
Abstract
Herbal medicines were subjected to enzyme reaction by using a commercial glycosidase AMG-300L, and were evaluated for enhancement of their antioxidative activities. The methanolic extract of Gentianae Scabrae Radix (GSR) showed the most dramatic changes after enzyme reaction, as seen in the high-performance liquid chromatography profiles and an increase in the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging effect. Trifloroside (
Introduction
F
Recently, it was confirmed that the crude enzyme extract derived from the fungus Aspergillus kawachii, which is required in the production of Meju (a traditional dried fermented soybean in Korea), has β-glucosidase activity. 2 The quercetin content and the neuroprotective effects of onion were increased by biotransformation of quercetin-4′-glucoside into its corresponding aglycone, quercetin. 2,3 Fermented Korean mistletoe extract showed an enhanced antioxidative activity that correlated with an increase in the amounts of caffeic acid and lyoniresinol. 1 These reports demonstrate that the decomposition of a glycoside into an aglycone by enzyme reaction could be a good strategy to enhance biological activities, such as antioxidation and neuroprotection.
Although cells have defensive systems against reactive oxygen species (ROS), imbalances between ROS production and the capacity of defense mechanisms may contribute to the pathogenesis of various aging-related disorders, including arthritis, atherosclerosis, cancer, and neurodegenerative diseases. 4 High concentration of glutamate causes oxidative stress through inhibition of the glutamate/cystine antiporter, as shown in the mouse-derived hippocampal neuronal cell line (HT22). Subsequently, the cellular antioxidant glutathione (GSH) becomes depleted, and this phenomenon propagates oxidative stress stimuli through increases in Ca2+ influx, intracellular ROS production, and lipid peroxidation. 5 Therefore, identification of antioxidants that can interfere with glutamate toxicity would be helpful for the development of foods or drugs with neuroprotective functions.
Intrigued by the idea that the simple process of enzyme reaction could enhance the antioxidant and neuroprotective activities of herbal medicines, we treated 11 medicinal plants with the commercial exo-1,4-α-
Materials and Methods
Plant materials and sample preparation
Eleven herbal medicines whose antioxidative and neuroprotective activities had never been proven were selected from a group of 60 different plants. Selected plant materials were purchased from a local market in Daegu, Republic of Korea, and identified by Prof. Kyung-Sik Song (College of Pharmacy, Kyungpook National University). The voucher specimens are deposited at the Laboratory of Natural Products Medicine, College of Pharmacy, Kyungpook National University. Plants were refluxed twice with MeOH at 70°C for 3 h. The methanolic extracts were used for enzyme reaction, HPLC analysis, and DPPH radical scavenging activity.
Enzyme reaction
Methanolic extracts (50 mg each) were suspended in 1 mL of distilled water (DW), and then an aliquot of 100 μL AMG-300L (enzyme activity, 300 amyloglucosidase Unit/mL; Novozymes, Bagsvaerd, Denmark), which was produced by Aspergillus niger and used in food industry, was added to the suspension. The mixture was incubated at 30°C. After 24 h, the reaction mixture was extracted with 1 mL EtOAc. The EtOAc-soluble extract was dissolved in dimethyl sulfoxide (DMSO) to a concentration of 100 mg/mL. The solution was filtered through a 0.2 μm polytetrafluoroethylene membrane filter (Whatman, Little Chalfont, United Kingdom), and filtrate (10 μL) was applied to the HPLC system. Boiled AMG-300L was used as a control.
To establish the optimal conditions for enzyme reaction, the methanolic extract of GSR (1 g) was suspended in 20 mL DW, and 2 mL AMG-300L was added. The mixture was incubated at 30°C for 0.5, 1, 3, 6, 12, 24, and 36 h. Then, 1 mL of the reaction mixture was partitioned with 1 mL EtOAc. The EtOAc-soluble extract was dried and the extract was dissolved in DMSO to a concentration of 100 mg/mL. The solution was filtered, and the filtrate (10 μL) was analyzed by HPLC.
HPLC analysis
For HPLC, HPLC-grade acetonitrile (MeCN) and acetic acid (HOAc) were used (Burdick & Jackson, Muskegon, MI, USA). The HPLC system was DIONEX UltiMate 3000 (Thermo Scientific, Dreieich, Germany) equipped with a diode array detector-3000, TCC-3000SD column compartment, WPS-3000SL Analytical autosampler, and LPG-3400SD quaternary pump. The stationary phase was a Kinetex RP18 (4.6 × 250 mm, 5 μm; Phenomenex, Torrance, CA, USA) and the mobile phases consisted of water (solvent A) and MeCN (solvent B), each containing 0.1% HOAc. Gradient elution was carried out as follows. Solvent B was increased from 5% to 40% in 30 min, then 40% to 100% in 5 min, held at 100% for 2 min, and returned to 5% in 3 min. The flow rate was held at 1.0 mL/min. Absorbance of ultraviolet light at 254 nm was used for detection.
DPPH radical scavenging assay
This was performed according to our previous report. 6 The DPPH (Sigma-Aldrich Co., St. Louis, MO, USA) was dissolved in EtOH to prepare a 150 μM solution. DPPH solution (190 μL) was reacted with 10 μL of test sample (which had been dissolved in DMSO with various concentrations) for 30 min at room temperature in the dark. The optical density (OD) was measured at 517 nm, and the DPPH radical scavenging activity was calculated as a percentage relative to the control (10 μL DMSO without test sample), which was considered as 0%.
Bulk reaction of GSR
To isolate a compound that was observed to be increased after enzyme treatment, the reaction was scaled up. The methanolic extract of GSR (1.0 kg) obtained from 4.0 kg of dried GSR was suspended in 1 L DW, mixed with 100 mL AMG-300L, and incubated for 24 h at 30°C. The reaction mixture was sequentially partitioned with n-hexane, CH2Cl2, and EtOAc to obtain the CH2Cl2-soluble extract.
Isolation of compounds
The CH2Cl2-soluble extract (17.2 g) was applied to a silica gel column (Ø 4 × 70 cm, CH2Cl2: MeOH = 100: 1–5: 1) to yield 15 fractions (Fr. 1–15). Fr. 13 (1.5 g) was rechromatographed with silica gel (Ø 2 × 50 cm, CH2Cl2: MeOH = 10: 1–5: 1) and Diaion HP-20 (Ø 1.0 × 7.5 cm, 100% water), sequentially, to isolate the compound designated
Trolox equivalent antioxidant capacity assay
The 2,2′-azino-bis(3-ethylethylbenzothiazoline-6-sulfuric acid (ABTS, Sigma-Aldrich Co.) radical scavenging activity of each isolated compound was evaluated as previously described. 1 To produce the ABTS cation radical, a mixture containing 7 mM ABTS in water and 4.9 mM potassium persulfate was stored at 4°C in the dark. After 12 h, the mixture was diluted with 80% EtOH to get an OD of 0.7 at 734 nm. Next, 190 μL of this mixture was added to 10 μL of compound solution or a solution of Trolox (Sigma-Aldrich Co.) standard at various concentrations. After incubation at room temperature for 30 min in the dark, the reduction of OD at 734 nm was measured relative to the blank. The ABTS radical scavenging activity was calculated according to the calibration curve of Trolox standard solution, and expressed as Trolox equivalent activity (μM).
Cell culture and MTT cell viability assay
The mouse-derived hippocampal neuronal cell line (HT22) was cultured as previous reports. 6,7
Cells were seeded at a density of 3 × 104 cells/mL on a 24-well plate. After incubation for 24 h, test samples were treated to the cells for 2 h before the induction of cell death by incubation with 10 mM glutamate for 24 h. The supernatant was removed and 0.5 mg/mL of 3-(4,5-demethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Amresco, Solon, OH, USA) in DMEM without serum and phenol red was added. After 2 h, the insoluble purple formazan, which was produced by live cells, was dissolved in DMSO, and the OD was measured at 575 nm. The cell viability was calculated relative to the vehicle control. Ten micromolar quercetin was used as a positive control. 3
Measurement of lactate dehydrogenase leakage
After HT22 cells were treated with test samples as well as glutamate as described in Cell culture and MTT cell viability assay section, the supernatant and cell lysate were tested with the CytoTox96 Non-Radioactive Cytotoxicity Assay Kit (Promega; Madison, WI, USA) according to the manufacturer's instructions.
Flow cytometry analysis
After the HT22 cells were treated with 10 mM glutamate for 6 h, the cells were collected by centrifugation and washed once in DMEM without serum and phenol red. Five micromolar Fluo-4 AM (Invitrogen, Eugene, OR, USA), 5 μM of 5-(and-6-) chloromethyl-2′,7′-dichlorohydrofluorescein diacetate and acetyl ester (CM-H2DCFDA; Invitrogen), and 1 μg/mL of 4,4-difluoro-5-(4-phenyl-1,3-butadienyl)-4-bora-3a,4a-diaza-s-indacene-3-undecanoic acid (BODIPY 581/591 C11; Invitrogen) were treated to cells and incubated for 15 min in the dark at 37°C, to evaluate the Ca2+ influx, intracellular ROS production, and lipid peroxidation, respectively. For each of the samples, 10,000 cells were applied to the FACS ARIA III (BD Biosciences, San Jose, CA, USA). Green fluorescence intensity was measured for the investigation of Ca2+ influx and intracellular ROS production, whereas red fluorescence intensity was measured for lipid peroxidation analysis. The ratio of the fluorescence intensity of green (oxidized form of BODIPY 581/591) to that of red (naive form of BODIPY 581/591) was considered as the lipid peroxidation. All data were expressed as fold values relative to the vehicle control. The positive control was 10 μM quercetin. 6
Statistical analysis
All experiments were performed in triplicate, and data are expressed as mean ± standard deviation. The statistical significance was determined by one-way ANOVA by using Prism7 software (GraphPad, La Jolla, CA). P-values of <.05 were considered to be significant.
Results
The effects of enzyme reaction
Changes in HPLC profile
AMG-300L hydrolyzes the terminal α-
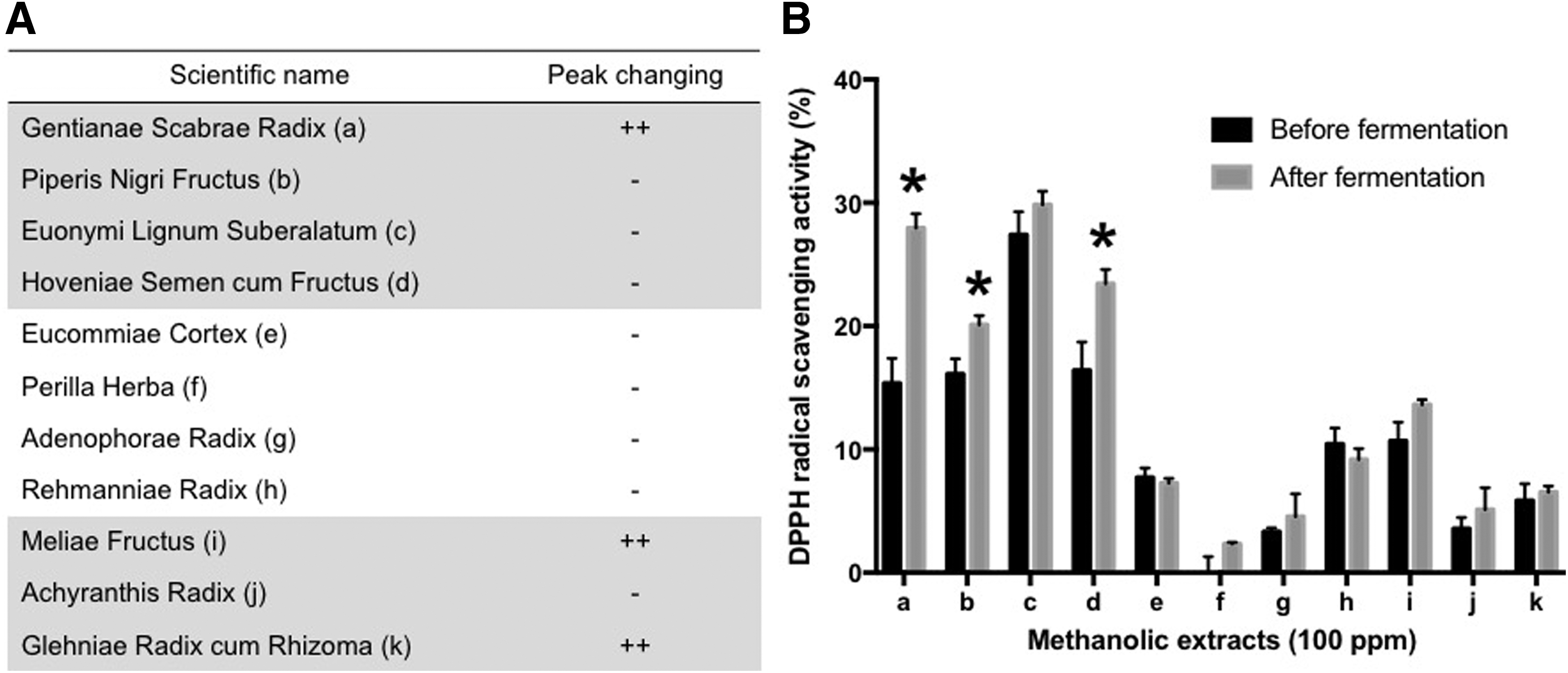
Effects of enzyme treatment on 11 medicinal plants, using the commercial enzyme AMG-300L, as assessed by high-performance liquid chromatography profile
Changes in DPPH radical scavenging activity
As shown in Figure 1B, GSR (a), Piperis Nigri Fructus (b), and Hoveniae Semen cum Fructus (d) exhibited significant changes in DPPH radical scavenging activities after enzyme treatment. Notably, the DPPH radical scavenging activity of GSR was the most dramatic [15.36% ± 2.02% to 27.96% ± 1.16% compared with the control (0.00% ± 1.43%)].
Establishment of optimal treatment time of enzyme
The peak at retention time (Rt) 34.88 min gradually increased as treatment time increased, whereas that at Rt 28.90 min decreased. The peak reached its maximal level after 24 h treatment and remained unchanged until 36 h (Fig. 2A). Therefore, 24 h was chosen as the incubation time for the bulk reaction.
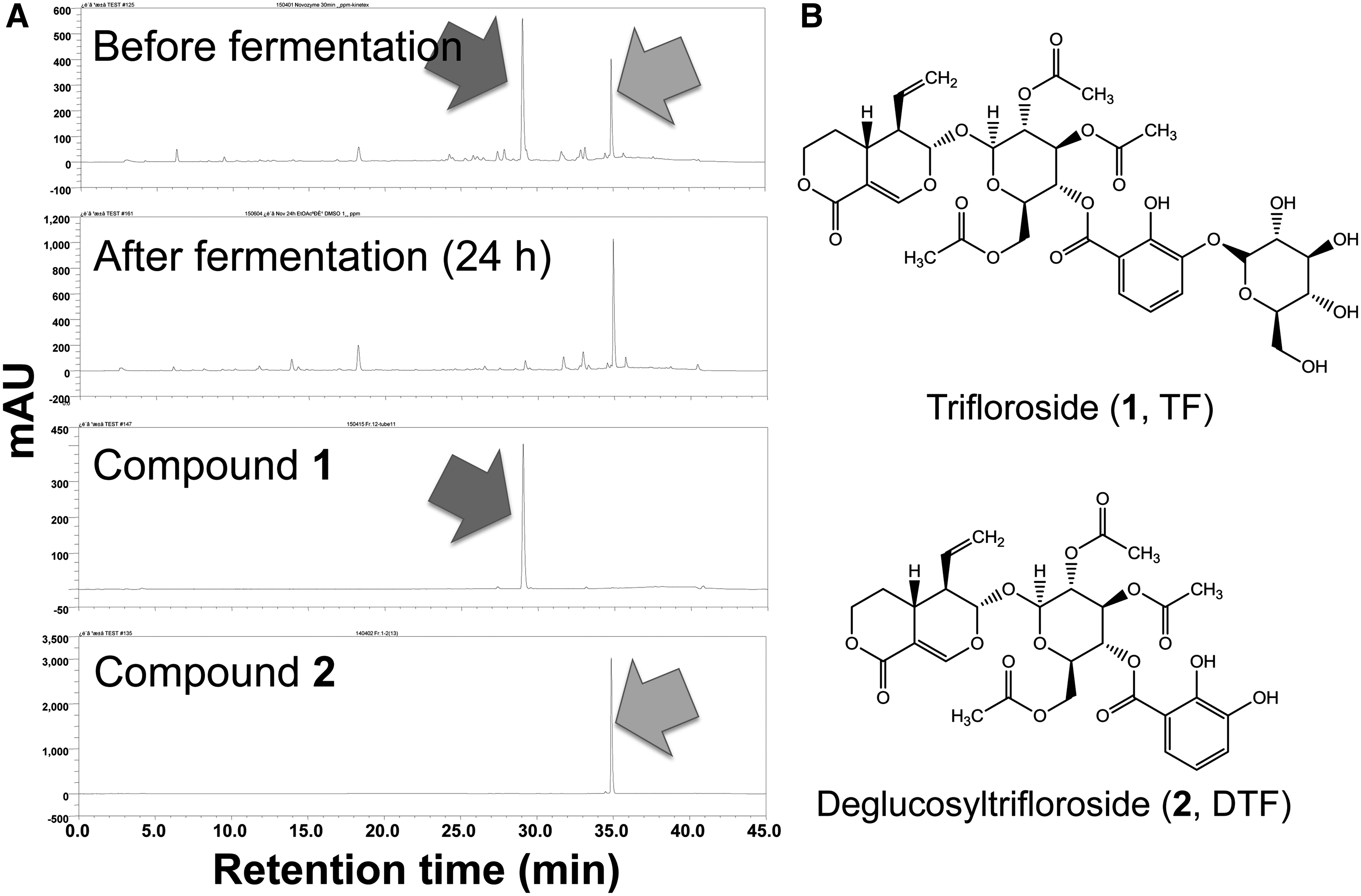
Changes in the contents of trifloroside (
Identification of isolated compounds
To elucidate the chemical structures of
The quantification of DTF in GSR
To evaluate the changes in the content of DTF in GSR, a calibration curve of
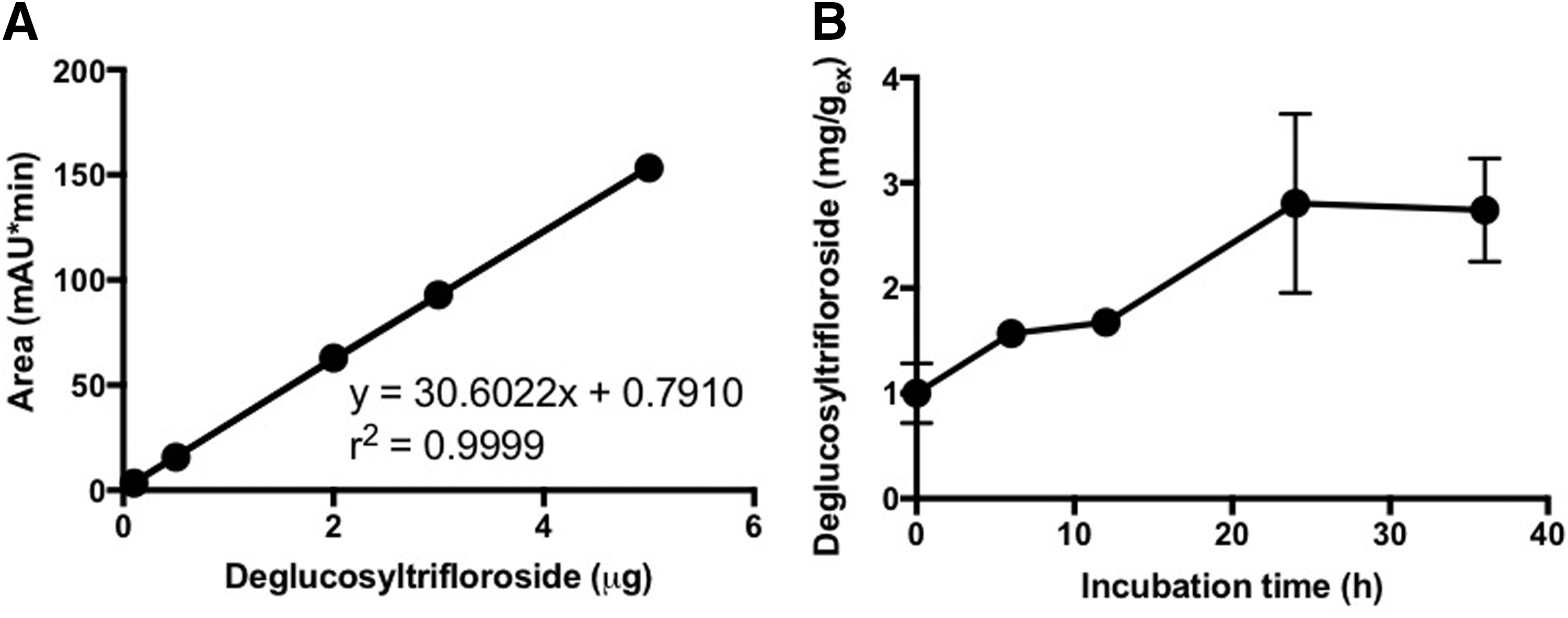
The calibration curve of DTF
Comparison of antioxidative and neuroprotective effects between TF and DTF
As shown in Figure 4A and B, the increased compound DTF displayed better antioxidative activity than the decreased compound TF in the DPPH radical scavenging assay as well as the Trolox equivalent antioxidant capacity (TEAC) assay. At 100 μM, DPPH radical scavenging activities of TF and DTF were 2.87% ± 0.97% and 51.97% ± 1.29%, respectively. In addition, the Trolox equivalent activities of TF and DTF were 43.17 ± 15.30 μM and 103.3 ± 5.03 μM, respectively.
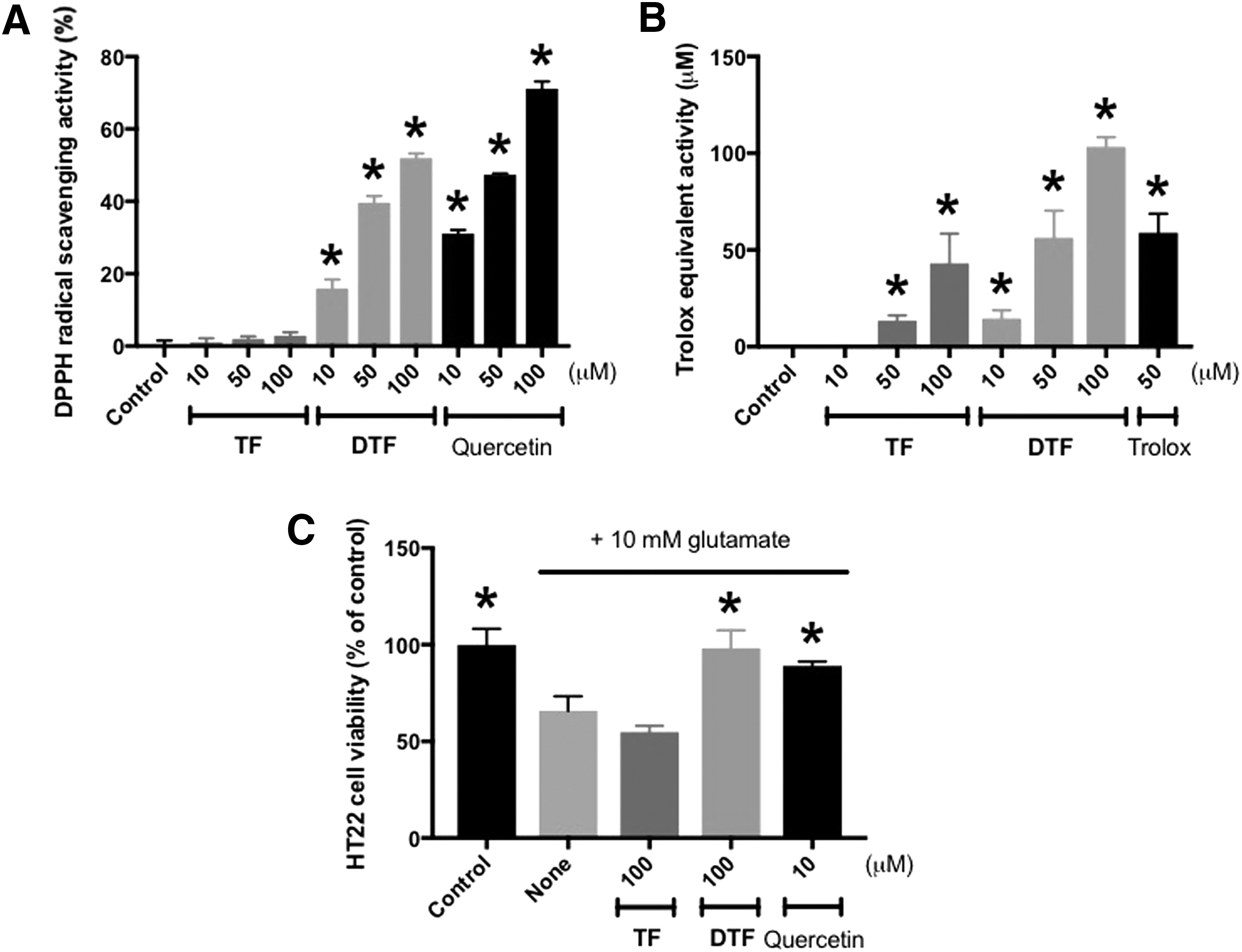
Comparison of antioxidative activities
In the MTT cell viability assay, the cell viability of HT22 was significantly recovered by 100 μM DTF to 98.25% ± 9.24% compared with the vehicle control (100.00% ± 8.21%), whereas that of the 10 mM glutamate-only treated group was 65.85% ± 7.44%. In contrast, the effect of TF was negligible. These data indicated that the increased compound DTF might have contributed to the enhanced DPPH radical scavenging activity of the enzyme-treated GSR as shown in Figure 1B.
The protective effect of DTF against glutamate-induced cytotoxicity
When cytotoxicity was induced by 10 mM glutamate, HT22 cells had the shrunken cell morphology; however, cells treated with a 50 μM solution of DTF or 10 μM quercetin looked healthy (Fig. 5A). The loss of cell viability was also attenuated by DTF in a concentration-dependent manner. Notably, the cell viability of 50 μM DTF-treated cells was 73.50% ± 3.27%, whereas that of the glutamate-only treated cells was 24.05% ± 0.38%, compared with the control (100.00% ± 2.52%; Fig. 5B). In addition, the increased lactate dehydrogenase (LDH) leakage induced by glutamate (4.79 ± 0.53-fold) was significantly reduced by DTF, in a concentration-dependent manner. The LDH leakage was decreased to 1.46 ± 0.56-fold by pretreatment with 50 μM DTF (control, 1.00 ± 0.08-fold; Fig. 5C).

The protective effect of DTF against glutamate-induced HT22 cell death.
The inhibitory effect of DTF on glutamate-induced Ca2+ influx, lipid peroxidation, and intracellular ROS production
The flow cytometry data indicated that 10 mM glutamate mediated the elevation of Ca2+ influx by 2.76 ± 0.10-fold over the control (0.92 ± 0.08-fold). However, DTF significantly suppressed Ca2+ influx in a concentration-dependent manner (0.91 ± 0.01-folds at 50 μM; Fig. 6A). The lipid peroxidation was also attenuated by pretreatment with DTF (2.33 ± 0.02-fold at 50 μM) compared with the control (1.00 ± 0.02-fold), whereas that of the glutamate-only treated group was 7.28 ± 0.10-fold (Fig. 6B). Moreover, 50 μM DTF decreased the glutamate-induced intracellular ROS production (1.73 ± 0.03-fold) to 1.05 ± 0.02-fold (control, 1.00 ± 0.04-fold; Fig. 6C).

The inhibitory effect of DTF on glutamate-induced Ca2+ influx
Discussion
GSR, an herbal medicine, containing the roots or rhizomes from Gentiana lutea, Gentiana scabra, Gentiana manshurica, and Gentiana triflora, has been used for stimulation of appetite and the treatment of stomach ailments, 11 hepatitis, and inflammation. 12 GSR mainly contains secoiridoid glycosides such as gentiopicroside, sweroside, swertiamarin, 12 and gentiascabraside A, 13 and triterpenoids including scabanol, 14 chiratenol, lupeol, and α-amyrin. 15
In this study, we treated a commercial exo-1,4-α-
The active compound DTF has been isolated as a constituent of GSR; however, there are few studies about its biological activities. Recently, Li et al. investigated the anti-inflammatory effect that DTF had on lipopolysaccharide-stimulated bone marrow-derived dendritic cells from wild type C57BL/6 mice through suppression of the production of proinflammatory cytokines such as interleukin (IL)-12 p40 and IL-6. 18
This is the first report about the enzyme reaction of GSR and its enhanced antioxidative and HT22 cell protective effects. Further studies such as in vivo and clinical studies on the enzyme-treated GSR might lead to the functional foods capable of preventing oxidative stresses, especially neurodegeneration.
Footnotes
Acknowledgment
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ011344)” Rural Development Administration, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.