
Abstract
Obesity is generally associated with low-grade chronic inflammation that involves the recruitment of macrophages and other inflammation factors to the adipocytes of obese individuals. Tumor necrosis factor-alpha (TNF-α), a cytokine associated with systemic inflammation, is elevated in conditions of obesity. TNF-α is an important factor that plays an important role in skeletal muscle wasting. Apoptosis of myonuclei contributes to the loss of muscle mass and therefore plays an important role in skeletal muscle atrophy. In mouse models that were fed a high fat diet (HFD), a lipolysis-stimulating peptide-VHVV (purified from hydrolysate resulting from flavourzyme treatment of soy protein) was found to reduce HFD-related apoptotic effects in mice skeletal muscle and potentially control atrophy. HFD fed mice had heavier body weight than those fed with normal chow, and VHVV administration restricted lipid accumulation in muscle tissues of mice fed with HFD but increased nutrient uptake. Moreover, specific concentrations of VHVV regulated TNF-α expression that was elevated by HFD, suppressed apoptosis-related proteins and regulated the proteins of lipid metabolism.
Introduction
T
Natural products such as acidic polysaccharide and hydroxypropyl methyl cellulose can suppress HFD-induced disease risk. 5,6 Several reports have indicated that hydrolyzed protein extracts including soy protein hydrolysates play an important role in reducing body fat and blood triglycerides. 7 –12 Flavourzyme is a fungal peptide complex containing both endo-peptidase and exo-peptidase activities and it has been used to produce protein hydrolysates with low bitterness. 13 Enzymatic hydrolysis of soy protein has promoted several bioactive functions, such as antioxidative and cholesterol-lowering activities. 14 A lipolysis-stimulating peptide, VHVV, derived from flavourzyme-soy protein isolate hydrolysate was found to have the potential to act as a functional anti-obesity diet ingredient. 9,14 Administration of VHVV has previously shown to reduce plasma triglycerides level and the low-density lipoprotein-cholesterol levels in mouse models. VHVV induced reduction in the plasma lipid levels also correlates with amelioration of hepatic apoptosis and fibrosis effects in a dose-dependent manner. VHVV also displays anti-inflammatory effect and inhibits HFD-induced liver damage. 9 This study is aimed at investigating the effects of VHVV on lipid accumulation and amelioration of apoptosis in skeletal muscles. Male C57BL/6 mice that were fed HFD for 8 weeks (HFD group) were found to develop symptoms obesity and apoptosis. However, HFD fed mice treated with different doses of VHVV for the final 6 weeks showed a significant reduction in symptoms, although VHVV did not cause any considerable change in HFD-induced elevation in the total body weight. Further the proteins such as Bad and cytochrome c involved in the mitochondrial apoptosis pathways that were elevated in mice fed with HFD were effectively suppressed by VHVV treatment in all the doses tested and therefore VHVV effectively promotes mitochondrial stability. Our results indicate that VHVV attenuates HFD-induced adverse effects in the skeletal muscles and, therefore, can be considered as a potential therapeutic agent in the amelioration of obesity-related muscle wasting.
Materials and Methods
Animal experiments
This study was conducted following the IACUC-100-12 protocol and approved by the IACUC Ethics Committee. The mice (6-week-old male C57BL/6) were allowed for a week to adapt to the pathogen-free animal room environment and the normal laboratory chow. During the adaptation period, the animals were individually housed in a room maintained at 24°C ± 2°C and 55% ± 10% humidity with a 12 h light cycle. The control animals were fed with a standard laboratory diet (PMI Nutrition International, Brentwood, MO, USA) and reverse osmosis-treated water was provided ad libitum throughout the experiment. During the first 4 weeks animals (Fig. 1) in the other groups were fed with HFD containing 60% of energy as fat to induce obesity. The obese mice were randomly divided into five groups (n = 8): sedentary control, HFD-induced obese sedentary control, HFD with low dose (5 mg/kg/day) VHVV treatment (LT), HFD with moderate (15 mg/kg/day) VHVV treatment (MT), and HFD with high (25 mg/kg/day) VHVV treatment (HT). The VHVV was administered through intra-peritoneal (IP) injection for the next 6 weeks of HFD induction. Equal volumes of vehicle (0.9% saline) were administered to the control and HFD group rats through IP injection. The body weight and the muscle weight and characteristics of soleus muscles were carefully harvested and analyzed as reported previously. 15
Representative cartoon of animal experiment schedule.
Hemotoxyline and eosin staining
The muscle tissue from the test mice were excised and then soaked in formalin before being dehydrated by passing consecutively through 100%, 95%, and 75% alcohol. The tissue samples were then embedded in paraffin wax. The embedded tissue blocks were cut into 0.2 μm-thick sections and de-paraffinized by soaking in xylene. The slices were stained by hematoxylin and eosin (H&E) and rinsed with water. Photomicrographs were obtained using Zeiss Axiophot microscopes (Carl Zeiss Microscopy, Thornwood, NY, USA).
Tissue protein extraction
Tissue extracts were obtained by homogenizing the muscle sections in 100 mg/mL lysis buffer (20 mM Tris, 2 mM ethylenediaminetetraacetic acid, 10% glycerol, 50 mM 2-mercaptoethanol, protease inhibitor, and phosphatase inhibitor, pH 7.4). The homogenates were placed on ice and then centrifuged at 12,000 g for 40 min. The supernatants were collected and stored at −80°C for further experiments.
Western blot
Protein concentration of the extract was determined by the Lowry method. Protein samples were separated in a 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) with a 75 V constant power supply. Proteins were then transferred to polyvinylidene fluoride (GE Healthcare Life Sciences, Pittsburgh, PA, USA) membranes using 50 V current for 3 h. The membranes were incubated in 3% bovine serum albumin in Tris-buffered saline and then the primary antibodies were added onto the membranes for conjugation with specific proteins. All of the following primary antibodies were procured from Santa Cruz Biotechnology (Santa Cruz, CA, USA), unless stated otherwise; TNF-α (sc-1350), IL-6 (sc-1266), Bad (sc-8044), Cytochrome c (sc-13560), cleaved caspase 3 (#9664; Cell Signalling), caspase 9 (sc-8355), Bcl-2 (sc-73820), foxo3a (#2497; Cell Signalling), foxo1 (#2880; Cell Signalling), PPARα (sc-9000), and tubulin (sc-5286). Horseradish peroxidase-labeled secondary antibodies were used for detection and pictures were finally taken with Fujifilm LAS-3000 (GE Healthcare Life Sciences).
Results
Effects of VHVV on HFD-induced obesity
HFD intake increased body weight of test mice by 5.5% and treatment with VHVV did not reduce the body weight but instead increased the body weight significantly. The low dose VHVV treatment groups showed a 12.59% increase in body weight whereas the moderate and high dose treatment showed 8.15% and 11.52% increase (Fig. 2A). Moreover, the total dried body fat was found in significantly reduced levels in the mice treated with VHVV (Fig. 2B). Therefore, treatment with VHVV can be inferred to provide antiobesity effect and it is also correlated with TG/HDL ratios that were significantly increased in HFD group and decreased in the VHVV treatment groups (Fig. 2C). The changes in the skeletal muscle architecture, such as increased interstitial space (black arrows) induced by HFD, was also reduced after VHVV treatment (Fig. 2D). Further, the soleus muscle weight reduced significantly in the HFD fed mice group as seen from the soleus muscle to body weight ratio (Fig. 2E), However, treatment with VHVV effectively restored the HFD-induced changes in the muscle weight. Therefore, these results show that VHVV treatment reduced muscle atrophic effects in HFD fed mice.

Effect of VHVV on total body weight and skeletal muscles.
Effect of VHVV on the levels of inflammatory cytokines
In the HFD fed group, the levels of TNF-α increased in the soleus muscle with a corresponding significant increase in the levels of interleukin-6 (IL-6). The levels of proteins remained low in the VHVV administered mouse groups (Fig. 3).
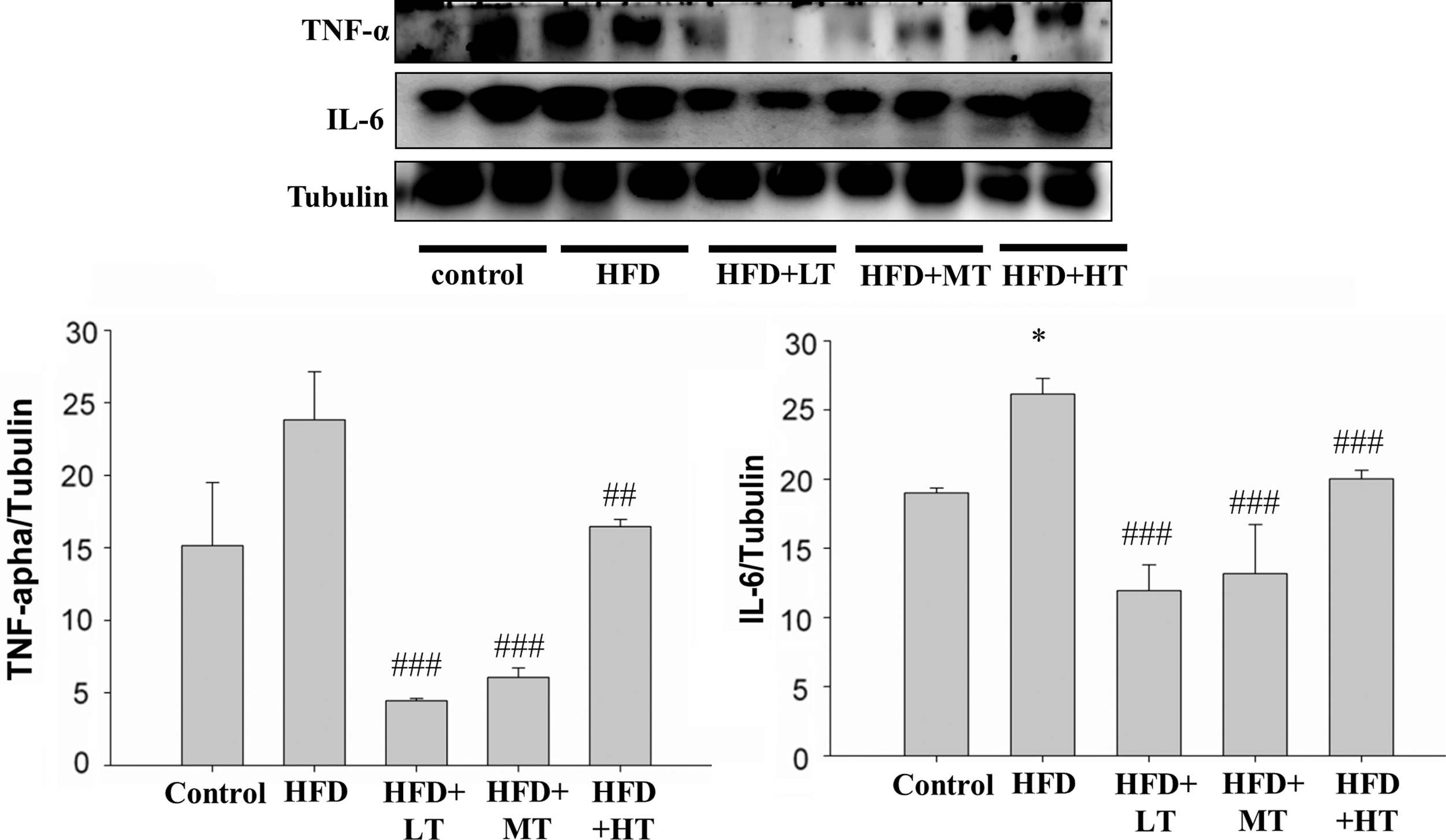
Effect of VHVV on inflammatory cytokines. Western blot analysis on the levels of IL-6 and TNF-α shows the difference between their respective expression in mice fed with regular diet (control), mice fed with HFD, mice fed with HFD with low dose VHVV (HFD+LT) treatment, mice fed with HFD with moderate dose VHVV (HFD+MT) treatment and mice fed with HFD with high dose VHVV (HFD+HT) treatment after 8 weeks of respective feeding. The results represent mean ± SD of three independent experiments. (*P < .05 indicates significant difference with respect to the control group; ## P < .01 and ### P < .001 indicate significant difference with respect to HFD challenged group). IL-6, interleukin-6; TNF-α, tumor necrosis factor alpha.
VHVV administration regulates apoptosis and survival-related proteins
Analysis of protein expression in soleus muscle by western blotting showed that HFD feeding in mice significantly suppressed the levels of the apoptosis inhibitor B cell lymphoma-2 (Bcl-2) and elevated the levels of proapoptotic proteins such as activated caspase 9, caspase 3, and cytochrome c in a significant manner. However, in the mice groups that were administered with low, moderate or high levels of VHVV, the expression of cytochrome c, caspase 9, and caspase 3 was found to be suppressed significantly. VHVV administration also significantly enhanced the expression of apoptosis inhibitor protein Bcl-2 in a dose-dependent manner (Fig. 4). The apoptotic proteins of the extrinsic pathway, such as caspase 8 and caspase 3, were also found to be elevated by HFD, and the increased levels were found to be suppressed in the mice groups treated with VHVV.
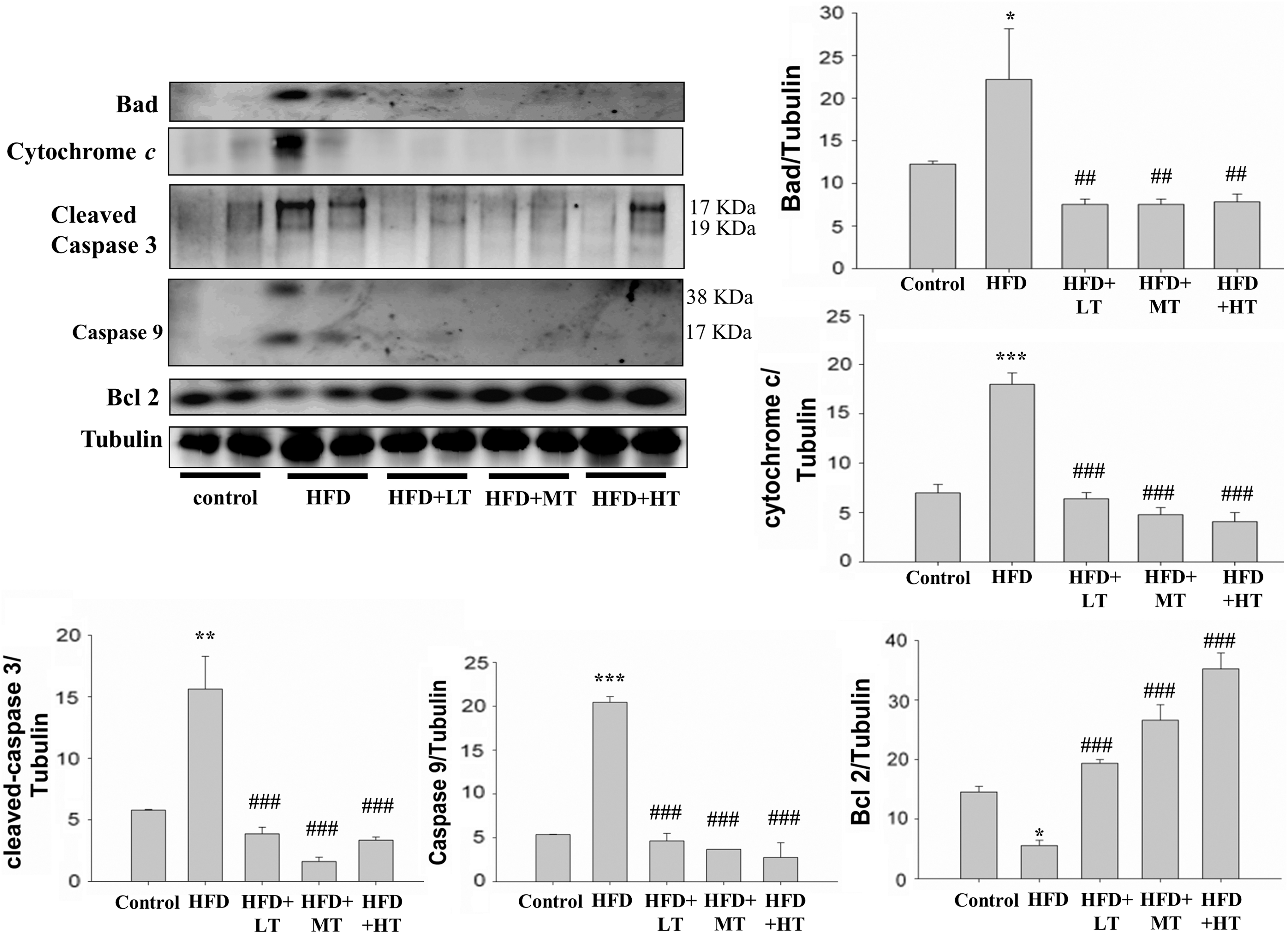
Effect of VHVV on proapoptotic proteins. The levels of proteins related to cell apoptosis, as determined by western blot analysis, when compared with the respective expression levels in the mice fed with regular diet (control) show a suppression in the expression levels of apoptosis proteins such as Bcl-2, Bad, cytochrome c, cleaved caspase 9, and caspase 3 in mice fed with HFD, HFD with moderate dose VHVV (HFD+MT), and HFD with high dose VHVV (HFD+HT) after 8 weeks of respective feeding. The results represent mean ± SD of three independent experiments. (*P < .05, **P < 0.01 and ***P < 0.001 indicate significant difference with respect to the control group; ## P < .01 and ### P < .001 indicate significant difference with respect to HFD challenged group). Bcl-2, B cell lymphoma-2.
Effect of VHVV on lipid metabolism
To determine the effect of VHVV on lipid metabolism, the levels of the regulatory nuclear receptor protein-peroxisome proliferator-activated receptor (PPAR) and phosphorylated-Forkhead box protein (Foxo) 1 and 3a in soleus muscle were examined. PPARα was reduced significantly in mice fed with HFD, whereas the level was comparatively high in the VHVV-treated groups. Meanwhile, the levels of peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), a co-activator of PPARγ, were lower in the HFD fed groups and remained low in the VHVV administered group as in controls (Fig. 5). Foxo3a expression that was suppressed in the mice fed with HFD was elevated in the mice treated with VHVV.
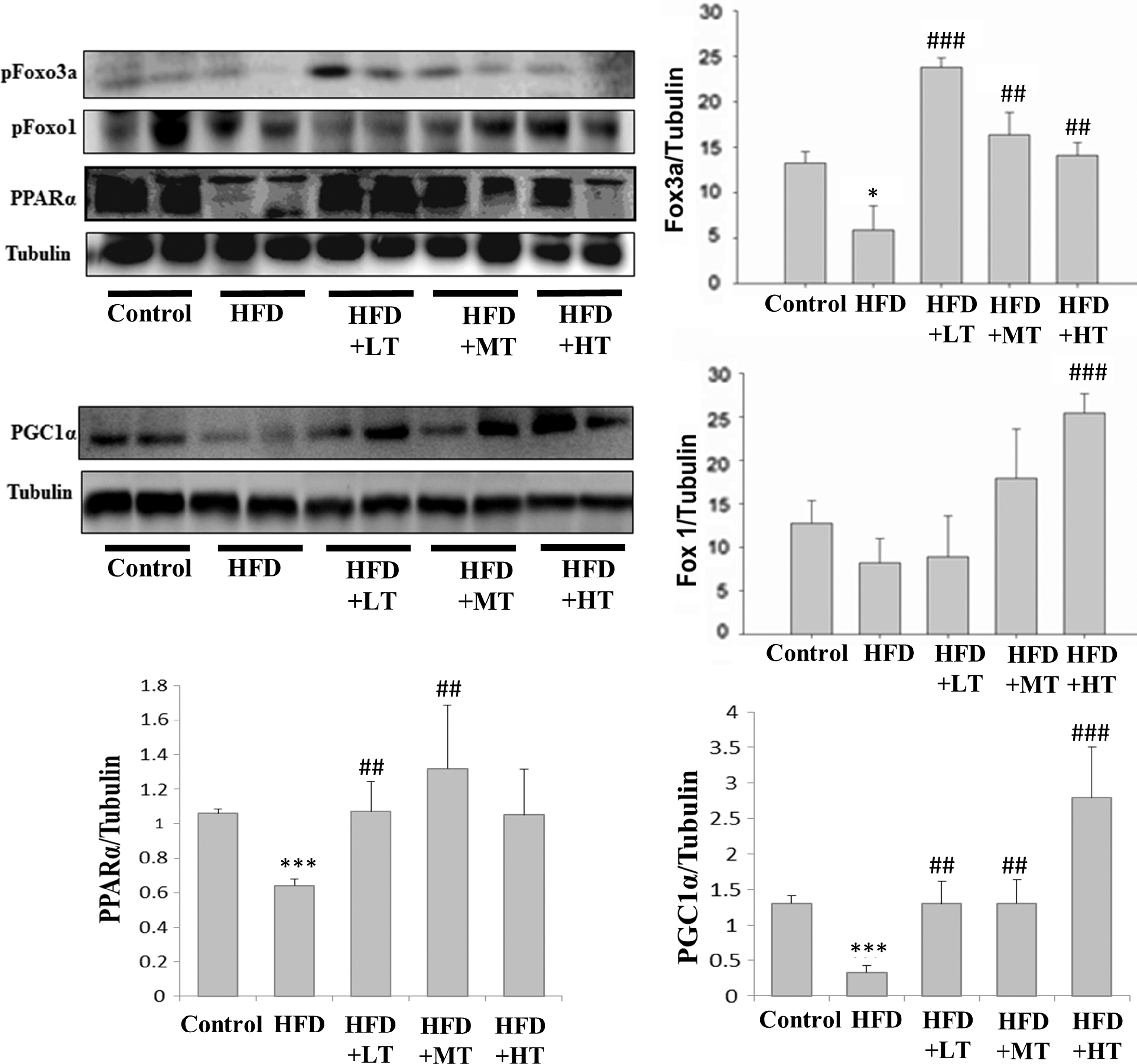
The levels of lipid metabolism related nuclear membrane proteins, as determined by western blot analysis, when compared with the respective expression levels in the mice fed with regular diet (control) show modulations in the expression levels of proteins such as PPARα, FOXO1, and FOXO3a in the mice fed with HFD, mice fed with HFD with low dose VHVV (HFD+LT) treatment, mice fed with HFD with moderate dose VHVV (HFD+MT) treatment, and mice fed with HFD with high dose VHVV (HFD+HT) treatment after 8 weeks of respective feeding. The results represent mean ± SD of three independent experiments. (*P < .05, and ***P < .001 indicates significant difference with respect to the control group; ## P < .01 and ### P < .001 indicate significant difference with respect to HFD challenged group). PPARα, peroxisome proliferator-activated receptor alpha.
Discussion
Obesity is generally defined by increased lipid storage in visceral and subcutaneous adipose tissue, but it is also characterized by ectopic lipid deposition in nonadipose tissues. 16 Unregulated fatty acid levels and intracellular accumulation of lipids generate detrimental changes in cellular function and morphology and that leads to lipotoxic conditions resulting in cell death. 17 Skeletal muscle accounts for 40% of our body mass and plays a major role in fatty acid uptake and oxidation. The excess accumulation of fatty acid, triacylglycerol, and fatty acid metabolites, such as ceramide, diacylglycerol, and long chain acyl CoA, causes changes in muscle function, size, and morphology and may lead to apoptosis. 16 –18
Muscle tissue such as the soleus is composed of high amount of slow fibers and is often susceptible to inactivity. 19 The soleus muscle in the lower hind limb contains higher proportions of slow twitch fibers (type I) and almost no fast-twitch fibers (type II). 20,21 The mitochondrial enzymes are highly active in the soleus showing a high oxidative capacity in the tissue. The mouse soleus muscle also has greater oxidative capacity as evidence by having greater activity of several mitochondrial enzymes. 22 Therefore, in correlation with previous findings our results show that HFD induced the susceptibility to higher protein degradation and loss of muscle mass in type I muscle fibers. 23
The HFD fed mice used in our experiments displayed symptoms of obesity as observed from the increase in their body weight and adipocyte size. The development of obesity in the HFD fed mice was also correlated with changes in the soleus muscle weight. Protein expression analysis on the skeletal muscle proteins shows an increase in inflammatory cytokines in the mice fed with HFD. The elevated plasma TNF-α levels observed in our obese mouse model has also been reported by several other researchers. TNF-α knockout mice, when fed a HFD, do not become obese, but instead increase their caloric intake. 24 Further, TNF receptors are known to cause obesity-associated inflammation in skeletal muscles. 25 The TNF-α signaling cascade has also been implicated as one of the reasons behind the loss of muscle mass. 3 Our results show that HFD feeding significantly increased the levels of TNF-α and IL-6 expression when compared with the mice that were fed standard chow. Supplementation of antioxidant drugs such as resveratrol have been shown to reverse TNF-α-associated muscle atrophy effects by elevating the Foxo1 expression. 26 Three isoforms of Foxo transcription factors: Foxo1, Foxo3a, and Foxo4 are present in skeletal muscles 27 and the modulation in the Akt/FoxO/ubiquitin involved proteolysis pathway is one of the major events in TNF-α-induced muscle atrophy. Phophosphorylation of Foxo proteins by Akt restricts their nuclear translocation and subsequent activation of their target gene expression. 28,29 Foxo1 and Foxo3 expressions trigger atrophy-associated ubiquitin ligases atrogin-1 and MuRF-1 that are responsible for reduction in muscle mass. 30,31 Therefore, decreased phosphorylated Foxo protein levels reflect elevated nuclear translocation and muscle atrophic effects. 32,33 Foxo protein can also act as a cofactor of nuclear receptor activity and regulate various cellular functions, such as the cell cycle and apoptosis. 34 The levels of Foxo1 and Foxo3a that were suppressed in HFD fed mice were found to be enhanced in mice treated with VHVV indicating a role in cell survival.
Excessive apoptosis plays critical role in the development of skeletal muscle atrophy. 35 Unrestricted apoptotic events may also result from suppression of autophagy. 36,37 Consistent with previous reports, our results show excessive apoptosis with respect to elevation in the levels of proapoptotic proteins and reduction in the antiapoptotic proteins; however, it can be significantly ameliorated when treated with low, moderate, and high levels of VHVV.
Our data show a significant increase in apoptosis in the skeletal muscle of the HFD fed obese mice. The process of apoptosis is essential for maintaining the equilibrium of highly proliferative tissue, such as those consisting of epithelial cells. The role of apoptosis in matured tissue, such as skeletal muscle, is less understood. Apoptosis of myocytes may contribute to the skeletal muscle wasting, 38 but the mechanisms underlying this process are not yet clear. In our experiments, the HFD fed group showed an increase in cleaved caspase 3, which is an indicator for the events of apoptosis in the skeletal muscle. 39 Other markers of apoptosis, such as caspase 9 and cytochrome c, were also enhanced in the HFD fed mice.
In our experiments, 6 weeks of treatment with VHVV suppressed TNF-α expression and thereby reduced the chances of obesity. However, the increase in the bodyweight in the VHVV-treated mice reflects increased energy uptake and lipid metabolism. Therefore, VHVV treatment indeed increases the muscle mass of HFD fed mice. The increase in muscle mass was correlated with modulations of the levels of the Bcl-2 family of proteins including the antiapoptotic Bcl-2 and proapoptotic Bad. VHVV treatment increased Bcl-2 expression in a dose-dependent manner and suppressed Bad expression, suggesting a cytoprotective effect conferred by VHVV on skeletal muscles. Decrease in the levels of other molecular markers of apoptosis such as cleaved caspase 3 cleaved caspase 9 and cytochrome c in all the doses tested also reflects the efficient antiapoptotic effect of VHVV.
PPARα, which belongs to the nuclear hormone receptor protein family, plays an important role in glucose and lipid metabolism by regulating transcription. 40,41 PPARα is expressed in skeletal muscle, and it reduces fatty acid content and lipotoxicity by increasing lipid oxidation in the respective tissue. 42 Nonesterified fatty acids are taken up by tissues in conditions such as caloric restriction, and they subsequently get bound to and activate PPARα. 43
While modulation of apoptotic proteins also show a deteriorating mitochondrial integrity in the HFD fed mouse muscles, PGC-1α which, is an inducer of mitochondrial biogenesis, was found to be reduced. The effect of HFD and VHVV on PGC-1α was also correlated with the activation of PPARδ whose enhanced expression results in elevated mitochondrial mass and function. However, treatment with VHVV reverted the levels of PGC-1α in a dose-dependent manner. PGC-1α is highly expressed in the soleus muscles but its expression is generally reduced in conditions such as atrophy. 44 Expression of PGC-1α is also correlated with suppression of muscle protein degradation. 45 Our results also show that the levels of PPARα that were suppressed in the HFD fed mice were comparatively high in the mice treated with VHVV, indicating a regulatory effect of VHVV on lipid metabolism.
In this study, the effect of VHVV to attenuate HFD-related apoptotic and fibrosis effects was determined in the skeletal muscles of HFD fed mouse. HFD induced effects, such as changes in mouse body weight, muscle architecture, lipid levels, and modulations in apoptosis related events were ameliorated by VHVV administration. Thereby, moderate and high concentration of VHVV administration could potentially rescue mice from obesity-related muscle loss.
Footnotes
Acknowledgments
This research was financially supported by a grant from Tunghai University for Global Research and Education on Environment and Society (GREEnS 004-3) and the Ministry of Science and Technology of Taiwan (MOST 104-2410-H-029-033-MY2).
Author Disclosure Statement
No competing financial interests exist.