
Abstract
Morinda citrifolia (Noni) leaf is an herbal medicine with application in the domestic treatment of a broad range of conditions, including bone fracture and luxation. However, the basic mechanism underlying the stimulation of osteogenic differentiation by Noni leaf extract remains poorly understood. This study aimed to examine the effect of this extract on osteogenic differentiation and the mechanism by which Noni leaf extract enhances osteogenic differentiation. Aqueous extract of Noni leaves was prepared, and rutin and kaempferol-3-O-rutinoside were identified to be two of its major components. C2C12 and human periodontal ligament (hPDL) cells were used to study the effect of Noni. Noni did not show cytotoxicity at a concentration range of 0.015%–1.0% (w/v%) and significantly enhanced the activity of alkaline phosphatase (ALP) and expression levels of osteoblast differentiation markers, including Runx2, ALP, osterix, and osteocalcin, bone morphogenetic protein 2, Wnt3a, and β-catenin. In addition, Noni enhanced the matrix mineralization of hPDL cells. In the signaling pathways, Noni increased the phosphorylation levels of Akt and GSK3β and nuclear translocation and transcriptional activity of β-catenin, which were attenuated by the addition of Dkk-1, a Wnt inhibitor, or LY294002, a PI3K inhibitor. These results suggest that Noni leaf extract enhances osteogenic differentiation through the PI3K/Akt-dependent activation of Wnt/β-catenin signaling. Noni leaf extract might be a novel alternative medicine for bone and periodontal regeneration in patients with periodontal diseases.
Introduction
P
The primary treatment of chronic periodontal disease is to eliminate the cause through practicing oral hygiene or through dental interventions such as mechanical scaling and root planing with or without periodontal surgery. Although some progress has been reported in treating this disease, conventional periodontal therapies that can fully regenerate the lost bone and periodontal ligament are few and unstable. 5,6 As a result, various regenerative therapies such as membrane-guided tissue regeneration, osseous grafts, and the application of protein/chemicals/growth factors have been proposed. 7 These clinical approaches contribute to the regeneration of the periodontal tissue; however, several issues, including the properties of some osseous materials that prevent their absorption, sensitivity of techniques, unpredictability of outcomes, and limitations in the therapeutic indication, remain unsolved. 6,8 –11
Natural products have been of great importance for the treatment of diseases since ancient times. 12 Studies have shown the effectiveness of medicinal plants in treating diseases such as cancer, 13 Alzheimer's disease, 14 chronic obstructive pulmonary disease, 15 diarrhea and cholera secretions, 16 stomatitis, 17 and periodontal disease. 18 Morinda citrifolia (Noni) is one of the plants used in traditional medicine commonly found in Southeast Asia and the Western Pacific. 19 To date, more than 100 compounds in Noni fruits have been identified and found to have a broad range of therapeutic effects, including antibacterial, antiviral, antifungal, antitumor, antihelminthic, analgesic, anti-inflammatory, and immune-enhancing effects. 20 –22
Although the chemical composition and pharmacological properties of Noni juice and fruit have been considerably characterized, few studies have investigated the properties of Noni leaves. Noni leaf has been traditionally used for the topical treatment of burns, headaches, fever, and various types of inflammation, including bone fracture or dislocation. 19,23 Few scientific studies have confirmed the anti-inflammatory, antioxidant, 24 and wound healing enhancement 25 properties of Noni leaves.
In vitro osteogenic differentiation of human periodontal ligament (hPDL) cells is commonly induced by incubating cells with osteogenic supplements (dexamethasone, β-glycerophosphate, and ascorbic acid). We have previously demonstrated that Noni leaf water extract increased alkaline phosphatase (ALP) activity and matrix mineralization in hPDL and MC3T3-E1 cells without the addition of osteogenic supplements, suggesting that Noni leaves have a strong effect on the induction of osteogenic differentiation. 26 However, in that study, the highest dose of Noni leaf extract without cytotoxicity (2.5 w/v%) was used to confirm the osteogenic effect of Noni.
Taking into account the clinical application and safe use of Noni leaf extract, it is necessary to determine the minimum dose of the extract that can enhance hPDL differentiation and molecular mechanisms underlying its osteogenic activity. The activation of Wnt/β-catenin signaling promotes osteogenic 27,28 and cementogenic 29 differentiation, suggesting that Wnt/β-catenin signaling is of interest as a promising therapeutic target for bone and periodontal regeneration. Therefore, in the present study, we determined the minimal dose of Noni leaf extract that enhances osteogenic differentiation, and investigated whether Wnt/β-catenin signaling plays a role in the stimulatory effect of the Noni extract on the osteogenic differentiation of C2C12 and hPDL cells.
Materials and Methods
Materials
Alpha minimum essential medium (αMEM) and Dulbecco's modified Eagle's medium (DMEM) were purchased from Hyclone (Logan, UT, USA). Fetal bovine serum (FBS) was obtained from BioWhittaker (Walkersville, MD, USA). The ALP Staining Kit, LY294002, a phosphoinositide 3-kinase (PI3K) inhibitor; Alizarin Red, p-nitrophenyl phosphate, and p-nitrophenol were purchased from Sigma-Aldrich (St. Louis, MO, USA). Bone morphogenetic protein 2 (BMP2) and dickkopf-related protein 1 (DKK1) were purchased from R&D Systems (Minneapolis, MN, USA). The PCR primers were synthesized by Cosmo Genetech (Seoul, Korea) and Macrogen (Seoul, Korea). Lipofectamine™ reagent was purchased from Invitrogen (Carlsbad, CA, USA). Glycogen synthase kinase-3β (GSK3β), pGSK3β, Akt, and pAkt antibodies were obtained from Cell Signaling Technology (Beverly, MA, USA). β-catenin, β-actin, and lamin B antibodies, and goat anti-rabbit and anti-mouse horseradish peroxidase (HRP)-conjugated IgG were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Cy3-conjugated goat anti-mouse lgG was obtained from Jackson lmmunoResearch (West Grove, PA, USA). VECTASHIELD mounting medium for fluorescence with DAPI was purchased from Vector Laboratories, Inc. (Burlingame, CA, USA). The PRO-PREP™ was obtained from iNtRON Biotechnology (Sungnam, Korea), and the Supex reagent was ordered from Dyne-Bio (Sungnam, Korea). NE-PER Nuclear and Cytoplasmic Extraction Reagent was obtained from Pierce Biotechnology (Rockford, IL, USA). The T cell factor (TCF)/lymphoid enhancing factor-1 (LEF) reporter plasmid TOP-Flash was purchased from Upstate Biotechnology (Lake Placid, NY, USA). The Dual-Glo Luciferase Assay Kit was purchased from Promega (Madison, WI, USA).
Preparation of Noni leaf aqueous extract
Fresh Noni leaves were collected from Chonburi Province, Thailand. After being washed with water, air dried, and powdered with an electric blender, the leaf powder was imported to the Department of Molecular Genetics at the Seoul National University School of Dentistry in Korea. Noni leaf powder was suspended in sterilized distilled water at 4°C. After 24 h, the Noni leaf mixture was filtered using a fine muslin cloth and was then centrifuged at 14,240 g for 10 min at 4°C. The supernatant was filtered through a Whatman filter paper No. 1 to remove remaining debris, freeze dried with a lyophilizer (Lyph Lock6 Model 77595-01; Labconco, Kansas City, MO, USA), and stored at −80°C. We obtained ∼1.28 g of freeze-dried extract from 10 g of Noni leaf powder. Freeze-dried Noni leaf extract was dissolved in a culture medium, filtered through a 0.22-μm cellulose acetate membrane filter (Pall Corporation, MI, USA), and sterilized by UV light for 30 min before use. 26
Chemical profile of Noni extract
An Agilent 1260 HPLC system (Agilent Technologies, Santa Clara, CA, USA) equipped with a solvent degasser, quaternary pump, autosampler, and diode array detector was used in the current study. Agilent ChemStation® software was used for instrument operation control and data collection. HPLC was performed using a Luna® C18 column (250 × 4.6 mm; i.d., 5 μm; Phenomenex, Torrance, CA, USA) under the following conditions: flow rate 1.0 mL/min and injection volume of 10 μL, with the mobile phase comprising water and acetonitrile. The gradient elution for samples was as follows: 5%–30% (acetonitrile) in 30 min, 35%–100% (acetonitrile) in 5 min, and 100% (acetonitrile) in 5 min. Constituents of Noni extract were identified by comparing the HPLC retention time of target peaks with those of the standards. The peaks that match to those of rutin and kaempferol-3-O-rutinoside were further confirmed by UV spectra and LC-MS/MS analysis. The UV spectra were monitored by a photodiode array detector in the range of 190–450 nm and detection of rutin and kaempferol-3-O-rutinoside was performed at 350 nm.
Cell culture
C2C12 cells were maintained in DMEM supplemented with 10% FBS and antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin). To induce osteogenic differentiation, the culture medium was changed to DMEM supplemented with 5% FBS and 20 ng/mL of BMP2.
hPDL cells were purchased from ScienCell™ Research Laboratories (Human PDLFs; Carlsbad, CA, USA) and maintained in α-MEM supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin. hPDL cells passaged three to six times were used for the following experiments.
Quantitative reverse transcription–polymerase chain reaction
To evaluate mRNA expression, quantitative reverse transcription–polymerase chain reaction (qRT-PCR) was performed as previously described. 30 Each sample was analyzed in triplicate, and target genes were normalized to the housekeeping gene, glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Fold differences were then calculated for each treatment group using normalized C T values for the control.
The primer sequences used for qRT-PCR of mouse genes were as follows: Wnt3a (f) 5′-CAT GCA CCT CAA GTG CAA ATG-3′ and (r) 5′-TGA GGA AAT CCC CGA TGG T-3′; Ctnnb1 (β-catenin gene) (f) 5′-GGT GCT GAC TAT CCA GTT G-3′ and (r) 5′-GGC AGA GTA AAG TAT TCA CCC-3′; Runx2 (f) 5′-TTC TCC AAC CCA CGA ATG CAC-3′ and (r) 5′-CAG GTA CGT GTG GTA GTG AGT-3′; Alpl (ALP gene) (f) 5′-CCA ACT CTT TTG TGC CAG-3′ and (r) 5′-GGC TAC ATT GGT GTT GAG CTT TT-3′; Sp7 (osterix (OSX) gene) (f) 5′-CCC ACC CTT CCC TCA CTC-3′ and (r) 5′-CCT TGT ACC ACG AGC CAT-3′; Bglap2 (osteocalcin (OC) gene) (f) 5′-CTG ACA AAG CCT TCA TGT-3′ and (r) 5′-GCG CCG GAG TCT GTT CAC-3′; Bmp2 (f) 5′-GGG ACC CGC TGT CTT CTA GT-3′ and (r) 5′-TCA ACT CAA ATT CGC TGA GGA C-3′; and Gapdh (f) 5′-TCA ATG ACA ACT TTG TCA AGC-3′ and (r) 5′-CCA GGG TTT CTT ACT CCT TGG-3′. 31
The primer sequences for the human genes were as follows: CTNNB1 (f) 5′-GTT CGT GCA CAT CAG GAT AC-3′ and (r) 5′-CGA TAG CTA GGA TCA TCC TG-3′; WNT3A (f) 5′-GTC CTG TGA CCC CCA TAG GT-3′ and (r) 5′-GCA CCG GTT AGT AGC CTT CC-3′; ALPL (f) 5′-AAC TTC ACC GGC TTG A-3′ and (r) 5′-TTG CCG CGT GTC TT-3′; RUNX2 (f) 5′-CAG ATG GGA CTG TGG CTG T-3′ and (r) 5′-GTG AAG ACG GTT ATG AAG G-3′; SP7 (f) 5′-ACC TAC CCA TCT GAC TTT GCT-3′ and (r) 5′-CCA CTA TTT CCC ACT GCC TTG-3′; BGLAP (f) 5′-GTG CAG AGT CCA GCA AAG GT-3′ and (r) 5′-CGA TAG GCC TCC TGA AAG C-3′; BMP2 (f) 5′-CGG ACT GCG GTC TCC TAA-3′ and (r) 5′-GGA AGC AGC AAC GCT AGA AG-3′; and GAPDH (f) 5′-CCA TCT TCC AGG AGC GAG ATC-3′ and (r) 5′-GCC TTC TCC ATG GTG GTG AA-3′.
Western blot analysis
For preparing the whole cell lysate, cells were lysed using PRO-PREP™, sonicated briefly and centrifuged at 16,000 g for 10 min, following which the supernatants were used for immunoblot analysis. When indicated, nuclear protein fractions were prepared using the NE-PER Nuclear and Cytoplasmic Extraction Reagents. Protein samples were subjected to SDS-PAGE, and electrophoresed proteins were subsequently transferred onto a PVDF membrane. After blocking the membrane with 5% nonfat dry milk in Tris-buffered saline with 0.1% Tween 20, the membrane was sequentially incubated with the indicated primary antibody overnight and the HRP-conjugated secondary antibody for 2 h. Immune complexes were visualized using the Supex reagent, and luminescence was detected using a MicroChemi Imaging System (DNR; Jerusalem, Israel).
Cytotoxicity test
To evaluate the cytotoxicity of Noni leaf extract, C2C12 and hPDL cells were plated at a density of 2 × 103 cells/well in a 96-well plate, and treated with Noni leaf extract at the indicated doses. After incubation for 48 h, cell viability was determined using the EZ-Cytox Cell Viability Assay Kit (Daeil Lab Service; Seoul, Korea) according to the manufacturer's instructions.
Immunofluorescence staining
To examine the effect of Noni extract on the activation of β-catenin, immunofluorescence staining was performed using a β-catenin antibody. C2C12 cells were seeded at a density of 1 × 104 cells per well in a four-well chamber slide. On the following day, cells were serum starved for 18 h and treated with Noni extract in the presence or absence of DKK1 (100 ng/mL) or LY294002 (10 μM) for 4 h. Untreated cells were used as a negative control. Cells were then fixed with 2% formaldehyde in phosphate-buffered saline (PBS), blocked with 5% normal goat serum, and incubated with β-catenin antibody overnight at 4°C. All specimens were rinsed with 0.1% Triton X-100/PBS to remove unbound antibodies, followed by incubation with a Cy3-conjugated secondary antibody for 60 min at room temperature and thorough washing with 0.1% Triton X-100/PBS. For nuclei staining, the cells were mounted with VECTASHIELD, which contains DAPI in the mounting solution, and observed using a Zeiss LSM800 Confocal Laser Scanning Microscope (Carl Zeiss; Munich, Germany) with a 40 × water objective lens. To quantify nuclear translocation of β-catenin, a minimum of 200 cells were counted from at least five different sites and data were presented as the fraction of cells with intense nuclear β-catenin staining.
TOP-Flash assay
C2C12 cells were plated into 96-well plates at a density of 2 × 104 cells/well. On the following day, the cells were transiently transfected with 0.2 μg of TOP-Flash reporter plasmid. To normalize for transfection efficiency, Renilla luciferase plasmid was used as an internal control. After incubation for 24 h, the cells were treated with Noni extract for 48 h in the presence or absence of DKK1 (100 ng/mL) or LY294002 (10 μM). Luciferase activity was measured using the Dual-Glo Luciferase Assay Kit and relative luciferase activity was calculated after normalization to Renilla luciferase activity.
ALP staining and ALP activity assay
To verify the effect of Noni extract on the osteogenic differentiation of C2C12 and hPDL cells, ALP activity was examined by ALP staining or biochemical enzyme activity assay. ALP staining was performed using the ALP Staining Kit according to the manufacturer's instructions and ALP activity assay was performed as previously described using p-nitrophenyl phosphate as the substrate. 28 The amount of p-nitrophenol was quantified by measuring absorbance at 405 nm on a 96-well plate reader. ALP activity was normalized with respect to the protein amount.
Alizarin red S staining
To examine the effect of Noni leaf extract on matrix mineralization of hPDL cells, the cells were plated in a 24-well plate at the density of 1 × 104 cells/well. Cells were incubated for 28 days in the presence or absence of the extract, and cell culture medium was changed every 2–3 days. At the end of the culture period, matrix mineralization was examined by Alizarin Red S staining. In brief, cells were fixed with 70% ethanol, washed with PBS and stained with 2% Alizarin Red S solution for 10 min at room temperature. After thorough rinsing with distilled water, the calcium deposition was quantified by elution of the stain with 0.5 N HCl containing 5% sodium dodecyl sulfate and measurement of the optical density at 415 nm. 32
Statistical analysis
All of the quantitative data are presented as the mean ± standard error (SEM). Statistical significance was analyzed by Student's t-test or ANOVA with multiple comparison tests using Prism6. A P-value of <.05 was considered statistically significant.
Results
HPLC analysis of major constituents of Noni leaf water extract
The standard materials used for the qualitative analysis of Noni leaf water extract were rutin (M.W. 610.52) and kaempferol-3-O-rutinoside (M.W. 594.52). Rutin and kaempferol-3-O-rutinoside were identified in the extract by comparing each standard retention time obtained during HPLC analysis (Fig. 1). UV spectra and LC-MS/MS analysis of those two peaks further confirmed that those peaks were rutin and kaempferol-3-O-rutinoside (data not shown).

Qualitative analysis of Noni leaf water extract. HPLC chromatogram of standard rutin at 350 nm
Noni leaf enhances osteogenic differentiation of hPDL cells
We first assessed the cytotoxicity of Noni leaf extract with respect to hPDL cells. A cell viability assay was performed after incubation for 48 h in the presence of Noni extract at a concentration of 0.015%–1.0% (w/v%). Noni leaf extract did not present any cytotoxic effect at doses used in the present study (Fig. 2A). Thus, to examine the minimal dose of Noni leaf extract that enhances osteogenic differentiation, we chose the Noni extract concentrations of 0.0038%, 0.0075%, and 0.015% (w/v%) for further study. When osteogenic differentiation of hPDL cells is induced by osteogenic supplements (ascorbic acid, β-glycerophosphate and dexamethasone), osteogenic marker genes are highly induced at 7–14 days and matrix mineralization starts to appear around 14–21 days after induction. 33,34 Therefore, in this study, hPDL cells were incubated with the indicated doses of Noni leaf extract for 14 or 28 days. At the end of the culture period, ALP staining, ALP activity assay, and Alizarin Red S staining were performed. Noni leaf substantially enhanced ALP activity as well as matrix mineralization of hPDL cells (Fig. 2B, C).

Noni leaf promoted osteogenic differentiation of hPDL cells. hPDL cells were treated with the indicated doses of Noni leaf extract, and untreated cells were used as negative control. After 2 days, a cell viability assay was performed
To assure that Noni leaf increases osteogenic differentiation, hPDL cells were cultured with or without Noni leaf (0.0038%) for 7 days. The qRT-PCR results showed that the levels of bone markers (RUNX2, ALP, OSX, and OC) were significantly increased by Noni leaf extract (Fig. 2D). It is well accepted that BMP2 and Wnt/β-catenin signaling pathways are crucial inducers of osteogenic lineage commitment and differentiation. 28,35,36 Hence, to understand the possible mechanisms of Noni leaf extract for osteogenic differentiation, qRT-PCR of BMP2, WNT3A, and β-catenin were also performed. Interestingly, Noni leaf extract also distinctly induced the expression levels of BMP2, WNT3A, and β-catenin (Fig. 2D).
Noni leaf induces osteogenic differentiation of C2C12 cells
C2C12 is a murine myoblast cell line, which can be differentiated to osteoblastic lineage cells under the appropriate culture conditions such as with BMP2 at a dose ≥50 ng/mL. ALP activity and osteogenic marker gene expression are highly induced after 2–6 days depending on the concentration and/or efficacy of BMP2. 37,38 Given that Noni leaf extract enhanced BMP2, Wnt3a, and β-catenin expression in hPDL cells, the next series of the study aimed to determine whether combining Noni leaf extract and a subminimal dose of BMP2 (20 ng/mL) could synergistically enhance lineage commitment and osteogenic differentiation in C2C12 cells.
We first assessed the cytotoxicity of Noni extract in C2C12 cells. As shown in Figure 3A, a subminimal dose of BMP2 treatment with or without the indicated doses of Noni leaf did not significantly affect cell viability.

Noni leaf induced osteogenic differentiation of C2C12 cells. C2C12 cells were treated with various doses of Noni leaf extract (0.030%–1.0%) in the presence of a subminimal dose of BMP2 (20 ng/mL). After 2 days, a cell viability was examined
C2C12 cells were then cultured with 20 ng/mL of BMP2 in the presence or absence of 0.0038% Noni leaf for 4 days. ALP assay results showed that the subminimal dose of BMP2 slightly enhanced ALP staining and activity, which were further enhanced by Noni leaf extract (Fig. 3B). Under the same experimental conditions, qRT-PCR analysis of bone markers showed that subminimal dose of BMP2 significantly upregulated ALP, OC, BMP2, and β-catenin expression, but not Runx2 and OSX expression in comparison with the control, suggesting that a 20 ng/mL dose of BMP2 has only a minimal effect on osteogenic differentiation. Consistent with Noni leaf-treated hPDL cells, all osteoblast markers, BMP2, Wnt3a, and β-catenin, were significantly enhanced on treatment with a combination of BMP2 and Noni leaf extract (Fig. 3C).
Our findings, as illustrated in Figures 2 and 3, suggest that the ability to induce osteogenic differentiation by Noni leaf was significant in both human and murine cells. In addition, BMP2 and Wnt/β-catenin signaling pathways may be involved in the enhancing effect of Noni leaf extract on osteogenic differentiation.
Noni leaf induces osteogenic differentiation through the activation of Wnt/β-catenin signaling
The Wnt/β-catenin pathway has emerged as a promising regulator in enhancing bone and periodontal regeneration. 27,28 Because Noni leaf extract could promote osteogenic differentiation and the expression of Wnt3a and β-catenin, we next examined whether the Wnt/β-catenin signaling pathway plays a crucial role in Noni leaf-induced osteogenic differentiation. C2C12 cells were incubated with BMP2 and Noni in the presence and absence of DKK1 (100 ng/mL), a Wnt inhibitor. As shown in Figures 4A and B, the expression levels of osteoblast marker genes and ALP activity were enhanced by Noni, which were blocked by DKK1, suggesting that activation of the Wnt/β-catenin pathway is involved in the osteogenic differentiation-inducing effect of Noni.
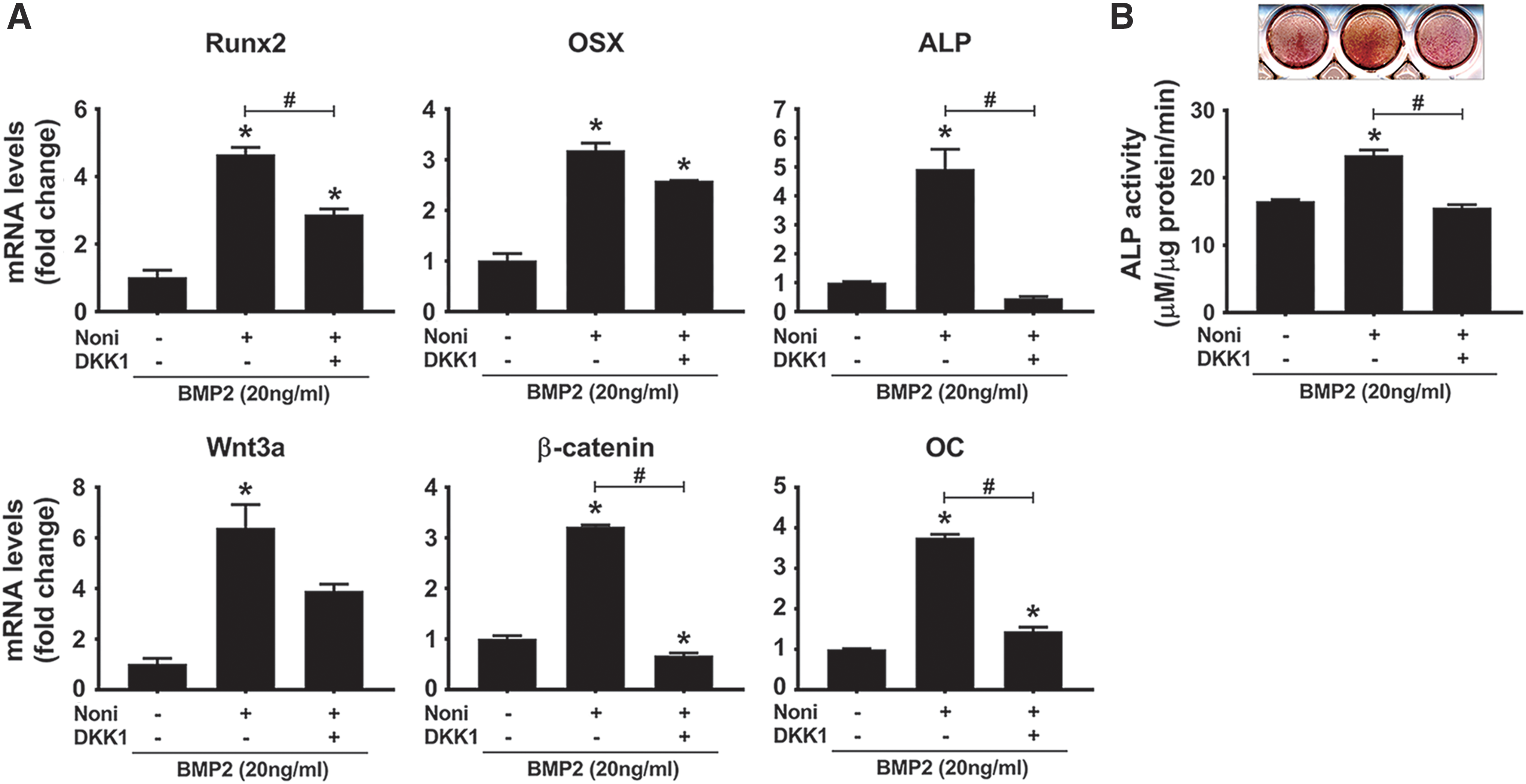
DKK1 attenuated Noni leaf-induced osteogenic differentiation. C2C12 cells were cultured with BMP2 in the presence or absence of 0.0038% of Noni leaf extract. DKK1 (100 ng/mL) was additionally added to the culture medium when indicated. After 4 days, qRT-PCR
We next examined whether Noni leaf extract alone could activate β-catenin and contribute to the increase in osteogenic differentiation. C2C12 cells were incubated in the presence of 0.0038% Noni leaf for 0, 24, and 48 h. The pattern observed in qRT-PCR demonstrated that the increases in mRNA expression levels of Runx2, OSX, BMP2, and Wnt3a were obvious at as early as 24 h, whereas those in mRNA expression of ALP, OC, and β-catenin were obvious at 48 h after incubation with Noni leaf extract alone (Fig. 5A).

Noni leaf induced osteogenic differentiation through β-catenin activation.
To further evaluate whether Noni leaf extract shows an osteogenic induction property by activating the Wnt/β-catenin signaling pathway, the TOP-Flash reporter assay in C2C12 was performed. All doses of Noni leaf extract (0.0038%, 0.0075%, and 0.015%) significantly enhanced luciferase activity (Fig. 5B). However, when DKK1 was added under similar experimental conditions, Noni-induced TOP-Flash activity was attenuated (Fig. 5C).
The nuclear accumulation of β-catenin is a crucial step, which is indicated for the activation of the Wnt/β-catenin signaling pathway. To explore the direct correlation of Noni leaf with Wnt/β-catenin activation, C2C12 cells were serum starved and treated with Noni leaf (0.0038%) in the presence and absence of DKK1. After 4 h, the nuclear localization of β-catenin was analyzed in the nuclear extract using western blot analysis, and this activation was visualized by immunofluorescence. The treatment of the cells with Noni leaf for 4 h upregulated the nuclear β-catenin protein level, whereas DKK1 noticeably blocked β-catenin activation (Fig. 5D). Corresponding results were also observed by immunofluorescence. β-catenin immunostaining was more intense and localized in the nucleus in the presence of Noni leaf extract. Conversely, DKK1 treatment prevented β-catenin activation to similar control cell levels (Fig. 5E). Quantifying the fraction of cells with nuclear localized β-catenin further confirmed that Noni extract significantly induced nuclear accumulation of β-catenin, which was blocked by the addition of DKK1 (Fig. 5F). These findings suggest that Noni leaf extract enhanced osteogenic lineage and differentiation, which are mediated by activation of the Wnt/β-catenin signaling pathway.
PI3K/Akt activation is involved in the activation of the Wnt/β-catenin signaling by Noni
The PI3K/Akt signaling pathway is one of the crucial transducers of responses to various kinds of molecules and growth factors. It is also well known that the Akt pathway plays an important role in bone development. 39 In addition, a recent study has suggested that the Akt pathway could interact with Wnt/β-catenin through phosphorylation and subsequent inactivation of GSK3β, resulting in the nuclear accumulation of β-catenin and thereby promoting osteogenic differentiation. 40 Therefore, we hypothesized that Noni leaf extract induction of the Wnt/β-catenin signaling may involve the PI3K/Akt and GSK3β pathways. To address this hypothesis, serum-starved C2C12 cells were incubated with Noni leaf extract (0.0038%) for 0, 5, 15, 30, and 60 min, and western blot analyses were performed. Figure 6A illustrates that Noni leaf extract induced phosphorylation of Akt and GSK3β and increased the β-catenin protein level.
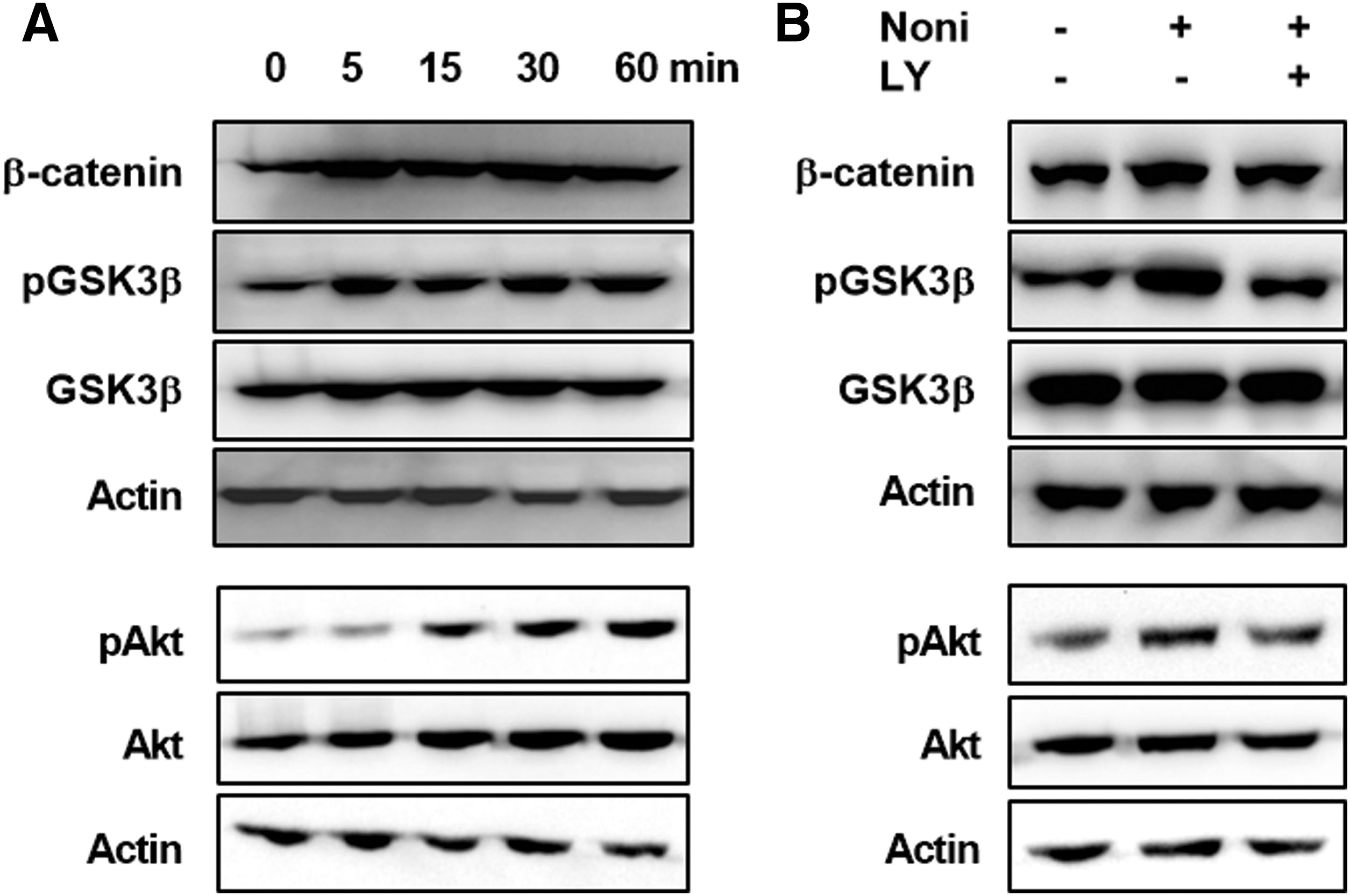
Noni extract enhanced phosphorylation of Akt and GSK3β and accumulation of β-catenin. Serum-starved C2C12 cells were treated with Noni leaf extract.
To assess the involvement of PI3K/Akt activation in Noni-induced Wnt/β-catenin activation, C2C12 cells were incubated with Noni in the presence and absence of LY294002 (10 μM), a PI3K inhibitor. Control cells were not treated. As expected, Noni leaf extract was able to increase phosphorylation of Akt and GSK3β and the protein level of β-catenin, which were blocked by LY294002 treatment (Fig. 6B). These findings suggest that Noni leaf-induced PI3K/Akt activation results in β-catenin activation by inactivating GSK3β.
Noni leaf-induced PI3K/Akt activation contributes to osteogenic differentiation
To investigate the contribution of PI3K/Akt activation in Noni-induced osteogenic differentiation, C2C12 cells were treated with Noni leaf extract (0.0038%) in the presence and absence of LY294002. Untreated cells were used as negative control. Total RNA was isolated and qRT-PCR of osteogenesis genes was performed. As shown in Figure 7A, LY294002 significantly reduced Noni leaf-induced expression levels of osteogenic marker genes, BMP2, Wnt3a, and β-catenin. These results indicate that a decrease in Akt phosphorylation suppresses Noni leaf-induced osteogenic differentiation.
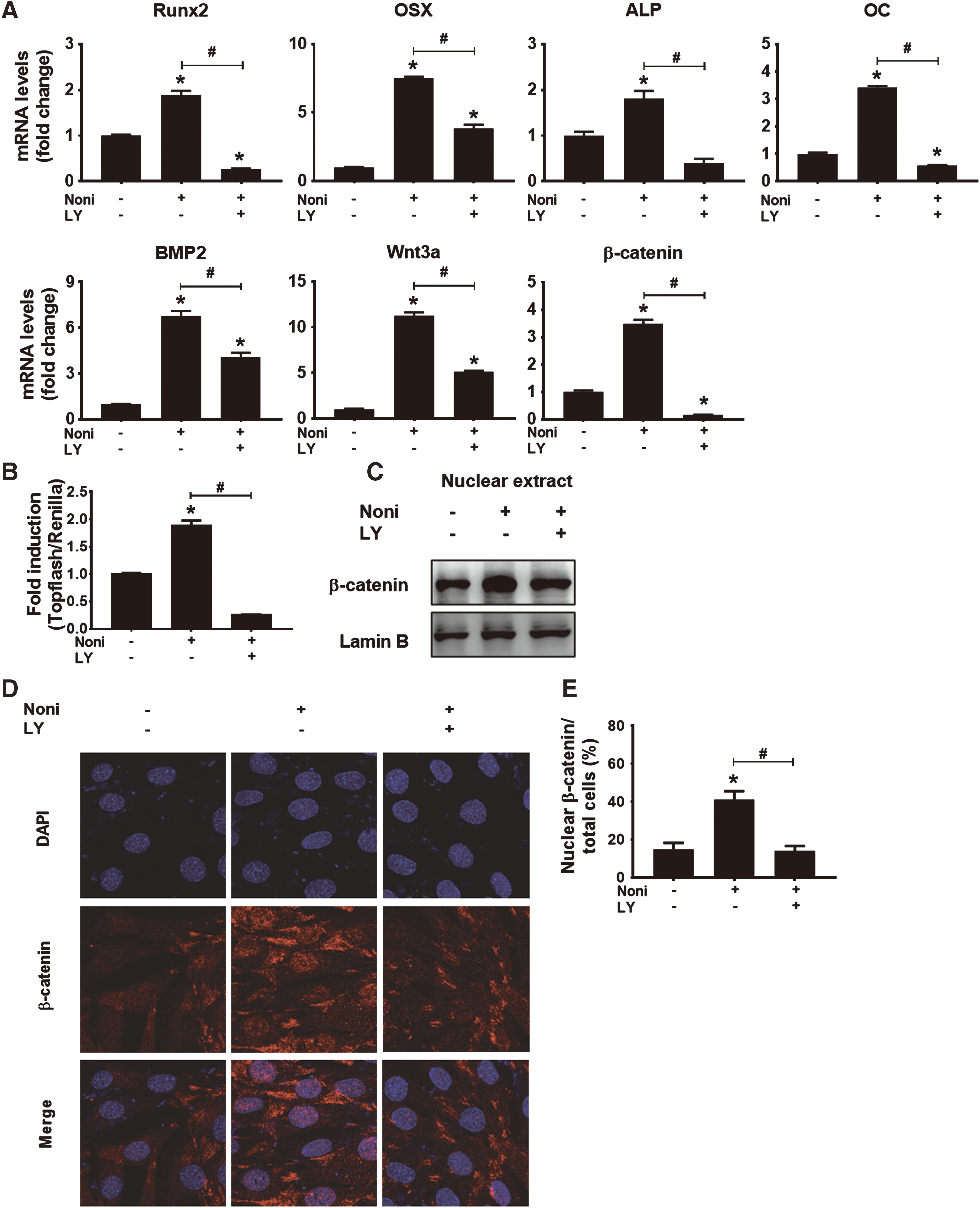
Noni leaf-induced PI3K/Akt activation contributed to osteogenic differentiation.
We next verified whether PI3K/Akt inhibition blocks Noni leaf-induced Wnt/β-catenin signaling pathway. Decreasing PI3K/Akt activation attenuated Noni leaf-induced TOP-Flash activity (Fig. 7B). Moreover, western blot analysis of nuclear extract revealed that prevention of Akt phosphorylation decreased the accumulation of β-catenin in the nucleus (Fig. 7C). Immunostaining of β-catenin further confirmed that Noni-induced accumulation and translocation of β-catenin into the nucleus is dependent on the PI3K/Akt activation (Fig. 7D, E). Taken together, these data suggest that PI3K/Akt activation is necessary for Noni leaf-induced Wnt/β-catenin signaling resulting in osteogenic differentiation.
Discussion
Microorganisms distributed across the entire oral cavity play a crucial role in periodontal destruction, including alveolar bone loss. 1,2 It is well known that regeneration of alveolar bone requires an osteogenic inducer (biological factors), progenitor cells/mesenchymal stem cells, and scaffolds. 6,10,11 The common clinically used osteogenesis-enhancing products (e.g., enamel matrix derivatives) are costly and provide unpredictable results. 9,11 Therefore, natural products, which are cost effective and have lower side effects, may be considered as an alternative medicine for bone regeneration. Furthermore, in a recent systemic review, natural products for osteogenic activity were identified from many studies. Morinda citrifolia, Aloe vera, Fructus cnidii, Zanthoxylum schinifolium, Centella asiatica, and Epimedium species and their related compounds, acemannan, osthole, hesperetin, asiaticoside, and icariin, have all been reported to have potential effects on osteogenic differentiation, cell proliferation, mineral deposition, and gene and protein expression. 41
Noni is an evergreen plant; therefore, the leaf is available during all seasons in tropical countries. 19,21 Noni leaf is not only used as a traditional medicine but also as a food in Thailand. As a result, the use of Noni leaf is very safe and Noni has few adverse effects. Recent scientific research has supported the indigenous uses of Noni leaf, which include promoting wound healing and bone repair. 24 –26 A number of major components such as scopoletin, octanoic acid, potassium, vitamin C, terpenoids, alkaloids, anthraquinones (such as nordamnacanthal, morindone, rubiadin, rubiadin-1-methyl ether, and anthraquinone glycoside), β-sitosterol, carotene, vitamin A, flavone glycosides, linoleic acid, Alizarin, amino acids, acubin, L-asperuloside, caproic acid, caprylic acid, ursolic acid, rutin, kaempferol-3-O-rutinoside, and a putative proxeronine have been identified in the Noni plant. 21 However, the role of Noni leaf extract in the induction of osteogenic differentiation remains unclear.
Osteogenic differentiation and subsequent bone formation requires the expression of many genes, including two master transcription factors (Runx2 and OSX), an early differentiation marker (ALP), and late differentiation marker (OC). 35,42,43 Therefore, the promising inducer for osteogenic differentiation should be able to enhance the expressions of all the genes involved. Our previous study found that 2.5% (w/v%) of Noni leaf extract could promote ALP activity and calcium deposition in hPDL and MC3T3E-1. 26 However, the quantitative detection of mineral composition and associated bone markers (Runx2, OSX, ALP, and OC), detailed molecular mechanisms, and signaling involved remain unknown. Furthermore, to gain more information relevant to safety and clinical application of Noni leaf extract, we aimed to determine the minimal effective dose of Noni leaf extract that can enhance osteogenic differentiation. These results showed that Noni leaf extract at concentrations of 0.0038%, 0.0075%, and 0.015% were sufficient to increase ALP activity and matrix mineralization in hPDL cells. In addition, either Noni leaf alone or its combined use with a subminimal dose of BMP2 can accelerate lineage commitment and osteogenic differentiation in C2C12 cells, suggesting that Noni leaves are promising osteogenic inducers of bone regeneration and repair. Our study involved the use of human and murine cells because the use of nonhuman cells would provide results that could be inconsistent due to species differences in response to Noni leaves. For example, BMP2 could induce in vitro mineralization in murine PDL cells but not human. 35,44 However, our data showed that Noni leaf promoted osteogenic differentiation in both human and murine cells.
Osteogenic differentiation is an important step in bone remodeling and periodontal regeneration, which is regulated by various signaling pathways, including Wnt/β-catenin, 27 parathyroid hormone, 45 bone morphogenetic protein, 31,35 epidermal growth factor, 28,29 and basic fibroblast growth factor. 5 Because Wnt/β-catenin activation mostly contributes to osteogenic and cementogenic differentiation and bone formation, this pathway is of present interest as a crucial target for bone and periodontal regeneration. 27,29,46 Given our results that Noni leaf extract could strikingly potentiate the mRNA expression of BMP2, Wnt3a, and β-catenin and the protein levels of pAkt, pGSK3β, and β-catenin, it is hypothesized that the underlying mechanisms of Noni leaf-induced osteogenic lineage commitment and differentiation may involve the PI3K/Akt and Wnt/β-catenin signaling pathways.
pAkt is known to phosphorylate and inactivate GSK3β, an inhibitor of the Wnt/β-catenin signaling pathway. Therefore, an increase in pAkt activity could have accelerated the Wnt/β-catenin signaling pathway resulting in osteogenic lineage commitment and differentiation. 40,47 Consistent with this report, it was found that pAkt, pGSK3β, and β-catenin were considerably enhanced in Noni-treated cells. In addition, Noni leaf significantly increased transcriptional activity of β-catenin, which was abolished in the presence of LY294002. Furthermore, LY294002 markedly inhibited Noni leaf-induced expression of osteogenesis-related genes, including Wnt3a and Ctnnb1. Collectively, our results indicate that Noni leaf extract enhanced osteogenic differentiation through PI3K/Akt-dependent activation of Wnt/β-catenin signaling.
The chemical profile of Noni leaf showed that its aqueous extract contains two major components: rutin and kaempferol-3-O-rutinoside. It has been shown that rutin alone significantly increased proliferation and osteogenic differentiation of human osteoblast-like MG-63 cells through its enhancements of ALP activity, collagen type I expression, and mineralization. 48 In addition, kaempferol-3-O-rutinoside significantly increased proliferation and osteogenic differentiation of MC3T3-E1 cells, an osteoblast-like cell line from C57BL/6 mouse calvaria, through its enhancements of cell viability, ALP activity, and mineralization. 49 Therefore, Noni leaf extract might exert its action on osteogenic lineage commitment and differentiation through PI3K/Akt-dependent activation of Wnt/β-catenin signaling through rutin and kaempferol-3-O-rutinoside.
In conclusion, Noni leaf extract induced the PI3K/Akt signaling pathway contributing to Wnt/β-catenin activation, which in turn enhanced osteogenic differentiation. Our results indicate that the aqueous extract of the Noni leaf might be a novel alternative medicine for bone and periodontal regeneration of patients with periodontal diseases.
Footnotes
Acknowledgment
This study was supported by an NRF grant through the Oromaxillofacial Dysfunction Research Center for the Elderly (No. 2014 050477) at Seoul National University in Korea.
Author Disclosure Statement
All of the authors state that they have no conflicts of interest.