positivity compared with the other treatments. It is noteworthy that CCRT treatment alone markedly increased CD15 levels, possibly because of an enrichment of GSC populations that are resistant to CCRT. 4 It is plausible that BRM270 administration effectively reduced the number of CD15-expressing GSCs that are enriched by CCRT treatment (Fig. 4D and Supplementary Fig. S2B). Taken altogether, our results indicate that consistent with the stemness-inhibiting effects of BRM270 that we observed in vitro, BRM270 effectively diminished GSC populations in vivo.
Discussion
Despite the standard therapy, most patients with GBM experience an inevitable relapse and have a median survival time of less than 14 months. 1,20,21 The main cause of tumor recurrence is attributed to GSCs that have stemness properties and are resistant to CCRT. 6,22 Mechanistically, GSCs are known to have active DNA damage response pathways, allowing them to repair damaged DNA and survive after CCRT. 23,24 Therefore, targeting GSCs is considered an important therapeutic strategy. Here, we demonstrated that BRM270 increased apoptotic cell death and decreased stemness properties in GSCs, but not in normal cells, both in vitro and in vivo. Especially, combining BRM270 and CCRT treatment significantly improved the mouse survival and decreased tumor recurrence in a GBM xenograft model.
Although BRM270 alone increased apoptotic cell death and decreased stem cell marker expression both in vitro and in vivo, it did not significantly extend survival time in tumor-bearing mice. However, tumors treated with BRM270 alone were smaller in size than tumors in the control group after mice were sacrificed at the same time. Thus, we conclude that BRM270 alone can induce GSC apoptosis, but it is not sufficient to increase mouse survival in vivo. Interestingly, all mice treated with CCRT alone experienced tumor recurrence, whereas most mice treated with both BRM270 and CCRT in combination exhibited complete tumor growth inhibition. We assume that CCRT promoted cell cycle arrest and apoptosis of the proliferating tumor cells, but did not target GSCs, whereas BRM270 suppressed stemness properties and increased apoptosis in GSCs. Therefore, it is likely that the combined treatment of BRM270 and CCRT exhibits synergistic effects for eliminating GSCs.
BRM270 inhibits NF-κB signaling in CD133-expressing cancer-initiating cells, resulting in suppression of interleukin-6-mediated drug resistance. 15 NF-κB signaling is essential for tumor progression, resistance to CCRT, and secretion of several cytokines and chemokines. 25,26 Especially, secretome induced by NF-κB signaling plays important roles in accumulation of M2-like immune cells, which is known to promote tumor growth, invasion, and resistance. 27 Although we examined the effects of BRM270 on GSCs directly both in vitro and in vivo, BRM270 might contribute toward eradicating GSCs indirectly through modulating the tumor microenvironment. We found that BRM270 promoted microglial cell recruitment around apoptotic cells in the tumor tissue (data not shown). It is plausible that the recruited microglial cells in the tumor tissues treated with BRM270 alone or both BRM270 and CCRT combined might act through M1 type macrophages that exert an anti-tumorigenic function by attacking tumor cells and inducing cell death. 28,29 Thus, BRM270 might play a crucial role in suppressing the conversion of M1 type macrophages into the M2 type that exert pro-tumorigenic effects by providing several cytokines and chemokines. 30,31 Therefore, a further investigation related to the effects of BRM270 on the tumor microenvironment should be performed.
Footnotes
Acknowledgments
This work was supported by grants from the National Research Foundation funded by the Ministry of Science, ICT and Future Planning (2015R1A5A1009024), from the Next-Generation Biogreen 21 Program (PJ01107701), and from Korea University. The authors are grateful to all members of the Cell Growth Regulation Laboratory for their helpful discussions and technical assistance.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.