
Abstract
Traditional herbal remedies stimulate and modulate the immune system, and it is thought that their glycoproteins and polysaccharides are responsible for this activity. We prepared crude water, protein, and polysaccharide extracts from Atractylodes macrocephala Koidz, Helianthus annuus L., Scutellaria barbata D. Don, and Hedyotis diffusa Willd, respectively, and compared their immune-stimulating activities in vitro and in vivo. All protein and polysaccharide samples of the plants led to greater lymphocyte proliferation and TNF-α and IL-6 production in cultured splenocytes than did the crude water extracts at the same concentrations tested. In addition, the protein and polysaccharide samples did not contain lectin- or lipopolysaccharide-like molecules, so glycoproteins were deduced to be responsible for the lymphocyte stimulation. Oral administration with each of the samples enhanced the hen egg-white lysozyme (HEL)-specific humoral immune and lymphocyte proliferative responses in HEL low-responder C57BL/6 mice. Splenocytes from the mice fed the samples showed significantly greater increases in the level of IFN-γ, but not IL-4, after stimulation with HEL compared with that from the untreated control. However, higher increases in HEL-specific IgG1, IgG2b, and IgG3 rather than IgG2a were found in the mice fed the samples. These results indicate that the sample-mediated enhancement of anti-HEL-specific humoral immune responses was due to the stimulation of B lymphocytes rather than a selective priming of helper T cell populations. Collectively, we suggest that glycoproteins and/or polysaccharides of traditional herbal remedies enhance cellular and humoral immune response induction and thus could be useful for patients who need enhanced immune function.
Introduction
T
It is thought that the beneficial effects of traditional medicines are derived from a synergistic interaction among active components present in the medicines, rather than a direct action of individual components. 4,5 This indicates that traditional medicines act as modulators rather than direct regulators of a specific signaling pathway. However, there has been increasing interest in identifying the active ingredients responsible for the biological activities of traditional medicines and in determining their therapeutic efficacies. 6,7
Among the active substances present in traditional medicines, glycoproteins and polysaccharides are considered important constituents for stimulation of the immune system. 8 –10 We previously found that Bo-yang-hwan-o-tang, a traditional Korean medicine, induces hen egg-white lysozyme (HEL)-specific immune responses in HEL low-responder mice, and this induction was closely associated with the immune-stimulating activity of glycoproteins. 11 We also demonstrated that Soamsan, another traditional Korean medicine consisting of a total of 11 components, enhanced anti-HEL antibody responses in mice through combined activity of glycoproteins and endotoxin-like compounds present in the medicine. 12
When lymphocyte proliferation and cytokine production activities of the crude water and protein extracts prepared from each component used in Soamsan were determined using mouse splenocytes, all protein samples showed greater activities than did the crude water extracts at the same concentration. 13 Collectively, these findings along with those of other reports 8 –13 indicate that glycoproteins and polysaccharides are active macromolecules in immune stimulation and modulation by traditional Oriental medicines.
In this study, we sought to confirm the ability of the glycoproteins and/or polysaccharides in several traditional medicines to stimulate the immune system. To this end, we selected Atractylodes macrocephala Koidz (AMK), Helianthus annuus L. (HAL), Scutellaria barbata D. Don (SBD), and Hedyotis diffusa Willd (HDW) among various traditional medicines and prepared crude water, protein, and polysaccharide extracts from these herbal plants. This was due to our previous findings that protein samples obtained from AMK, HAL, and SBD showed greater activities on in vitro lymphocyte proliferation and cytokine production compared to those from other components consisting of the Soamsan. 13 These herbal plants also exerted immune-stimulatory, antimicrobial, antioxidant, and antitumor activities. 14 –16
Specifically, polysaccharide extract from AMK improved disordered intestinal microflora 17 and activated macrophages by stimulating nuclear factor-κB-dependent mechanisms. 18 HDW, a herbal plant that has been widely used in traditional medicine prescriptions, is also known to stimulate macrophage activation and lymphocyte proliferation, 19 as well as to protect mice from endotoxin-induced renal inflammation. 20 We next evaluated the effects of the extractions on lymphocyte proliferation and cytokine production and determined the physiochemical properties of the protein and polysaccharide samples. We also explored whether oral administration of the samples modulates an antigen-specific immune response using HEL as a model antigen and HEL low-responder C57BL/6 mice as an animal model.
Materials and Methods
Chemicals, laboratory equipment, and mice
All reagents required for cytokine production assays were obtained from Pharmingen, Inc. (San Diego, CA, USA). Fetal bovine serum (FBS) was purchased from HyClone (Logan, UT, USA), and [methyl- 3 H] thymidine deoxyribose (TdR) was purchased from Amersham Pharmacia Biotech, Inc. (Piscataway, NJ, USA). Unless otherwise specified, all chemicals were purchased from Sigma Chemical Co. (St. Louis, MO, USA), and all laboratory consumables were purchased from Falcon Labware (Becton-Dickinson, Franklin Lakes, NJ, USA). Inbred male C57BL/6 mice (5-weeks-old) were provided by Damul Science Co. (Yusung, South Korea). Mice were housed in automatically controlled conditions with a 12-h light/12-h dark cycle at 22°C ± 1°C and 45–55% relative humidity. All mice had free access to standard rodent pellet food and water. The animal care and use protocol implemented in this study was approved by the Chonbuk National University Committee on Ethics in the Care and Use of Laboratory Animals (CBU 2010-0007). All mice were acclimatized to the new laboratory environment for1 week before experimental use.
Sample preparations
We prepared crude water, protein, and polysaccharide extract samples from AMK, HAL, SBD, and HDW according to the methods described previously. 13 Briefly, 100 g of each dried component was cut into small pieces, mixed with 1000 mL of distilled water in a screw-capped flask, and then shaken at 4°C for 3 days. The resulting water extracts were filtered through paper (Whatman No. 3) and centrifuged at 5000 g for 10 min. The supernatants were collected and lyophilized to yield dried powders of the crude water extracts, in which the extraction rates of AMK, HAL, SBD, and HDW were around 10%, 6%, 4%, and 8%, respectively, of the initial amount. We repeated this extraction thrice and combined the powders into one tube per sample type. To obtain protein extract samples, the crude water extract (2 g per sample) was resuspended in 40 mL of 70% ammonium sulfate and centrifuged at 10,000 g at 4°C for 20 min to collect the protein precipitates. The precipitates were dialyzed against phosphate-buffered saline (PBS) and lyophilized to yield ∼140, 160, 80, and 120 mg of protein extract samples specific to AMK, HAL, SBD, and HDW, respectively. We repeated this preparation until ∼300 mg of protein was collected per component. To prepare polysaccharide samples, the dried components (100 g per component) were cut into small pieces and boiled in 1000 mL of ethanol (96% purity) for 1 h, followed by filtration using filter paper (Whatman No. 3). The filtrates were extracted twice with 100 mL of heated distilled water (∼80°C), and the collected extracts were mixed with absolute ethanol (1:4, v/v) to allow precipitation of polysaccharides. After centrifugation, the precipitates were collected in absolute ethanol, washed with acetone, and then lyophilized to yield the dried polysaccharide samples. Finally, all samples were stored at −20°C and freshly dissolved before use.
Characterization of protein and polysaccharide samples
All procedures for characterization of the samples were described elsewhere. 13 In brief, protein and polysaccharide samples were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) using a Mini-PROTEAN II cell (Bio-Rad, Hercules, CA, USA) with 10–15% gel. After electrophoresis, gels were stained either with Coomassie brilliant blue (CBB) to detect proteins or with Schiff reagent to detect glycoproteins. To determine whether the immune stimulating activity of protein samples was due to glycoproteins, the protein samples were treated with pronase E to degrade the protein or with NaIO4 to degrade the carbohydrate residues. In brief, each of the protein extracts (5 mg/sample) was incubated in 4 mL of 0.1 M Tris-HCl buffer (pH 8.0) containing 0.6 mg pronase E and 50 mM CaCl2 for 72 h at 30°C, and then the mixtures were heated at 100°C for 10 min to inactivate the enzyme. The same amount of each protein sample was also incubated with 100 μL of 0.1 M NaIO4 at 25°C for 4 h before the addition of 250 μL of 20% ethylene glycol. The protein samples treated with pronase E or NaIO4 were dialyzed against PBS before use. We next explored if the protein and polysaccharide samples contained a lectin compound such as concanavalin A (Con A). To this end, these samples (10 mg/sample) were dissolved in 1 mL distilled water in a tube, and the tubes were soaked in a boiling water bath for 20 min. Alternatively, the protein and polysaccharide samples (1 mg/sample) were dissolved in 1 mL of culture medium containing 1000 units of polymyxin B (PMB) and incubated at 37°C for 3 h to determine if the samples contained lipopolysaccharide (LPS)-like compounds. Each of the samples pretreated with pronase E, NaIO4, PMB, or heating was added to splenocyte cultures at a concentration of 10 μg/mL. The levels of lymphocyte proliferation and cytokine production by the samples were compared to those of the untreated samples through TdR incorporation assay and enzyme-linked immunosorbent assay (ELISA) methods. In these experiments, heat-treated Con A or PMB-treated LPS (5 μg/mL/sample) was added to splenocyte cultures and used as a comparative control.
In vitro assays on lymphocyte proliferation and cytokine production
A single-cell population of splenocytes was prepared from naive C57BL/6 mice and resuspended in RPMI-1640 medium supplemented with antibiotics and 10% FBS. The suspension of splenocytes (4 × 106 cells/mL) was divided onto 24- (500 μL/well) or 96-well flat-bottomed plates (200 μL/well). After 24 h of incubation, culture media were replaced with fresh RPMI medium supplemented with 0.5% FBS and/or 25 μL PBS containing each of the samples at various concentrations (0–50 μg/mL). To assess the effects of the samples on lymphocyte proliferation, 0.5 μCi/mL of [methyl- 3 H] TdR was added to each well of the 96-well plates for the last 16 h of a 48-h culture period. The cells were harvested using a cell harvester (Inotech, Inc., Switzerland), and the incorporated tritium contents were determined using a liquid scintillation counter (Packard Instrument Co., Downers Grove, IL, USA). In addition, supernatants of the splenocyte cultures were collected from 24-well plates after 48 h of incubation, and the levels of cytokines representing the stimulation of Th1-type (IFN-γ), Th2-type (IL-4 and IL-6) T cells, and macrophages (and/or B cells) (TNF-α) were determined by ELISA. The amounts of cytokines produced were calculated from standard curves generated with recombinant cytokines of known concentrations.
In vivo sample treatment, immunization, and collection of splenocytes and serum
We examined whether an antigen-specific immune response is modulated by oral administration of the samples. In this experiment, we selected the samples derived from AMK and HDW based on their extraction rates and in vitro lymphocyte stimulating activities. Briefly, male C56BL/6 mice were randomly divided into 10 groups (n = 10/group), including two control groups (PBS treatment alone). The experimental groups received oral administration of 300 μL PBS containing crude water (200 mg/kg body weight), protein (10 mg/kg body weight), polysaccharide (10 mg/kg body weight), or a combination of protein (5 mg/kg body weight) and polysaccharide extract samples (5 mg/kg body weight) derived from the AMK or HDW every other day for a total of three times. The dosage of the crude water samples was selected based on previous findings, 12,13 where the concentration was extrapolated from the amount administered in Soamsan 12 prescription for humans. We also selected the concentration of AMK or HDW protein and polysaccharide samples according to the extraction rates (from 6% to 7%) from crude water extracts and the activity on splenocyte proliferation in vitro. One day after the last feeding, all groups were immunized subcutaneously with 50 μg of HEL emulsified in complete Freund's adjuvant. To evaluate HEL-specific lymphocyte proliferation and cytokine production levels, half of the immunized mice (n = 5/group) were orally ingested in additional samples at the same dose every other day for a total of five times, and splenocytes were collected from the mice. In contrast, the other half of the immunized mice (n = 5/group) were fed the same dose of the samples every third day for a total of three times and received boost immunization by subcutaneous injection with 50 μg of HEL emulsified in incomplete Freund's adjuvant. Sera were collected from the mice 4 days after boost immunization, and the level of HEL-specific humoral immune responses was determined. The control group received the same volume of PBS before and after immunization with HEL throughout the experimental period.
Evaluation of the level of HEL-specific antibody responses
HEL-specific immunoglobulin (Ig) levels in mice sera were determined by ELISA. Briefly, each well of the ELISA plate was coated with 1 μg of HEL for 2 h at 37°C and blocked with PBS containing 10% FBS and 0.05% Tween-20 for 3 h. After washing, 50 μL of each serum sample was serially diluted in PBS supplemented with 10% FBS and then added to each well, followed by incubation for 2 h at 37°C. The total amount of the HEL-specific antibodies was determined using goat anti-mouse Ig as a secondary antibody. In addition, rat antibodies specific to mouse Ig subclasses—γ1, γ2a, γ2b, and γ3—were used to determine the amount of HEL-specific antibodies of IgG1, IgG2a, IgG2b, and IgG3 subtypes, respectively. The absorbance specific to the substrate added was measured at 405 nm using a SpectraCount™ ELISA reader (Packard Instrument Co.).
Determination of the levels of HEL-specific lymphocyte response and cytokine production
Single-cell suspensions (2 × 106 cells/mL) of splenocytes were prepared from the control and the experimental mice groups fed AMK or HDW samples as described above and divided into 96-well flat bottom plates (100 μL/well). After 24 h of incubation, splenocytes were stimulated with various concentrations (1–30 μM) of HEL and labeled with 0.5 μCi/mL of [methyl- 3 H] TdR for the last 16 h of a 72-h culture period. The levels of TdR incorporated by the cells were determined as described above. To evaluate HEL-specific cytokine production, splenocyte suspensions (2 × 106 cells/mL) prepared from the mice were spread onto 24-well flat bottom plates (500 μL/well) and stimulated with various concentrations (1–30 μM) of HEL. After 48 h of incubation, culture supernatants were collected, and the amounts of cytokines produced by HEL-stimulated lymphocytes were determined by ELISA using the IFN-γ- and IL-4-specific OptEIA™ Kit (PharMingen, Inc.).
Statistical analyses
All results are expressed as the mean ± standard error (SE) of five different samples. One-way analysis of variance was used to determine the significance of differences among three or more groups using SPSS version 12.0. Student's t-test was used only when the significance of differences between two sets of data was determined using the SPSS program. A value of P < .05 was considered statistically significant.
Results
Protein and polysaccharide extracts are the active components responsible for lymphocyte proliferation and cytokine production
Treatment with various concentrations (1–50 μg/mL) of crude water, protein, or polysaccharide extracts prepared from AMK (Fig. 1A), HAL (Fig. 1B), SBD (Fig. 1C), or HDW (Fig. 1D) increased TdR incorporation by splenocytes in a dose-dependent manner. The addition of protein or polysaccharide extracts induced greater splenocyte proliferation compared to that of crude water extracts at the same concentrations. However, the effects of the protein and polysaccharide extracts on splenocyte proliferation differed according to the plant. Polysaccharide extracts from AMK and HAL more strongly enhanced TdR incorporation by the cells than the protein samples from the plants at the same doses (Fig. 1A, B). For example, the polysaccharide extract of AMK increased the TdR uptake approximately fourfold higher compared with its protein extract at the same concentration. In contrast, there was no significant difference in lymphocyte proliferating activity between the protein and polysaccharide extracts derived from SBD and HDW (Fig. 1C, D). A Trypan blue exclusion assay showed that the crude water, the protein extract, and the polysaccharide extract did not have any significant cytotoxic effects on cultured splenocytes at the concentrations tested (data not shown).
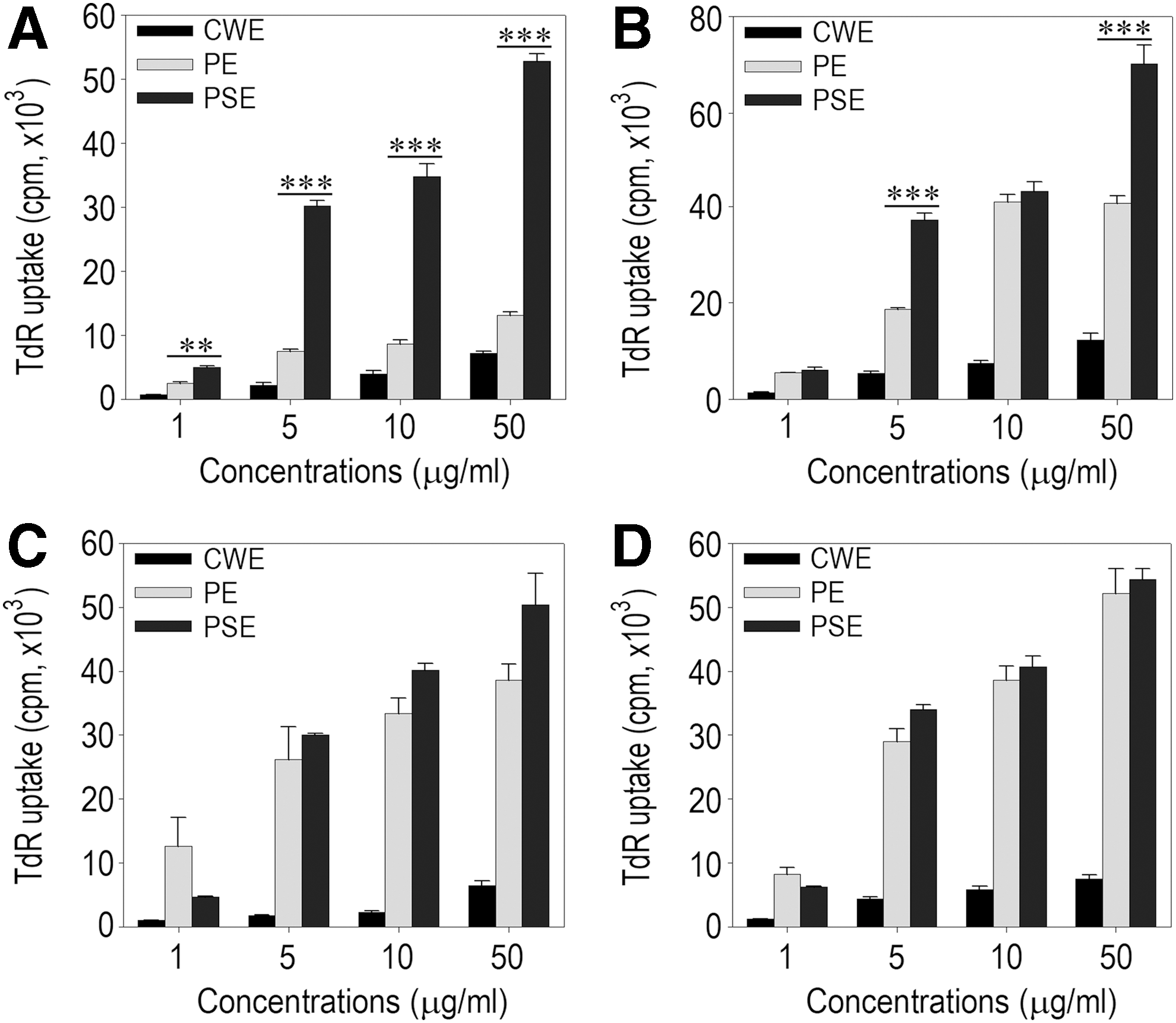
Effects of the sample preparations from traditional medicines on lymphocyte proliferation. Mouse splenocytes were incubated with the indicated concentrations of the CWE, PE, or PSE prepared from AMK
We next examined the stimulating effect of the samples on cytokine production by exposing splenocytes to various concentrations (1–50 μg/mL) of crude water, protein, or polysaccharide extracts prepared from AMK (Fig. 2A), HAL (Fig. 2B), SBD (Fig. 2C), or HDW (Fig. 2D). Results from cytokine ELISA revealed that the concentrations of TNF-α and IL-6 in cultured splenocytes increased in most of the cases where the cells were treated with more than 1 μg/mL of sample. AMK protein and polysaccharide samples greatly stimulated the production of IFN-γ in the splenocyte cultures, and this production was higher compared to that induced by other plant samples at the same concentrations. However, none of the plant samples enhanced IL-4 production in cultured splenocytes, even at the highest dose (50 μg/mL). When splenocytes were treated with SBD or HDW samples, greater TNF-α production was observed in the cultures exposed to protein rather than polysaccharide extract. Both the protein and polysaccharide extracts from AMK, HAL, and SBD more highly increased IL-6 levels in the cultures than did the crude water extracts from these plants at the same concentrations. When HDW samples were added to the cultures, protein extract stimulated the production of IL-6 significantly higher than the crude water or polysaccharide extract. In addition, treatment with more than 1 μg/mL of protein extract from the medicinal plants enhanced the production of TNF-α and IL-6 in cultured splenocytes up to the levels similar to those of the cells stimulated with 5 μg/mL of Con A or LPS (Fig. 2E).

Effects of the sample preparations of traditional medicines on cytokine production by mouse splenocytes. Splenocytes were exposed to various concentrations (1–50 μg/mL) of the CWE,PE, or PSE obtained from AMK
Protein and polysaccharide samples did not contain lectin compounds and were not contaminated with LPS-like mitogens
We characterized the physiochemical properties of protein and polysaccharide extracts by separating the samples by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) before staining with CBB or Schiff reagent (Fig. 3). AMK protein extract showed two major groups of bands around 27 kDa and smaller than 15 kDa together with a broad spectrum of molecular weights by CBB staining (Fig. 3, left-upper panel) and mainly of ∼150 and 27 kDa by Schiff reagent staining (Fig. 3, right-upper panel). The protein extract samples of HAL and SBD revealed one major band of ∼13 kDa, while the HDW protein sample showed several bands of ∼32, 24, 19, and 14 kDa by CBB staining. All protein and polysaccharide samples from HAL, SBD, and HDW revealed a broad spectrum of molecular weights by CBB and Schiff reagent staining. When polysaccharide samples of the plants were separated by SDS-PAGE, the results from CBB or Schiff reagent staining showed similar patterns with those of protein samples, with the only exception that the HAL and SBD samples did not show any apparent bands. These results indicate that the protein and polysaccharide samples from the medicinal plants contained both glycoproteins and polysaccharides with various molecular weights.

SDS-PAGE analysis of protein and polysaccharide extracts prepared from the traditional medicines. Each protein (upper panels) and polysaccharide (lower panels) extract of the medicines was loaded at 800 μg per lane, electrophoresed in a 12% SDS-PAGE, and then stained with CBB (left panels) or Schiff reagent (right panels). The figure shows a representative result from triplicate experiments. CBB, Coomassie brilliant blue; M, molecular weight marker; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis.
To further characterize the active constituents responsible for the splenocyte-stimulating activity, we treated the protein samples with pronase E, NaIO4, PMB, or heating and assessed their ability to stimulate lymphocyte proliferation (Table 1). Treatment of the protein extracts with either pronase E or NaIO4 significantly decreased the potential for the samples to enhance TdR incorporation in the splenocytes. Similarly, treatment with pronase E or NaIO4 led the protein samples to exhibit significantly attenuated production of TNF-α and IL-6 (data not shown). Treatment with PMB, an inhibitor of an LPS-like mitogen-mediated response, or heating did not reduce the activity of the samples to stimulate splenocyte proliferation. In contrast, the stimulating activities of LPS and Con A on splenocyte proliferation were significantly inhibited by treatment with PMB and heating, respectively, where the activity of Con A almost completely disappeared with heating. When the polysaccharide samples of the plants were treated with PMB or heating, there was no significant reduction in their stimulating activity on splenocyte proliferation (data not shown).
Splenocytes were incubated with 10 μg/mL of each protein sample that was pretreated with pronase E, NaIO4, PMB, or heating, and the proliferation rate was determined by TdR incorporation assay. The mitogens (5 μg/mL of LPS or Con A) treated with PMB or heating were added into the cells as positive controls. The results are expressed as the mean TdR uptake in cpm ± standard error of triplicate cultures. * P < .05, ** P < .01, and *** P < .001 compared to the control values.
AMK, Atractylodes macrocephala Koidz; CWE, crude water extract; HAL, Helianthus annuus L; HDW, Hedyotis diffusa Willd; LPS, lipopolysaccharide; PMB, polymyxin B.
Oral administration with AMK and HDW samples enhanced HEL-specific humoral immune responses
We investigated the effect of the samples prepared from AMK and HDW on HEL-specific antibody immune responses. To this purpose, we collected serum from C57BL/6 mice administered the plant samples 4 days after booster immunization and determined the level of HEL-specific IgG by ELISA. Figure 4 shows the HEL-specific IgG values obtained with equal volumes of the sera diluted 128-fold. The level of HEL-specific IgG in the serum collected from the control group was significantly lower than that in the serum from the groups fed each of the AMK samples (left panel). The groups administered crude water extract or combination with protein and polysaccharide extracts showed greater enhancement of the IgG value compared to the groups fed either protein or polysaccharide extract alone. Similarly, all groups fed an HDW sample or combination with its protein and polysaccharide extracts revealed significantly higher HEL-specific IgG in the serum compared to that of the control group (right panel). These results suggest that oral administration of AMK or HDW samples enhanced the induction of antigen-specific antibody immune responses, and this enhancement was closely associated with the activities of glycoproteins and polysaccharides to stimulate immune response induction.
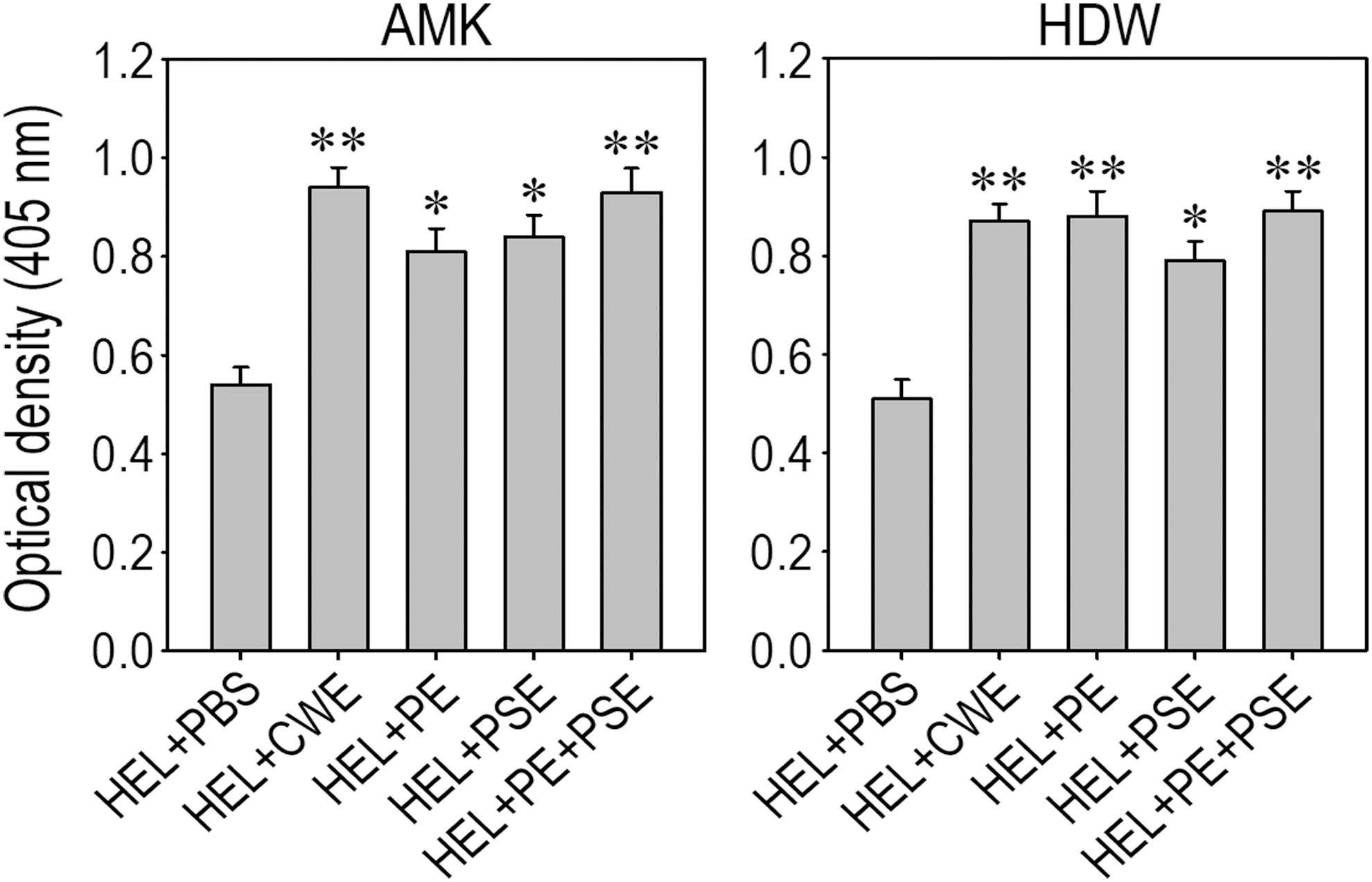
Effects of the medicine samples on anti-HEL IgG production in HEL low-responder C57BL/6 mice. Mice were orally administered one of the AMK or HDW samples and then immunized with HEL. ELISA was performed with equal volumes of sera pooled from five individual mice per group, collected 4 days after booster immunization. The results show the optical density of anti-HEL antibodies in the sera diluted 128-fold. Results were analyzed by ANOVA and *P < .05, **P < .01 compared to the values of the control group administered with PBS alone (n = 5). HEL, hen egg-white lysozyme.
To explore the possible mechanisms involved in the enhancement of antigen-specific immune response induction, we determined serum titers of HEL-specific antibodies belonging to different IgG subclasses by ELISA. Oral feeding with AMK samples increased serum titers of the four different IgG subtypes specific to HEL; while the increases in IgG1 and IgG3 levels were significant at P < .01, the levels of IgG2a and IgG2b were higher at P < .05 compared to those in the control group (Fig. 5A). Similarly, the serum titers of HEL-specific IgG subtypes were higher in most of the groups of mice fed HDW samples compared with the control mice, even though there was no significant difference in HEL-specific IgG2a level between the groups fed PBS only, crude water, or polysaccharide extract alone (Fig. 5B).
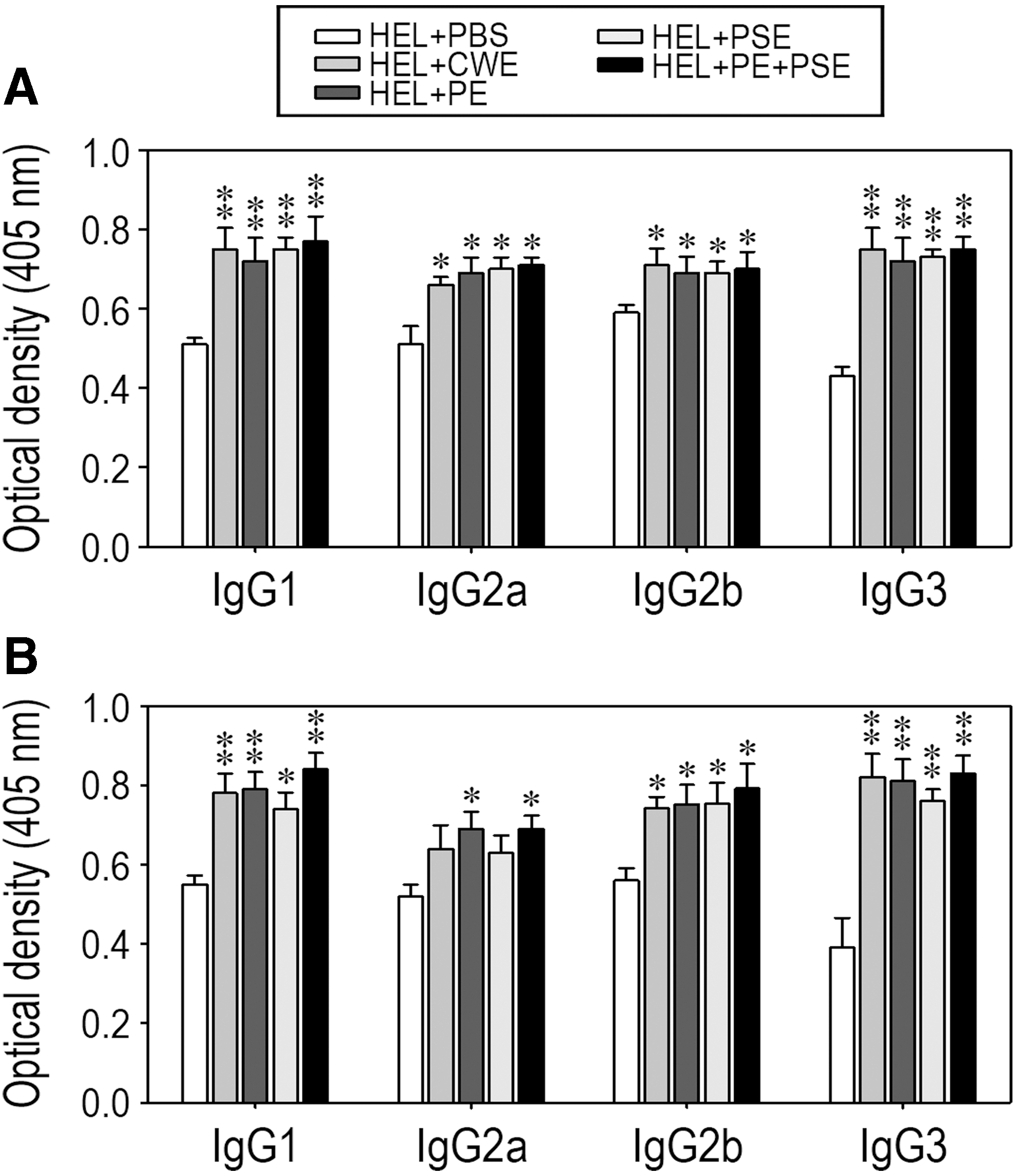
Effects of oral ingestion of medicine samples on the levels of Ig subclasses after HEL immunization. All procedures in this experiment were performed as described in Figure 4, except that isotype-specific secondary antibodies were used to identify the Ig subclasses. The results represent the values obtained with the sera diluted 128-fold, and panels
Oral feeding of AMK and HDW samples enhanced HEL-specific lymphocyte proliferative responses
To assess the effects of AMK and HDW samples on HEL-specific lymphocyte proliferative responses, splenocytes were collected from the mice 10 days after a single immunization with HEL and stimulated with various concentrations (1–10 μM) of HEL for 72 h. The results of the TdR incorporation assay showed that the proliferative capacity of splenocytes in response to the antigen was significantly higher in the groups fed the AMK samples compared to the control group (Fig. 6A). Similarly, the groups administered HDW samples showed significantly greater proliferative responses to HEL stimulation than did the control group, although stimulation with more than 1 μM HEL induced significant increases (Fig. 6B). These results suggest that oral feeding with AMK or HDW samples enhances the antigen-specific cellular immune response.
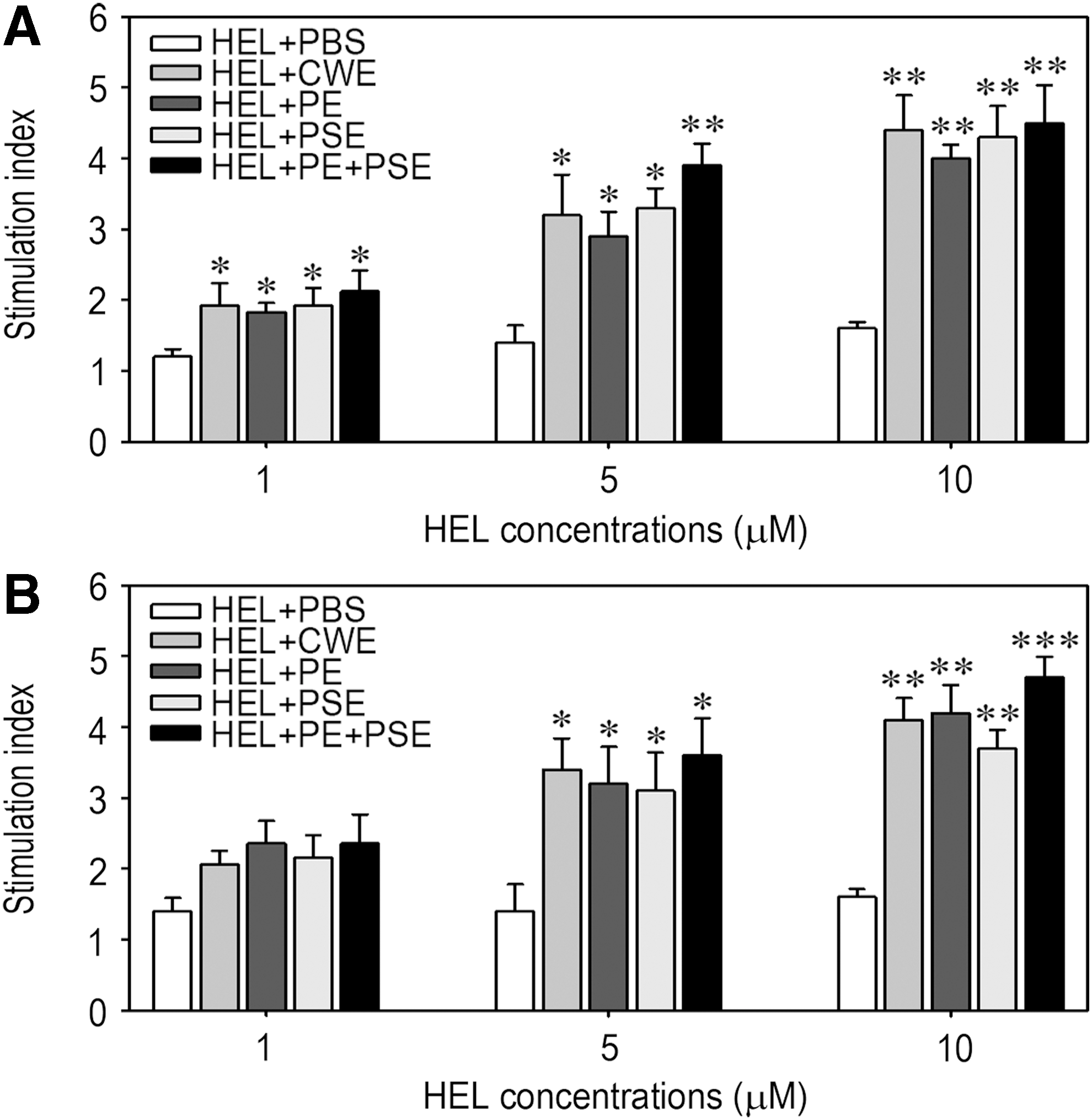
Effect of the medicine samples on anti-HEL T cell proliferative responses. Splenocytes were collected from C57BL/6 mice administered AMK
Oral administration of AMK and HDW samples increased IFN-γ production in HEL-stimulated splenocytes
We further investigated whether oral feeding with AMK or HDW samples would enhance the HEL-specific cellular immune response by determining the levels of Th1- and Th2-type T cell-specific cytokines IFN-γ and IL-4. Exogenous stimulation with more than 1 μM HEL induced a significantly higher production of IFN-γ in cultured splenocytes of the mice fed AMK samples compared to that of the control mice (Fig. 7A). Similarly, the splenocytes from the groups administered HDW samples produced significantly higher levels of IFN-γ after stimulation with HEL than did the cells from the control group (Fig. 7B). Specifically, the production of IFN-γ in splenocytes collected from the mice fed in combination with HDW protein and polysaccharide extracts was enhanced significantly, even at stimulation with 1 μM HEL. However, IL-4 production in the splenocytes collected from the mice fed AMK or HDW was not enhanced after stimulation with HEL compared to that from the control mice (data not shown). These results suggest that oral administration of AMK or HDW samples stimulates Th1-type T cells, whereas it has little effect on the selective priming of Th2-type T cells.
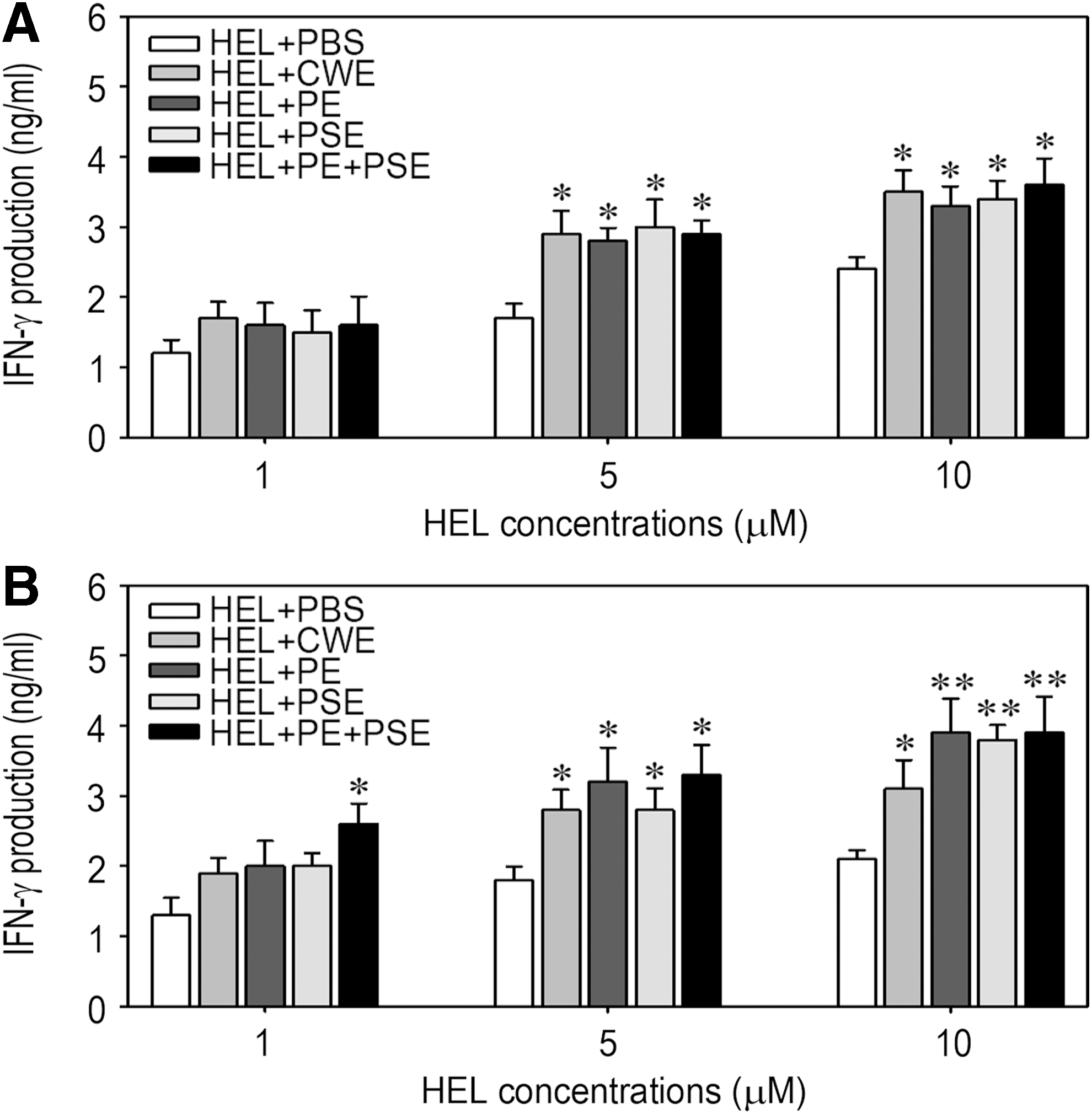
Effect of oral ingestion of the medicine samples on IFN-γ production by splenocytes. Splenocytes were prepared from the C57BL/6 mice administered AMK
Discussion
Glycoproteins and polysaccharides are capable of stimulating lymphocyte proliferation and cytokine production, as well as inducing an immune response. Traditional herbal medicines are attractive sources of immune-modulating compounds that enhance the functional activities of immune cells and inhibit the growth of tumor cells. 21 –23 We previously found that the stimulatory effects of traditional medicines on lymphocyte proliferation, antibody production, and cytokine secretion are closely associated with the presence of glycoproteins in a medicine. 13,24
In this study, we explored whether protein and/or polysaccharides were responsible for the immune stimulation activities of the traditional herbal plants AMK, HAL, SBD, and HDW. Protein and polysaccharide extracts of the plants exerted greater activities on lymphocyte proliferation than the crude water extracts at the same concentrations, thus supporting their important role as active constituents for immune stimulation. However, the results from a cytokine ELISA led us to deduce that the stimulating activity of the medicinal plant samples on cytokine production is not correlated with their effect on lymphocyte proliferation. Because the crude water extracts enhanced TNF-α production to a similar degree as did the protein or polysaccharide samples at the same concentrations, we also could not exclude the presence of immune stimulating components in crude water extracts other than macromolecules such as glycoproteins.
We then sought to verify the nature of the active constituents responsible for the splenocyte-stimulating activity induced by the medicinal plant samples. CBB or Schiff reagent staining revealed the presence of glycoproteins with various molecular weights in the protein and polysaccharide samples. Treatment with pronase E or NaIO4, but not PMB or heating, significantly reduced lymphocyte proliferative activity of the protein samples, whereas LPS- or Con A-stimulated lymphocyte proliferation was clearly inhibited by treatment with PMB or heating. However, treatment of the polysaccharide samples with PMB or heating did not diminish their activity on lymphocyte proliferation (data not shown). Taken as a whole, the current findings support that the immune-stimulating activity of the protein and polysaccharide samples of the traditional herbal plants is mainly due to glycoproteins, and these samples do not contain lectin compounds and LPS-like mitogens. 8,13,24
In general, glycoproteins may have large number of molecular weights. As shown by CBB or Schiff reagent staining, all protein and polysaccharide samples from the herbal plants revealed the presence of glycoproteins with various molecular weights. It is worth investigating which of the glycoproteins are the major active ingredients responsible for the traditional medicine-induced immune stimulation.
In a previous study, we eluted the groups of AMK protein bands into three fractions and designated them as F1, F2, and F3 to correspond to the molecular masses of around 150, 30, and <21 kDa, respectively. 24 When the stimulating activity of the fractions was tested, the F2 fraction stimulated splenocyte proliferation and cytokine production more greatly than did other fractions. This observation with the current findings suggest that the glycoproteins, especially those with a molecular mass around 27–30 kDa, are the active ingredients of AMK in modulating immune responses. It was also reported that a polysaccharide with 109.4 kDa, composed of glucose, mannose, arabinose, galactose, xylose, ribose, and rhamnose, was the active component to modulate immune activity on macrophages. 25 In this study, we showed the presence of glycoproteins mainly around 27 and 150 kDa in AMK polysaccharide extract by Schiff reagent staining. Considering these findings, we postulate that AMK polysaccharides with 100–150 kDa are the major potential ingredients for modulating intestinal immune responses.
SBD, known as Ban-Zhi-Lian, contains a large number of polysaccharides, flavones, and alkaloids. 26 A recent study reported that a water-soluble polysaccharide containing 53.6% carbohydrates, 38.5% uronic acid, and 8.2% proteins is an active ingredient of SBD and its molecular weight is 26 kDa composed of arabinose, mannose, glucose, and galactose at the ratio of 1.31:1.00:3.59:1.59. 27 In contrast, very little information on the characterization of immune active ingredients from the glycoproteins and polysaccharides isolated from HAL and HDW is available. It is also important to note that these macromolecules are differently isolated from traditional medicines according to extraction methods applied. Further experiments to characterize the active ingredients from glycoproteins and/or polysaccharides tested in this study will be needed.
In an initial attempt to explore the effects of the medicinal plant samples on humoral immune response induction, we compared the activity of the samples to induce anti-HEL-specific antibody production. As evidenced by the results that oral feeding with AMK or HDW significantly enhanced total IgG level in the sera of HEL-immunized mice, these samples induced an antigen-specific humoral immune response. This observation was corroborated by the observation that oral feeding of AMK and HDW stimulated HEL-specific lymphocyte proliferation in the mice. However, we found that the level of HEL-specific IgG2a, which is closely related to the stimulation of Th1 type T cell populations, was not increased dramatically compared to that of IgG1 and IgG3 subtypes, which are associated with the stimulation of B lymphocytes. These results led us to postulate that the traditional herbal plants are capable of stimulating antigen-specific immune responses through a direct activation of B lymphocytes, as well as Th1-type T lymphocytes, rather than Th2-type T lymphocytes.
CD4+ T cells are divided into Th1- and Th2-type T cell populations according to their pattern of cytokine production in response to antigenic stimulation. 28 The balanced interaction between Th1 and Th2 cell subpopulations is critical for immune response induction and modulation. 29,30 Th1 cells produce IFN-γ, IL-2, and lymphotoxin and mediate cell-mediated immune reactions, while Th2 cells are capable of producing IL-4, IL-5, IL-6, IL-10, and IL-13, which are mostly associated with antibody and allergic immune responses. 31 –33
The results from cytokine ELISA revealed that the level of IFN-γ was significantly enhanced after exogenous HEL stimulation in cultured splenocytes that were collected from mice administered AMK or HDW samples compared to that of the control mice. However, none of the samples increased IL-4 level in the cultured splenocytes after HEL stimulation (data not shown). These results agree with our previous findings that the HEL-specific serum titers of IgG1 and IgG2a were increased in mice fed a traditional medicine prescription compared to those of control mice, whereas the production of IFN-γ, but not IL-4, was enhanced in splenocytes from the mice fed the medicine after HEL stimulation. 11,12
There was also a similar report showing that oral feeding with an aqueous extract of Epimedium koreanum Nakai increased the levels of total serum IgG with IgG1 and IgG2a, as well as IFN-γ, compared with that of the untreated control. 34 These findings indicate that oral administration with AMK or HDW samples may stimulate lymphocytes through two separate pathways. One is that oral treatment with the samples increases the level of IFN-γ versus IL-4; thus, the elevated HEL-specific stimulation index is more associated with enhanced proliferation of Th1-type T cells rather than Th2-type T cells. Another possible pathway is that oral feeding of AMK or HDW increases titers of antigen-specific antibodies related to the direct stimulation of B lymphocytes.
Indeed, HEL-specific sera titers of IgG2b and IgG3, which directly stimulate B lymphocytes, were significantly increased in the mice fed the samples compared to that of the control mice. However, it is important to note that all samples of the medicinal plants enhanced greatly the production of TNF-α and IL-6 in cultured splenocytes. Although in vitro results do not directly correlate with the in vivo situation, these results may suggest that the sample-mediated increases in the HEL-specific antibody response were at least, in part, modulated by Th2-type lymphocytes in addition to the direct stimulation of B lymphocytes. Collectively, we consider that direct lymphocyte stimulation by oral ingestion of the medicinal plants plays an important role in enhancing anti-HEL-specific humoral immune responses, in which the selective priming of Th1- and Th2-type specific T cell populations is partially associated. 35,36
One of the key findings in this study is that glycoproteins and/or polysaccharides are the active macromolecules responsible for the immune stimulation of traditional medicines. In contrast, a major concern with oral administration of immune stimulating glycoproteins will be the degradation of protein components in the stomach and gut during the digestive process. 37
It is suggested that the degradation of glycoproteins reduces their activity to elicit an immune response. The current findings that treatment of protein samples with pronase E or NaIO4 inhibited their stimulating effects on splenocyte proliferation support such suggestion. It is likely that the use of delivery vehicles such as liposomes may protect glycoproteins from degradation and ferry their intact forms to the gut. However, more specific experiments regarding the protein degradation of glycoproteins and its protection will be required. Furthermore, additional investigations to characterize the physicochemical properties of glycoproteins of the medicinal plants and to clarify the signaling pathways that mediate immune stimulation, as well as to clarify how the plant samples affect the production of other co-stimulatory molecules, are necessary.
In summary, immunological dysfunctions can occur when the activities of T and B lymphocytes are suppressed and/or when the balance between Th1- and Th2 type T cell populations is disrupted. 38,39 Much interest has been paid to the use of traditional herbal medicines as therapeutic approaches for immune modulation. Our current findings highlight that traditional medicines are capable of stimulating antigen-specific immune responses in vivo and in vitro. Our results also indicate that the stimulation of B lymphocytes and Th1-type T cells, rather than Th2-type T cells, is more positively induced by oral administration of traditional medicines.
Footnotes
Acknowledgment
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ077822012016)” Rural Development Administration, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.